Acetylcholinesterase and Nicotinic Acetylcholine Receptors in Schistosomes and Other Parasitic Helminths


Abstract
:1. Introduction
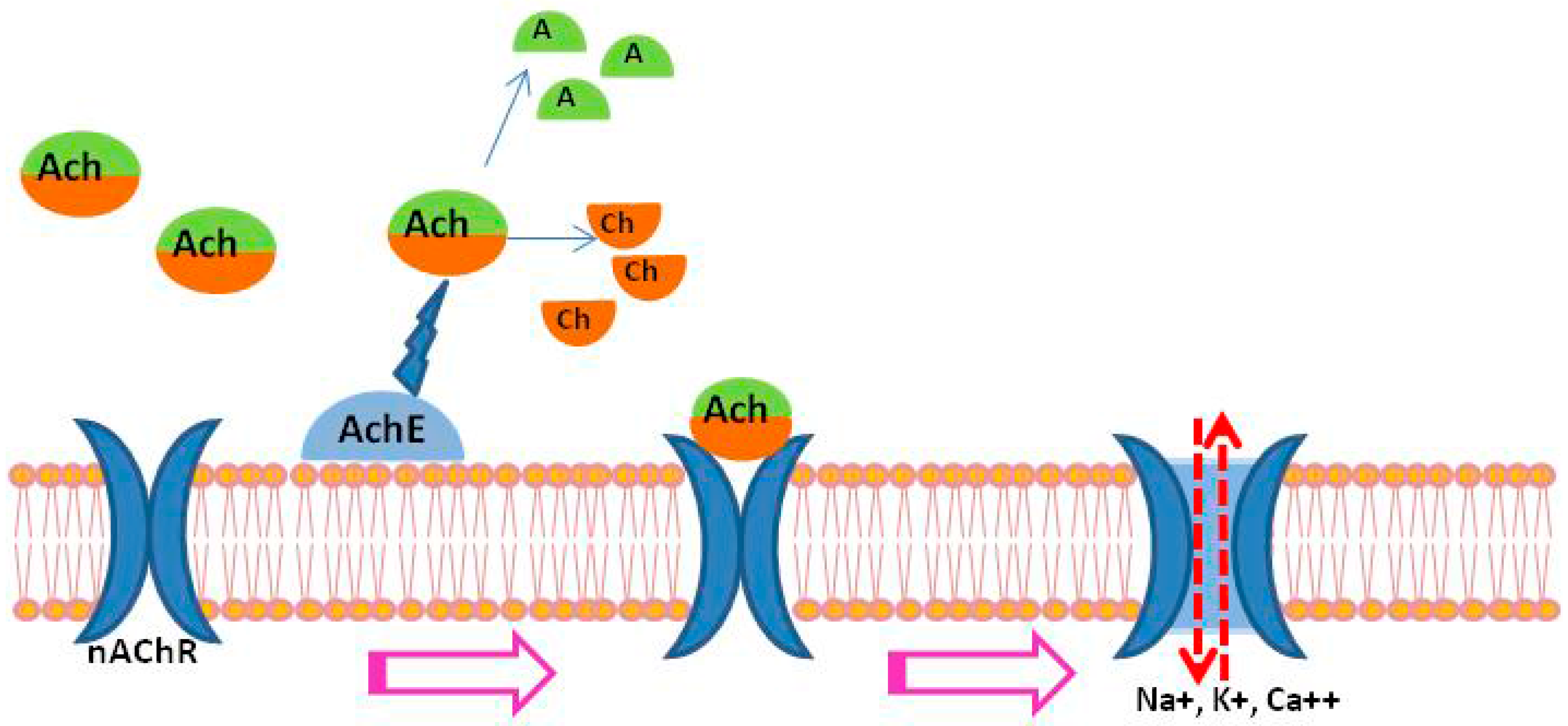
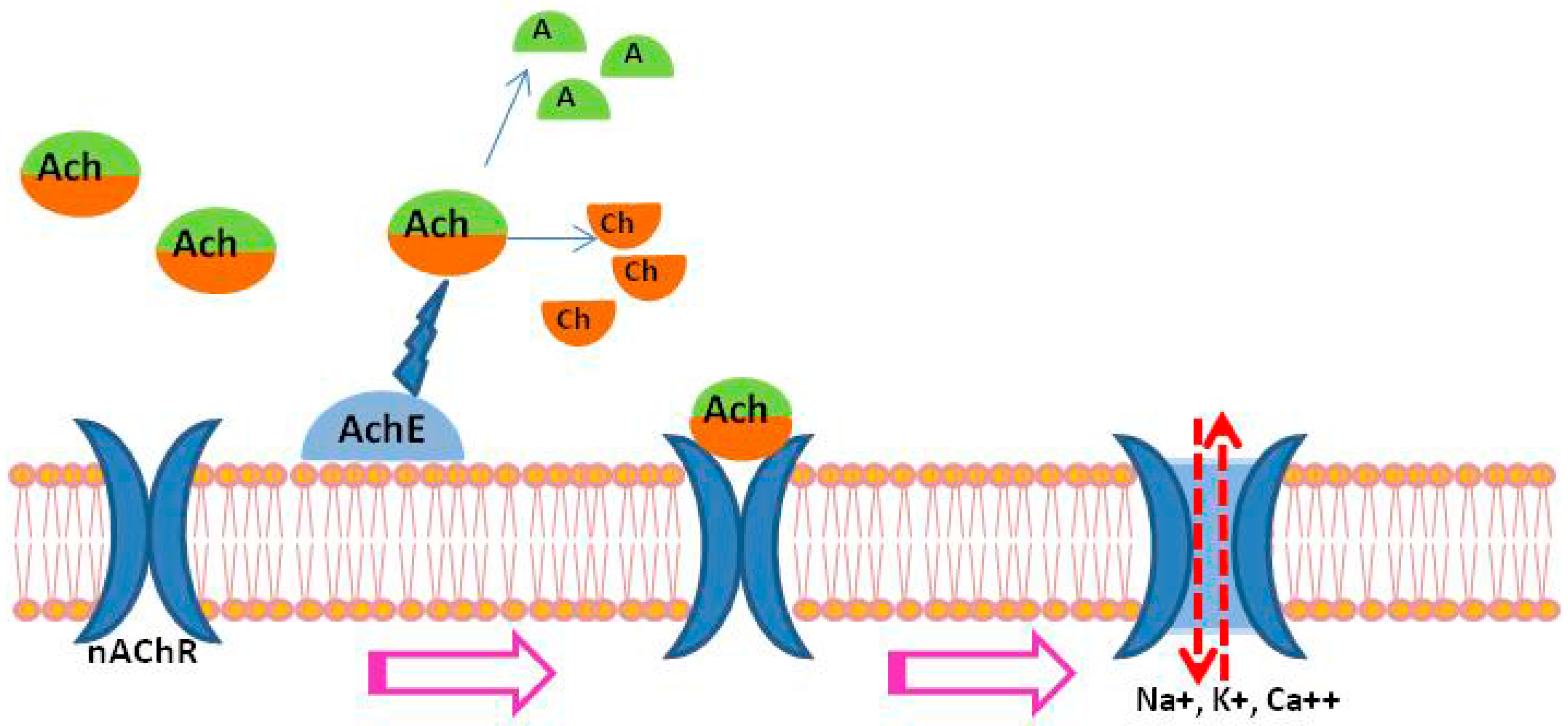
2. Acetylcholinesterase (AChE) in Schistosomes
3. Nicotinic Acetylcholine Receptors (nAChRs) in Schistosomes
4. AChE and AChRs in Other Helminths
5. Conclusions
Supplementary Materials
Acknowledgments
Conflicts of Interest
Abbreviations
| PZQ | praziquantel |
| AChE | acetylcholinesterase |
| ACh | acetylcholine |
| nAChRs | nicotinic acetylcholine receptors |
| GPI | glycophosphatidylinositol |
| PIPL-C | PI-specific phospholipase C |
| SmACCs | S. manosni acetylcholine gated chloride channels |
| ShAR1α | S. haematobium nicotinic acetylcholine receptor 1 α |
| ShAR1β | S. haematobium nicotinic acetylcholine receptor 1 β |
| ShAR2β | S. haematobium nicotinic acetylcholine receptor 2 β |
References
- Beaumier, C.M.; Gillespie, P.M.; Hotez, P.J.; Bottazzi, M.E. New vaccines for neglected parasitic diseases and dengue. Transl. Res. 2013, 162, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.P.; Martin, R.J. Ion-channels on parasite muscle: Pharmacology and physiology. Invert. Neurosci. 2007, 7, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, R.; Gauvry, N.; Weber, S.S.; Skripsky, T.; Bouvier, J.; Wenger, A.; Schroeder, F.; Desaules, Y.; Hotz, R.; Goebel, T.; et al. Identification of the amino-acetonitrile derivative monepantel (AAD 1566) as a new anthelmintic drug development candidate. Parasitol. Res. 2008, 103, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Bueding, E.; Liu, C.L.; Rogers, S.H. Inhibition by metrifonate and dichlorvos of cholinesterases in schistosomes. Br. J. Pharmacol. 1972, 46, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, J.E.; Wilson, R.A. Schistosoma mansoni: A scanning electron microscope study of the developing schistosomulum. Parasitology 1980, 81, 553–564. [Google Scholar] [CrossRef] [PubMed]
- El Ridi, R.A.; Tallima, H.A. Novel therapeutic and prevention approaches for schistosomiasis: Review. J. Adv. Res. 2013, 4, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, R. Strategies for Control of Infection and Disease: Current Practice and Future Potential. In Schistosomiasis; Mahmoud, A.A.F., Ed.; Imperial College Press: London, UK, 2001. [Google Scholar]
- Salafsky, B.; Fusco, A.C.; Whitley, K.; Nowicki, D.; Ellenberger, B. Schistosoma mansoni: Analysis of cercarial transformation methods. Exp. Parasitol. 1988, 67, 116–127. [Google Scholar] [CrossRef]
