Exogenous Melatonin Alleviates Alkaline Stress in Malus hupehensis Rehd. by Regulating the Biosynthesis of Polyamines


Abstract
:1. Introduction
2. Results
2.1. The Effect of Melatonin on Phenotypes of Malus hupehensis Rehd. under Alkaline Stress
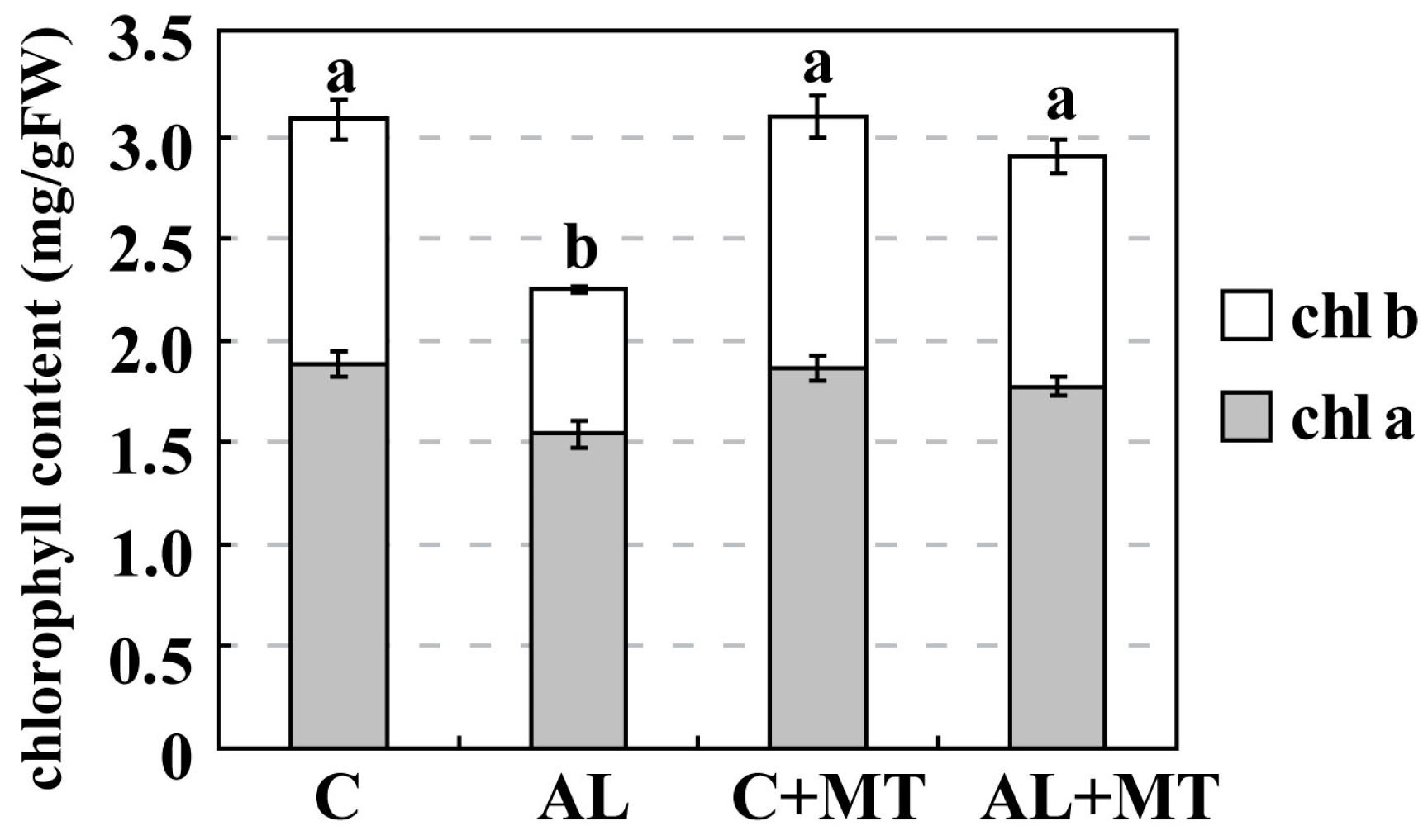