- Jones, A.K.; Bentley, G.N.; Parra, W.G.O.; Agnew, A. Molecular characterization of an acetylcholinesterase implicated in the regulation of glucose scavenging by the parasite Schistosoma. FASEB J. 2002, 16, 441–443. [Google Scholar] [PubMed]
- Thompson, A.J.; Lester, H.A.; Lummis, S.C. The structural basis of function in Cys-loop receptors. Q. Rev. Biophys. 2010, 43, 449–499. [Google Scholar] [CrossRef] [PubMed]
- Quinn, D.M. Acetylcholinesterase: Enzyme structure, reaction dynamics and virtual transition states. Chem. Rev. 1987, 87, 955–979. [Google Scholar] [CrossRef]
- Wolstenholme, A.J. Ion channels and receptor as targets for the control of parasitic nematodes. Int. J. Parasitol. Drugs Drug Resist. 2011, 1, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Bennekou, P. The voltage-gated non-selective cation channel from human red cells is sensitive to acetylcholine. Biochim. Biophys. Acta 1993, 1147, 165–167. [Google Scholar] [CrossRef]
- Barker, L.R.; Bueding, E.; Timms, A.R. The possible role of acetylcholine in Schistosoma mansoni. Br. J. Pharmacol. 1966, 26, 656–665. [Google Scholar] [CrossRef]
- Day, T.A.; Chen, G.Z.; Miller, C.; Tian, M.; Bennett, J.L.; Pax, R.A. Cholinergic inhibition of muscle fibres isolated from Schistosoma mansoni (Trematoda:Digenea). Parasitology 1996, 113, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Zouridakis, M.; Zisimopoulou, P.; Poulas, K.; Tzartos, S.J. Recent advances in understanding the structure of nicotinic acetylcholine receptors. IUBMB Life 2009, 61, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.; Flood, P.; McArdle, J.J.; Brenner, H.R. Advances in neurobiology of the neuromuscular junction: Implications for the anesthesiologist. Anesthesiology 2002, 96, 202–231. [Google Scholar] [CrossRef] [PubMed]
- Kabbani, N.; Nordman, J.C.; Corgiat, B.A.; Veltri, D.P.; Shehu, A.; Seymour, V.A.; Adams, D.J. Are nicotinic acetylcholine receptors coupled to G proteins? Bioessays 2013, 35, 1025–1034. [Google Scholar] [CrossRef] [PubMed]
- Bentley, G.N.; Jones, A.K.; Oliveros Parra, W.G.; Agnew, A. ShAR1alpha and ShAR1beta: Novel putative nicotinic acetylcholine receptor subunits from the platyhelminth blood fluke Schistosoma. Gene 2004, 329, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Bueding, E. Acetylcholinesterase activity of Schistosoma mansoni. Br. J. Pharmacol. 1952, 7, 563–566. [Google Scholar] [CrossRef]
- Pax, R.A.; Day, T.A.; Miller, C.L.; Bennett, J.L. Neuromuscular physiology and pharmacology of parasitic flatworms. Parasitology 1996, 113 (Suppl. 1), S83–S96. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, B.; Tarrab-Hazdai, R.; Himmeloch, S.; Arnon, R. Acetylcholinesterase from Schistosoma mansoni: Immunological characterization. Immunol. Lett. 1991, 28, 167–174. [Google Scholar] [CrossRef]
- Bentley, G.N.; Jones, A.K.; Agnew, A. Expression and comparative functional characterisation of recombinant acetylcholinesterase from three species of Schistosoma. Mol. Biochem. Parasitol. 2005, 141, 119–123. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Gobert, G.N.; Du, X.; Pali, G.; Cai, P.; Jones, M.K.; McManus, D.P. Functional characterisation of Schistosoma japonicum acetylcholinesterase. Parasit. Vectors 2016, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Levi-Schaffer, F.; Tarrab-Hazdai, R.; Schryer, M.D.; Arnon, R.; Smolarsky, M. Isolation and partial characterization of the tegumental outer membrane of schistosomula of Schistosoma mansoni. Mol. Biochem. Parasitol. 1984, 13, 283–300. [Google Scholar] [CrossRef]
- Pearson, M.S.; Becker, L.; Driguez, P.; Young, N.D.; Gaze, S.; Mendes, T.; Li, X.H.; Doolan, D.L.; Midzi, N.; Mduluza, T.; et al. Of monkeys and men: Immunomic profiling of sera from humans and non-human primates resistant to schistosomiasis reveals novel potential vaccine candidates. Front. Immunol. 2015, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Arnon, R.; Silman, I.; Tarrab-Hazdai, R. Acetylcholinesterase of Schistosoma mansoni—Functional correlates. Contributed in honor of Professor Hans Neurath’s 90th birthday. Protein Sci. 1999, 8, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, B.; Parizade, M.; Ortega, E.; Tarrab-Hazdai, R.; Zilberg, D.; Arnon, R. Monoclonal antibodies against acetylcholinesterase of Schistosoma mansoni: Production and characterization. Hybridoma 1995, 14, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Agnew, A. Schistosoma: Rate of glucose import is altered by acetylcholine interaction with tegumental acetylcholine receptors and acetylcholinesterase. Exp. Parasitol. 1995, 81, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro-dos-Santos, G.; Verjovski-Almeida, S.; Leite, L.C. Schistosomiasis—A century searching for chemotherapeutic drugs. Parasitol. Res. 2006, 99, 505–521. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Tarrab-Hazdai, R.; Espinoza, B.; Arnon, R.; Agnew, A. The amount of acetylcholinesterase on the parasite surface reflects the differential sensitivity of schistosome species to metrifonate. Parasitology 1994, 108, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Harder, A. Chemotherapeutic approaches to schistosomes: Current knowledge and outlook. Parasitol. Res. 2002, 88, 395–397. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.L. Why do some nematode parasites of the alimentary tract secrete acetylcholinesterase? Int. J. Parasitol. 1996, 26, 499–508. [Google Scholar] [CrossRef]
- Berriman, M.; Haas, B.J.; LoVerde, P.T.; Wilson, R.A.; Dillon, G.P.; Cerqueira, G.C.; Mashiyama, S.T.; Al-Lazikani, B.; Andrade, L.F.; Ashton, P.D.; et al. The genome of the blood fluke Schistosoma mansoni. Nature 2009, 460, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Young, N.D.; Jex, A.R.; Li, B.; Liu, S.; Yang, L.; Xiong, Z.; Li, Y.; Cantacessi, C.; Hall, R.S.; Xu, X.; et al. Whole-genome sequence of Schistosoma haematobium. Nat. Genet. 2012, 44, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Schistosoma japonicum Genome Sequencing and Functional Analysis Consortium. The Schistosoma japonicum genome reveals features of host-parasite interplay. Nature 2009, 460, 345–351. [Google Scholar]
- Massoulie, J.; Bon, S. The molecular forms of cholinesterase and acetylcholinesterase in vertebrates. Annu. Rev. Neurosci. 1982, 5, 57–106. [Google Scholar] [CrossRef] [PubMed]
- Tarrab-Hazdai, R.; Toker, L.; Silman, I.; Arnon, R. Acetylcholinesterase from Schistosoma mansoni: Interaction of globular species with heparin. Biochem. J. 1999, 344, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Tarrab-Hazdai, R.; Levi-Schaffer, F.; Gonzales, G.; Arnon, R. Acetylcholinesterase of Schistosoma mansoni. Molecular forms of the solubilized enzyme. Biochim. Biophys. Acta 1984, 790, 61–69. [Google Scholar] [CrossRef]
- Espinoza, B.; Silman, I.; Arnon, R.; Tarrab-Hazdai, R. Phosphatidylinositol-specific phospholipase C induces biosynthesis of acetylcholinesterase via diacylglycerol in Schistosoma mansoni. Eur. J. Biochem. 1991, 195, 863–870. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, B.; Tarrab-Hazdai, R.; Silman, I.; Arnon, R. Acetylcholinesterase in Schistosoma mansoni is anchored to the membrane via covalently attached phosphatidylinositol. Mol. Biochem. Parasitol. 1988, 29, 171–179. [Google Scholar] [CrossRef]