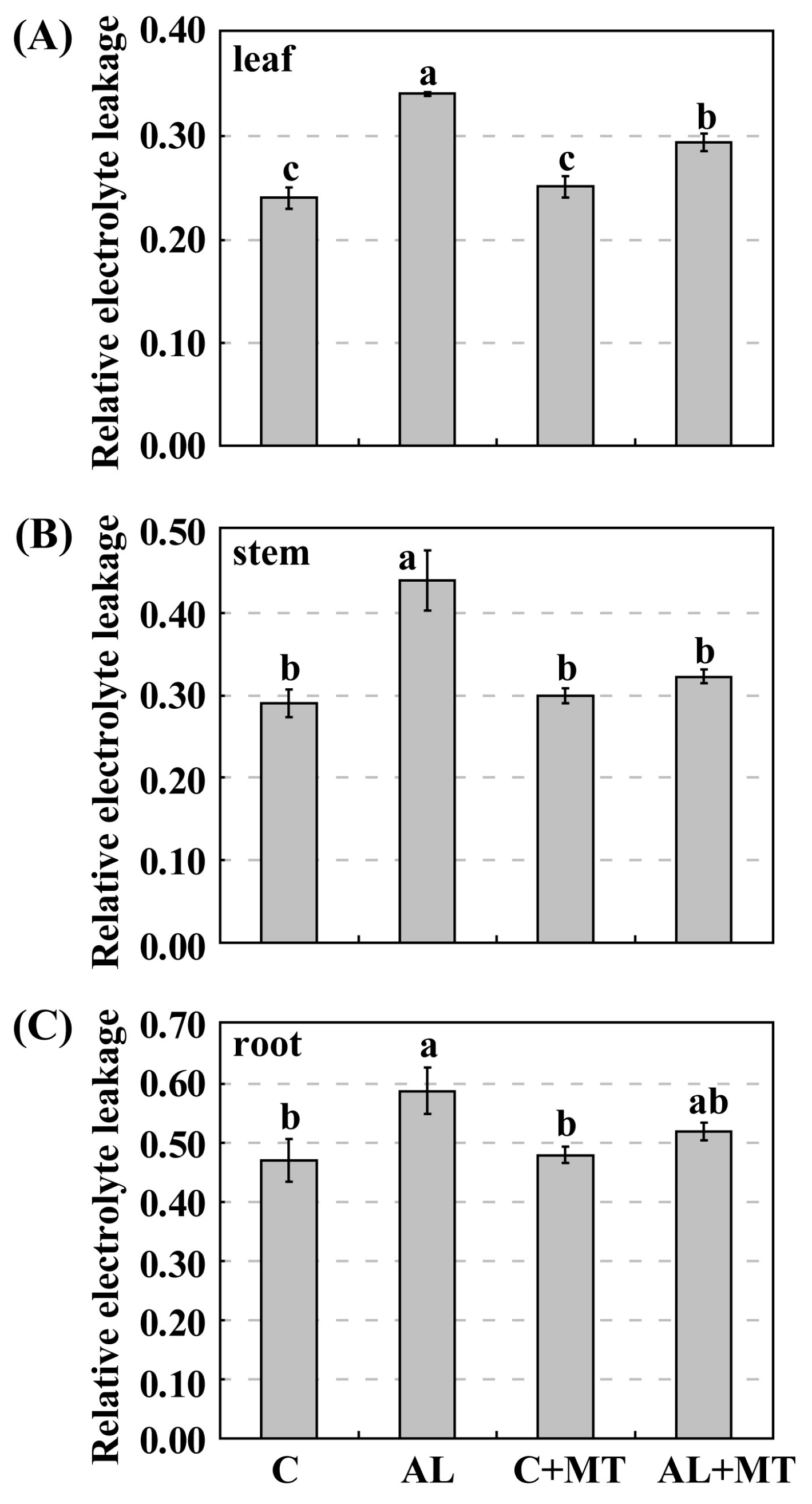
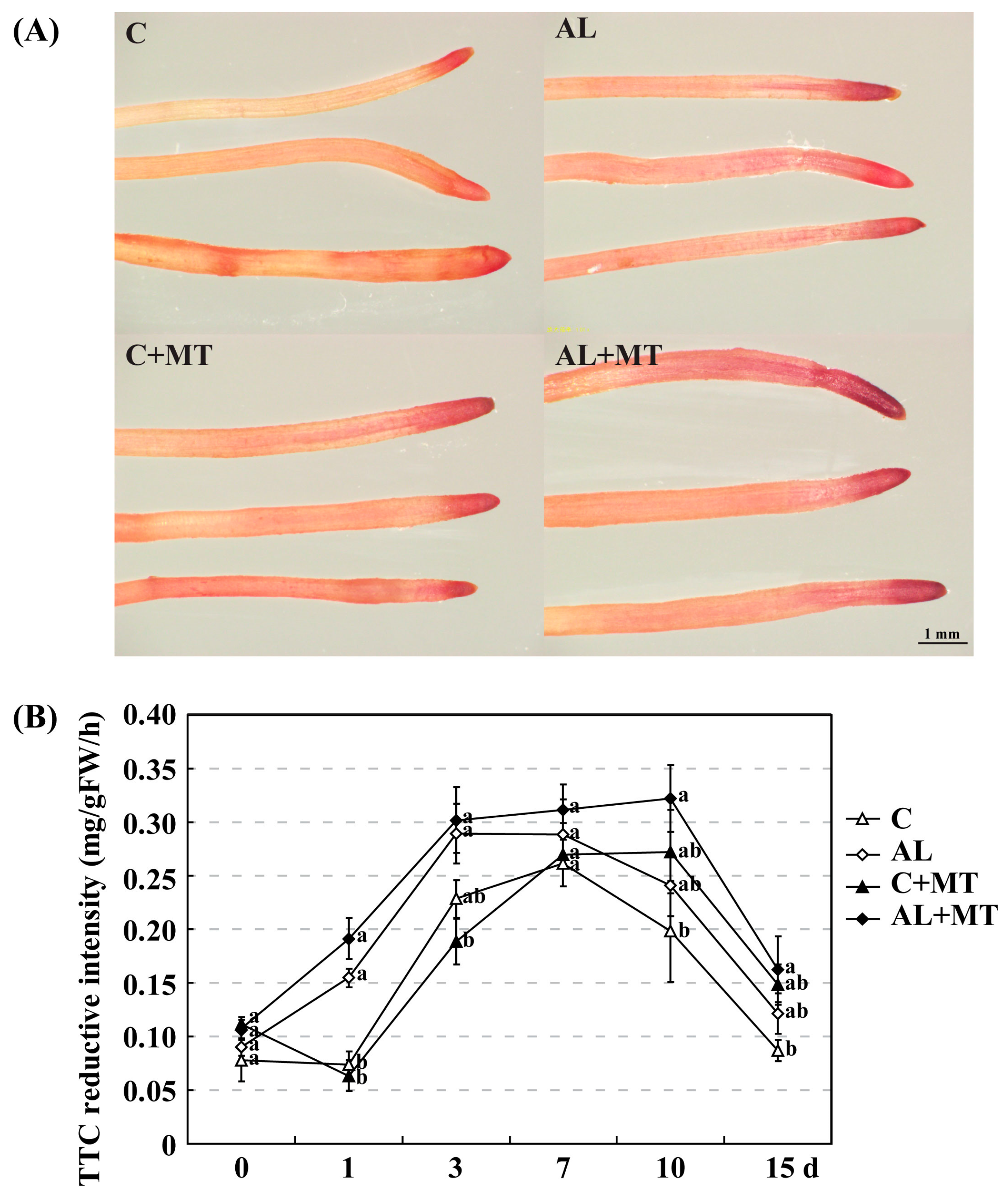