- MacDonald, K.; Buxton, S.; Kimber, M.J.; Day, T.A.; Robertson, A.P.; Ribeiro, P. Functional characterization of a novel family of acetylcholine-gated chloride channels in Schistosoma mansoni. PLoS Pathog. 2014, 10, e1004181. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Alsford, S.; Jones, A.; Agnew, A. Nicotinic acetylcholine receptors on the surface of the blood fluke Schistosoma. Mol. Biochem. Parasitol. 1995, 71, 127–134. [Google Scholar] [CrossRef]
- Bentley, G.N.; Jones, A.K.; Agnew, A. ShAR2beta, a divergent nicotinic acetylcholine receptor subunit from the blood fluke Schistosoma. Parasitology 2007, 134, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Agnew, A. Glucose uptake rates by Schistosoma mansoni, S. haematobium, and S. bovis adults using a flow in vitro culture system. J. Parasitol. 1995, 81, 637–640. [Google Scholar] [CrossRef] [PubMed]
- Dani, J.A. Neuronal Nicotinic Acetylcholine receptor structure and function and response to nicotine. Int. Rev. Neurobiol. 2015, 124, 3–19. [Google Scholar] [PubMed]
- Beech, R.N.; Callanan, M.K.; Rao, V.T.; Dawe, G.B.; Forrester, S.G. Characterization of cys-loop receptor genes involved in inhibitory amine neurotransmission in parasitic and free living nematodes. Parasitol. Int. 2013, 62, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Mansour, T.E. Chemotherapeutic Targets in Parasites Contemporary Strategies; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Sukhdeo, S.C.; Sangster, N.C.; Mettrick, D.F. Effects of cholinergic drugs on longitudinal muscle contractions of Fasciola hepatica. J. Parasitol. 1986, 72, 858–864. [Google Scholar] [CrossRef] [PubMed]
- Rapson, E.B.; Chilwan, A.S.; Jenkins, D.C. Acetylcholinesterase secretion—A parameter for the interpretation of in vitro anthelmintic screens. Parasitology 1986, 92, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, B.M.; Rothwell, T.L.; Bremner, K.C.; Schnitzerling, H.J.; Nolan, J.; Keith, R.K. Acetylcholinesterase secretion by parasitic nematodes—I. Evidence for secretion of the enzyme by a number of species. Int. J. Parasitol. 1973, 3, 589–597. [Google Scholar] [CrossRef]
- Vaux, R.; Schnoeller, C.; Berkachy, R.; Roberts, L.B.; Hagen, J.; Gounaris, K.; Selkirk, M.E. Modulation of the Immune Response by Nematode Secreted Acetylcholinesterase Revealed by Heterologous Expression in Trypanosoma musculi. PLoS Pathog. 2016, 12, e1005998. [Google Scholar] [CrossRef] [PubMed]