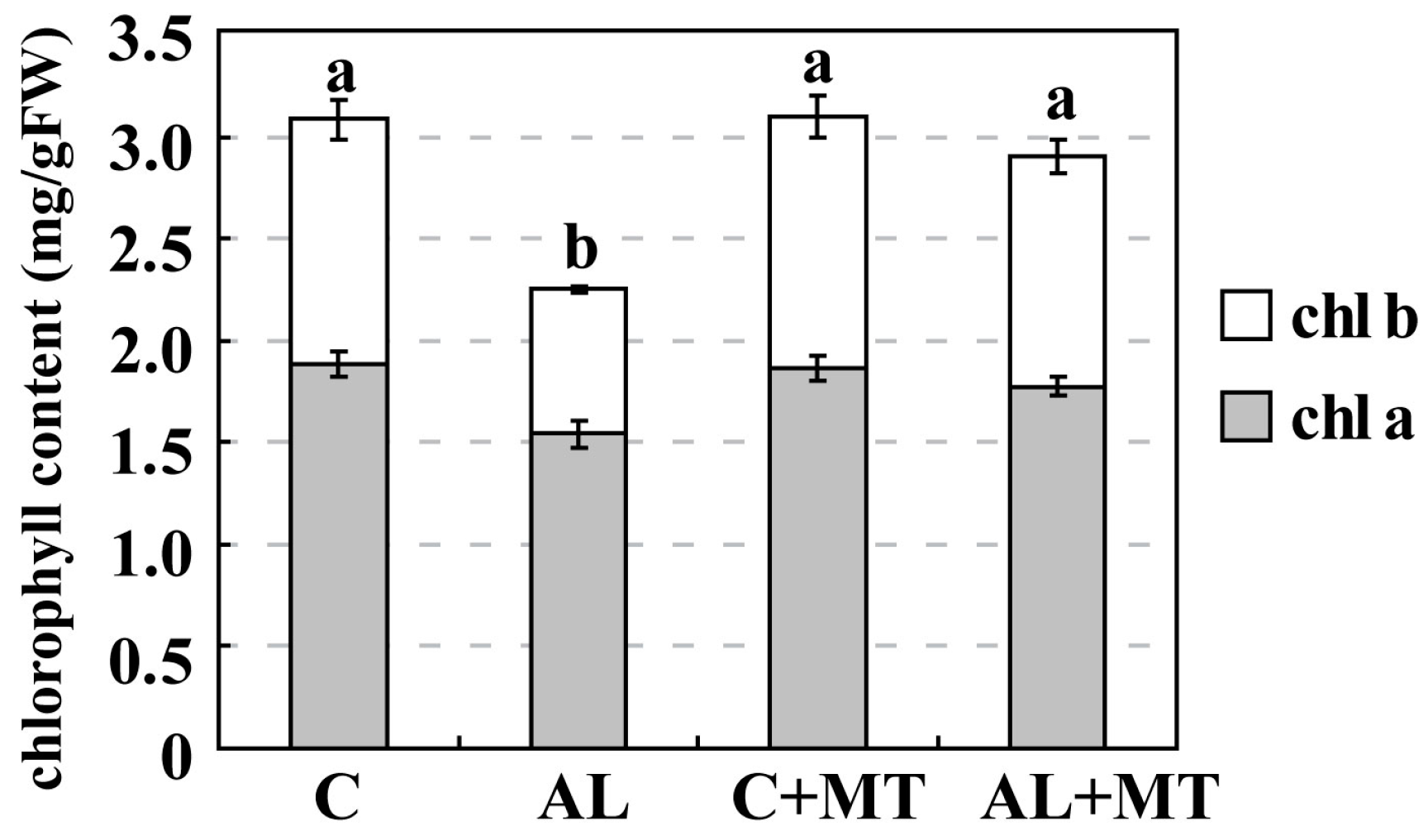
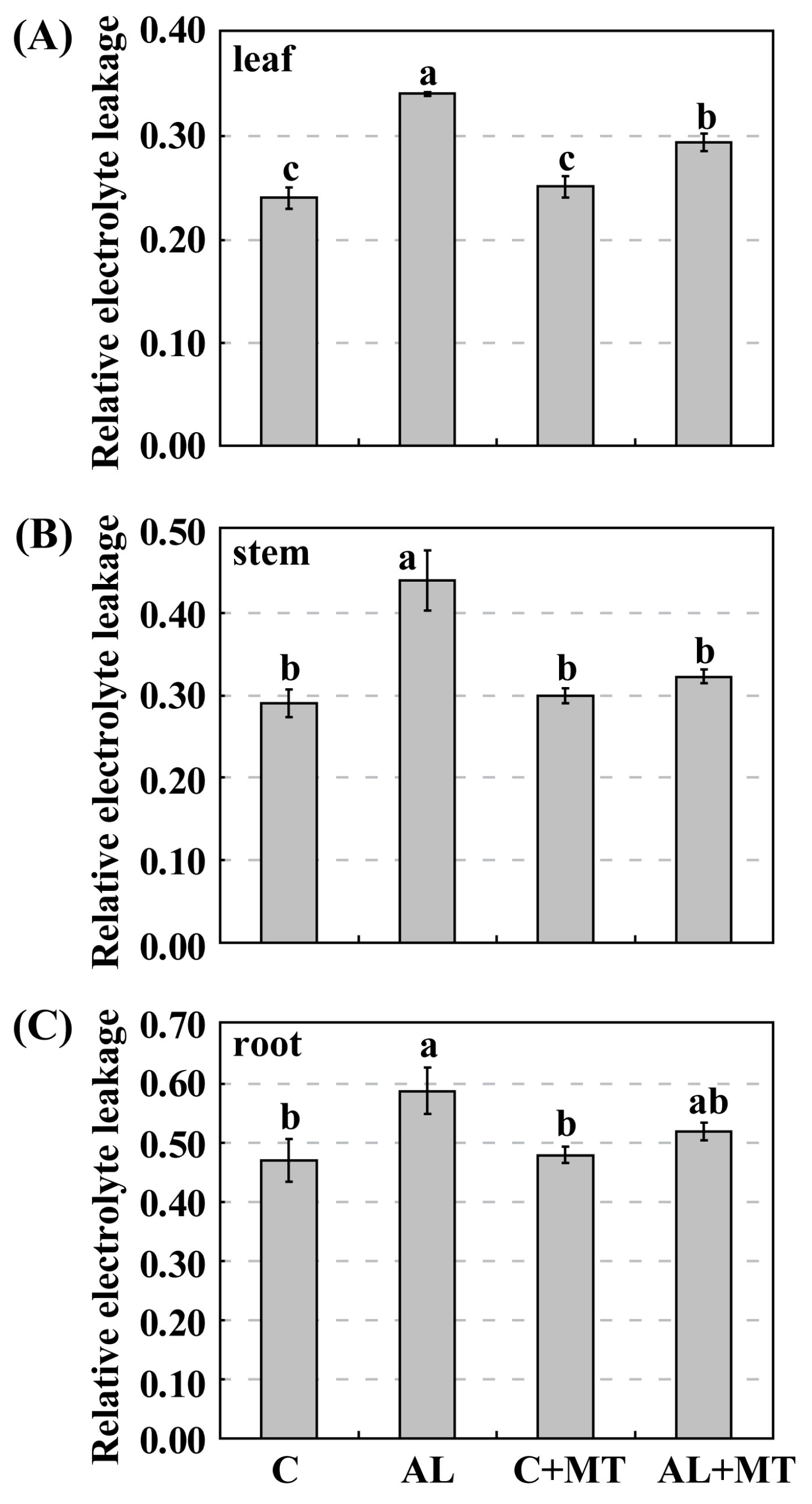
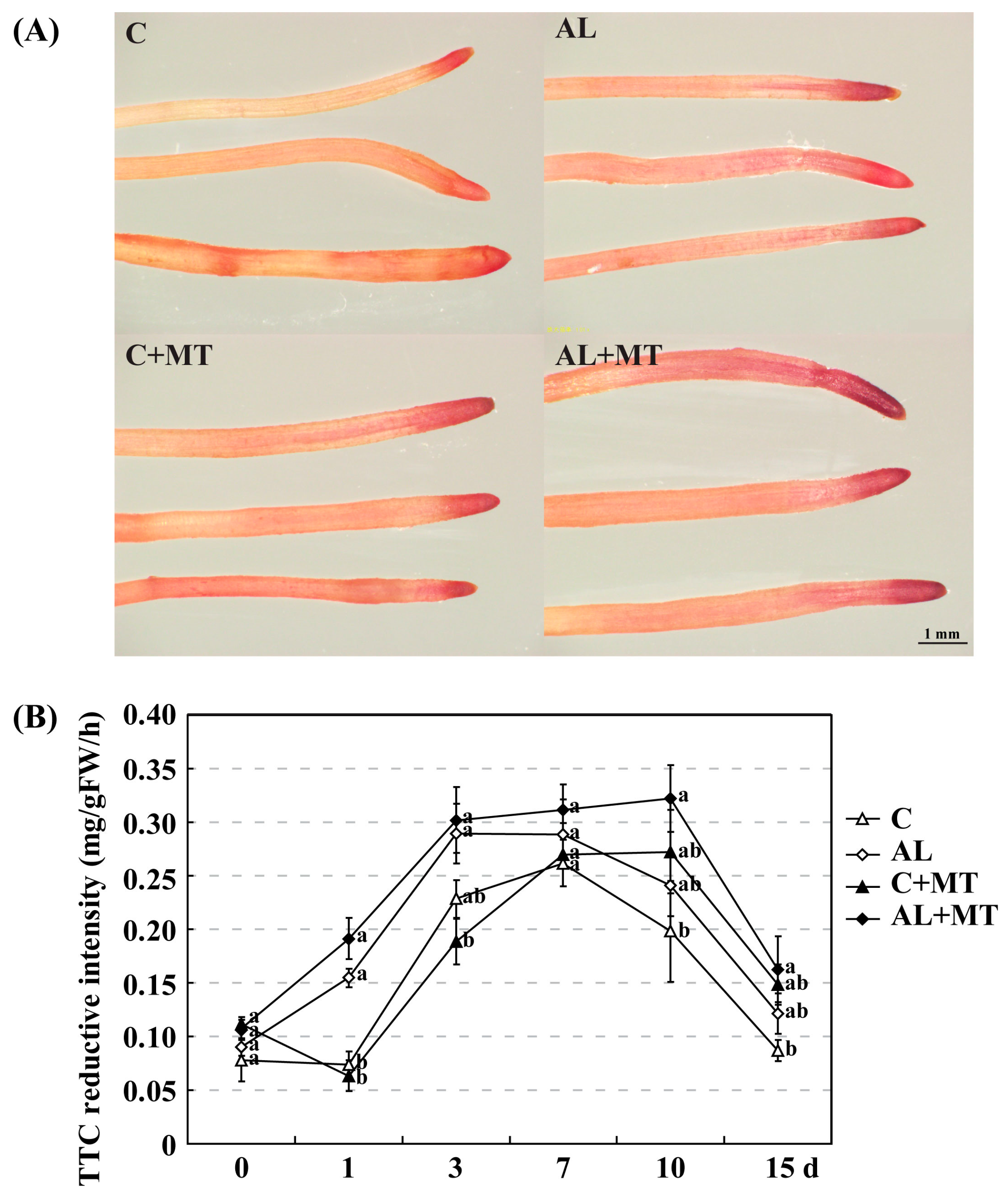
2.2. The Effect of Melatonin on Physiological