- Sukhdeo, S.C.; Sukhdeo, M.V.; Mettrick, D.F. Histochemical localization of acetylcholinesterase in the cerebral ganglia of Fasciola hepatica, a parasitic flatworm. J. Parasitol. 1988, 74, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Kemmerling, U.; Cabrera, G.; Campos, E.O.; Inestrosa, N.C.; Galanti, N. Localization, specific activity, and molecular forms of acetylcholinesterase in developmental stages of the cestode Mesocestoides corti. J. Cell Physiol. 2006, 206, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.J. Modes of action of anthelmintic drugs. Vet. J. 1997, 154, 11–34. [Google Scholar] [CrossRef]
- Jones, A.K.; Sattelle, D.B. Functional genomics of the nicotinic acetylcholine receptor gene family of the nematode, Caenorhabditis elegans. Bioessays 2004, 26, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Richmond, J.E.; Jorgensen, E.M. One GABA and two acetylcholine receptors function at the C. elegans neuromuscular junction. Nat. Neurosci. 1999, 2, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Touroutine, D.; Fox, R.M.; Von Stetina, S.E.; Burdina, A.; Miller, D.M., 3rd; Richmond, J.E. acr-16 encodes an essential subunit of the levamisole-resistant nicotinic receptor at the Caenorhabditis elegans neuromuscular junction. J. Biol. Chem. 2005, 280, 27013–27021. [Google Scholar] [CrossRef] [PubMed]
- Boulin, T.; Gielen, M.; Richmond, J.E.; Williams, D.C.; Paoletti, P.; Bessereau, J.L. Eight genes are required for functional reconstitution of the Caenorhabditis elegans levamisole-sensitive acetylcholine receptor. Proc. Natl. Acad. Sci. USA 2008, 105, 18590–18595. [Google Scholar] [CrossRef] [PubMed]
- Buxton, S.K.; Charvet, C.L.; Neveu, C.; Cabaret, J.; Cortet, J.; Peineau, N.; Abongwa, M.; Courtot, E.; Robertson, A.P.; Martin, R.J. Investigation of acetylcholine receptor diversity in a nematode parasite leads to characterization of tribendimidine- and derquantel-sensitive nAChRs. PLoS Pathog. 2014, 10, e1003870. [Google Scholar] [CrossRef] [PubMed]
- Duguet, T.B.; Charvet, C.L.; Forrester, S.G.; Wever, C.M.; Dent, J.A.; Neveu, C.; Beech, R.N. Recent Duplication and Functional Divergence in Parasitic Nematode Levamisole-Sensitive Acetylcholine Receptors. PLoS Negl. Trop. Dis. 2016, 10, e0004826. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.M.; Robertson, A.P.; Brown, L.; Williams, T.; Woods, D.J.; Martin, R.J.; Sattelle, D.B.; Wolstenholme, A.J. The nicotinic acetylcholine receptors of the parasitic nematode Ascaris suum: Formation of two distinct drug targets by varying the relative expression levels of two subunits. PLoS Pathog. 2009, 5, e1000517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.J.; Clark, C.L.; Trailovic, S.M.; Robertson, A.P. Oxantel is an N-type (methyridine and nicotine) agonist not an L-type (levamisole and pyrantel) agonist: Classification of cholinergic anthelmintics in Ascaris. Int. J. Parasitol. 2004, 34, 1083–1090. [Google Scholar] [CrossRef] [PubMed]