State of Malus hupehensis Rehd. under Alkaline Stress
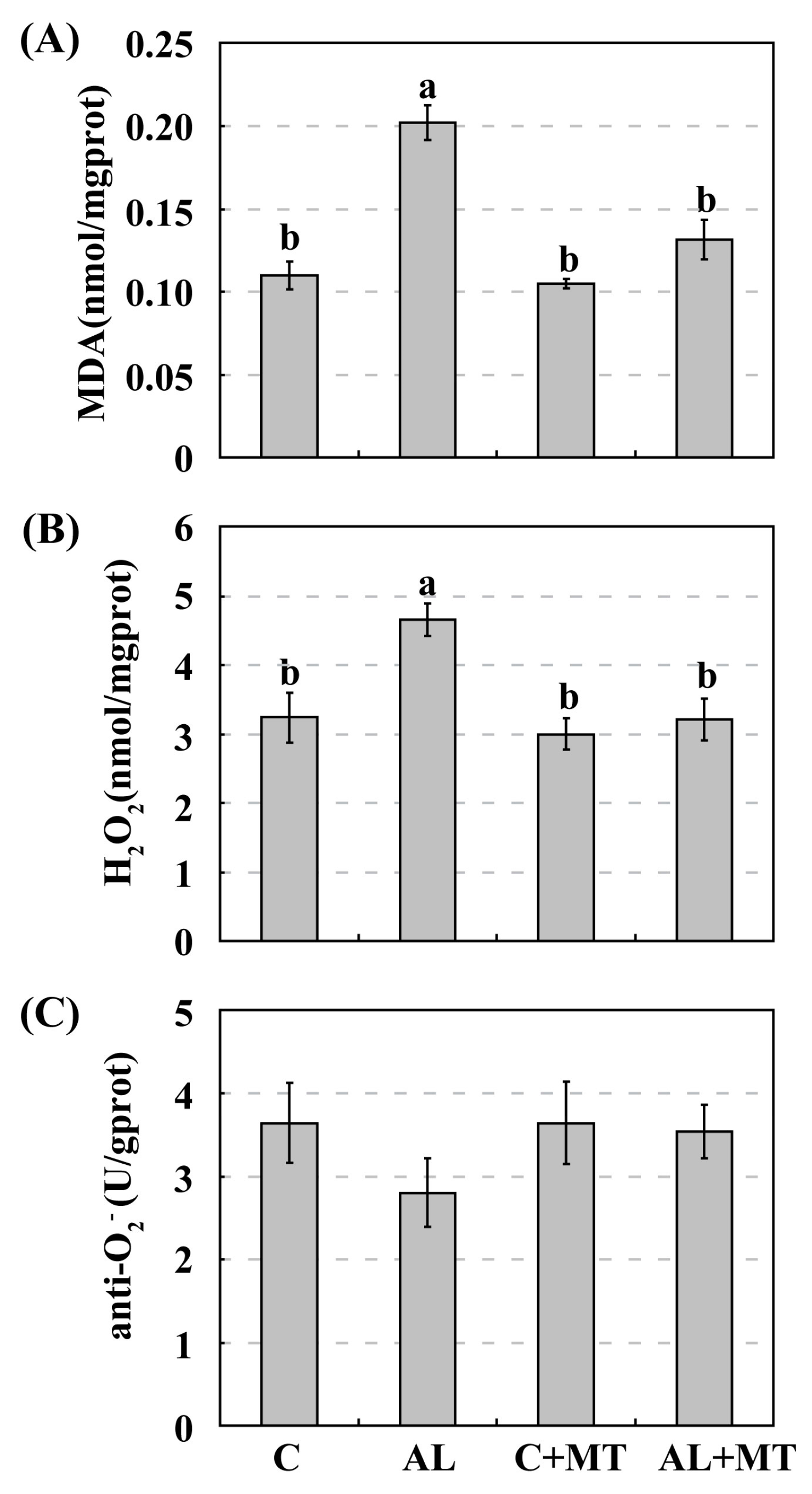