- Boulin, T.; Fauvin, A.; Charvet, C.L.; Cortet, J.; Cabaret, J.; Bessereau, J.L.; Neveu, C. Functional reconstitution of Haemonchus contortus acetylcholine receptors in Xenopus oocytes provides mechanistic insights into levamisole resistance. Br. J. Pharmacol. 2011, 164, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Fauvin, A.; Charvet, C.; Issouf, M.; Cortet, J.; Cabaret, J.; Neveu, C. cDNA-AFLP analysis in levamisole-resistant Haemonchus contortus reveals alternative splicing in a nicotinic acetylcholine receptor subunit. Mol. Biochem. Parasitol. 2010, 170, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Sloan, M.A.; Reaves, B.J.; Maclean, M.J.; Storey, B.E.; Wolstenholme, A.J. Expression of nicotinic acetylcholine receptor subunits from parasitic nematodes in Caenorhabditis elegans. Mol. Biochem. Parasitol. 2015, 204, 44–50. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Schistosome Species | AChE | nAChR | ||||
|---|---|---|---|---|---|---|
| Protein ID | Description | Protein ID | Conserved Domains/Motifs | |||
| Ligand Domain | Trans-Membrane Region | Cys-Loop | ||||
| S. haematobium | A_03825 | AChR 1alpha | AAR84357 * | √ | √ | √ |
| KGB33661 * (XP_012793429 *) | AChR 2beta | AAR84358 * | √ | √ | √ | |
| AChR 2beta | AAX59989 * | √ | √ | √ | ||
| KGB33101 * (XP_012792873 *; A_04487 ~) | nAChR beta 3 (Dbeta 3) subunit | A_01504 ~ | ||||
| A_01761 ~ | √ | |||||
| KGB37011 * (XP_012796773 *; A_02007 ~) | Putative nAChR alpha 9b subunit | B_00805 ~ | √ | √ | ||
| AChR beta, putative | A_05298 ~ | √ | ||||
| AAO49838 * (AAO62355 *, AAQ14322 *) | AChR-related | A_06497 ~ | √ | √ | ||
| ACh-gated chloride channel-1 (ShACC-1) | A_02378 ~ | √ | √ | √ | ||
| ACh-gated chloride channel-2 (ShACC-2) | A_06346 ~ | |||||
| S. mansoni | AAQ14321 * (CCD58664 * XP_018645024 *) | AChR alpha subunit precursor | AAR84361 * | √ | ||
| AChR non-alpha subunit precursor | AAR84362 * | √ | ||||
| ACh-gated chloride channel-1 (SmACC-1) | Smp_176310 # | √ | √ | √ | ||
| ACh-gated chloride channel-2 (SmACC-2) | Smp_142690 # | √ | √ | √ | ||
| Putative nAChR alpha 9b subunit | Smp_135040 # | |||||
| AChR-related | Smp_012000 # | √ | √ | √ | ||
| Putative nAChR subunit | Smp_101990 # | |||||
| Smp_037960 # | √ | √ | √ | |||
| Smp_157790 # | √ | √ | √ | |||
| Smp_132070 # | √ | |||||
| Smp_180570 # | √ | √ | √ | |||
| Smp_197600 # | √ | √ | ||||
| Smp_142700 # | √ | √ | ||||
| Smp_031680 # | √ | √ | √ | |||
| Smp_139330 # | √ | √ | √ | |||
| S. japonicum | Sjp_0045440 # | Putative nAChR alpha 9b subunit | Sjp_0071780 # | |||
| Sjp_0034800 # | √ | √ | √ | |||
| Neuronal AChR subunit alpha-7 | Sjp_0082390 # | √ | √ | |||
| Sjp_0070510 # | AChR-related | Sjp_0131150 # | √ | √ | √ | |
| Sjp_0036280 # | Putative nAChR subunit | Sjp_0015560 # | √ | √ | ||
| Sjp_0066940 # | √ | |||||
| ANH56887 [24] * | ACh-gated chloride channel (SjACC-1) | Sjp_0115170 # | ||||
| Schistosome Species | nAChR | Cys-Loop Sequence |
|---|---|---|
| S. haematobium | AAR84357 | CNIDILWFPFDEQSC |
| AAR84358 | CDIEVNWFPFDSQNC | |
| AAX59989 | CQVEITYFPFDSQVC | |
| A_01761 | CSVDIKYFPFDRQKC | |
| A_06497 | CPLDVSFFPFDYQTC | |
| A_02378 | CPVKIKYFPYDKQVC | |
| S. mansoni | AAR84362 | CDIEVNWFPFDSQNC |
| Smp_139330 | CDIEVNWFPFDSQNC | |
| AAR84361 | CNIDILWFPFDEQSC | |
| Smp_031680 | CNIDILWFPFDEQSC | |
| Smp_197600 | CKIDITYFPFDDQSC | |
| Smp_157790 | CKIDIKSFPFDEQTC | |
| Smp_132070.1 | CPIDIKNFPFDYQHC | |
| Smp_132070.2 | CPIDIKNFPFDYQHC | |
| Smp_012000 | CPLDVSFFPFDYQTC | |
| Smp_142690 | CQVEITYFPFDSQVC | |
| Smp_037960 | CEVEITYFPFDTQIC | |
| Smp_176310 | CPVKIKYFPYDKQVC | |
| Smp_180570 | CQVDITLFPFDQQNC | |
| S. japonicum | Sjp_0034800 | CNVDVLYFPFDHQLC |
| Sjp_0131150 | CPLDVSFFPFDYQTC | |
| Sjp_0082390 | CDIEVNWFPFDSQNC |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, H.; Liu, C.; Du, X.; McManus, D.P. Acetylcholinesterase and Nicotinic Acetylcholine Receptors in Schistosomes and Other Parasitic Helminths. Molecules 2017, 22, 1550. https://doi.org/10.3390/molecules22091550
You H, Liu C, Du X, McManus DP. Acetylcholinesterase and Nicotinic Acetylcholine Receptors in Schistosomes and Other Parasitic Helminths. Molecules. 2017; 22(9):1550. https://doi.org/10.3390/molecules22091550
Chicago/Turabian StyleYou, Hong, Chang Liu, Xiaofeng Du, and Donald P. McManus. 2017. "Acetylcholinesterase and Nicotinic Acetylcholine Receptors in Schistosomes and Other Parasitic Helminths" Molecules 22, no. 9: 1550. https://doi.org/10.3390/molecules22091550
APA StyleYou, H., Liu, C., Du, X., & McManus, D. P. (2017). Acetylcholinesterase and Nicotinic Acetylcholine Receptors in Schistosomes and Other Parasitic Helminths. Molecules, 22(9), 1550. https://doi.org/10.3390/molecules22091550