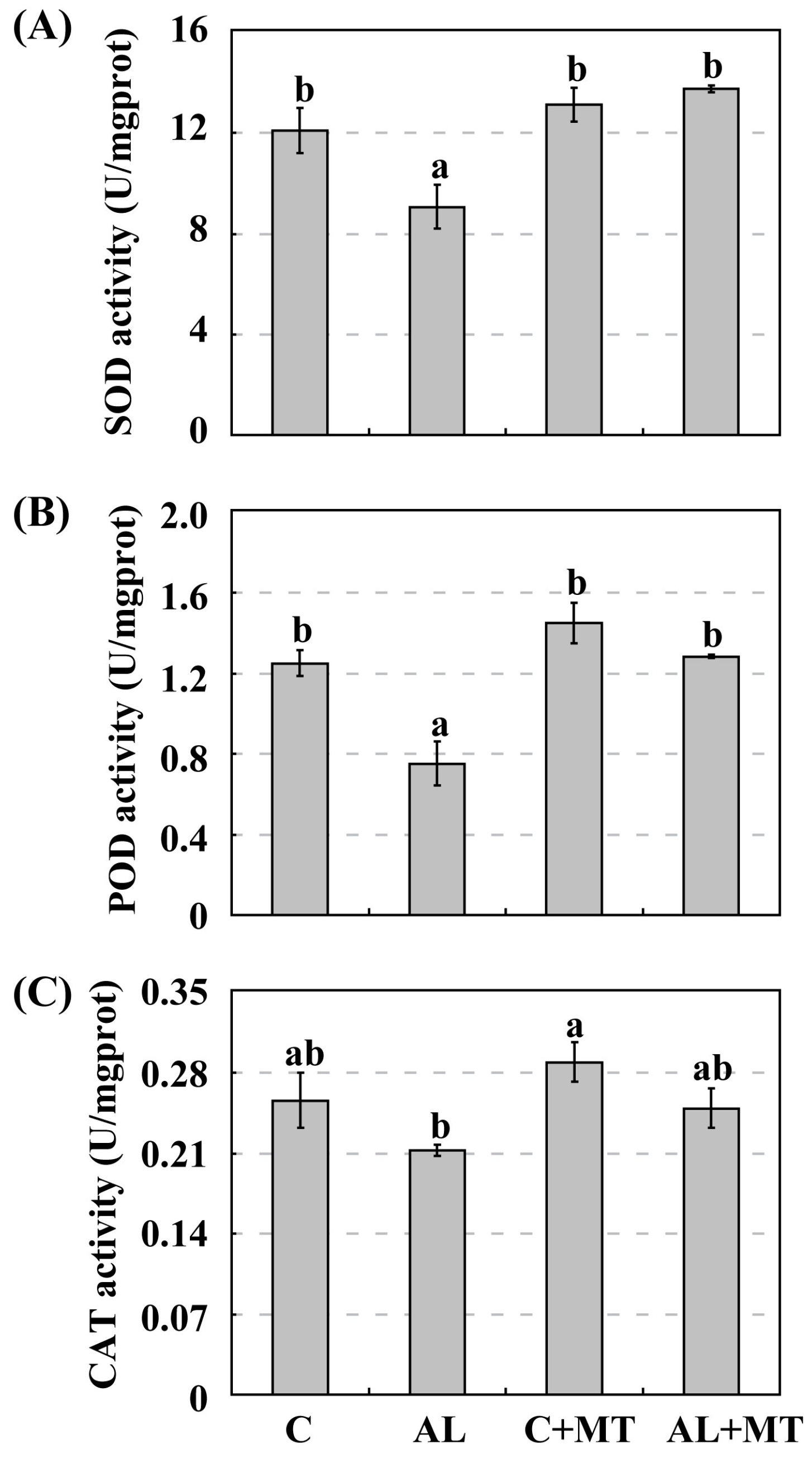
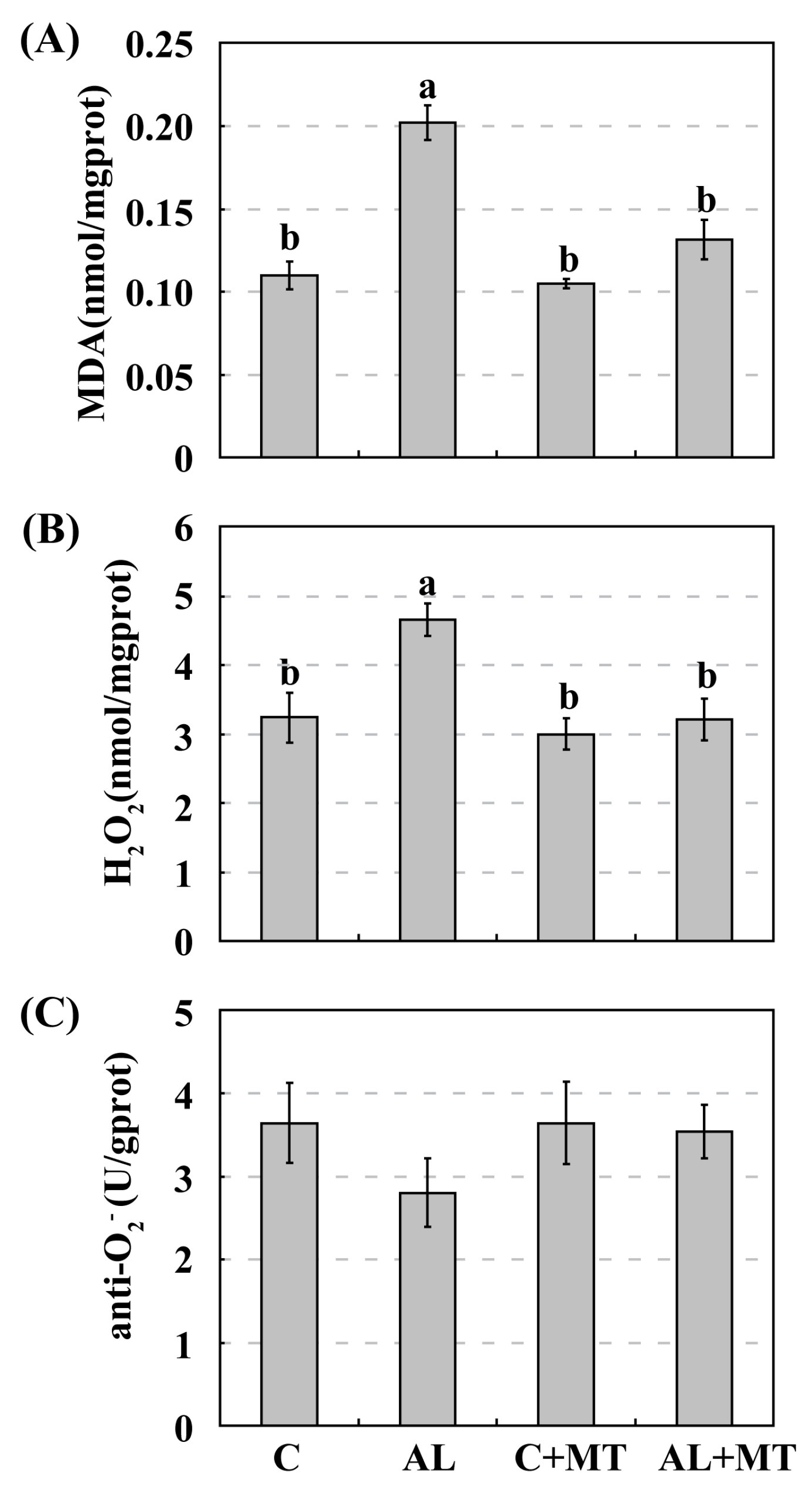
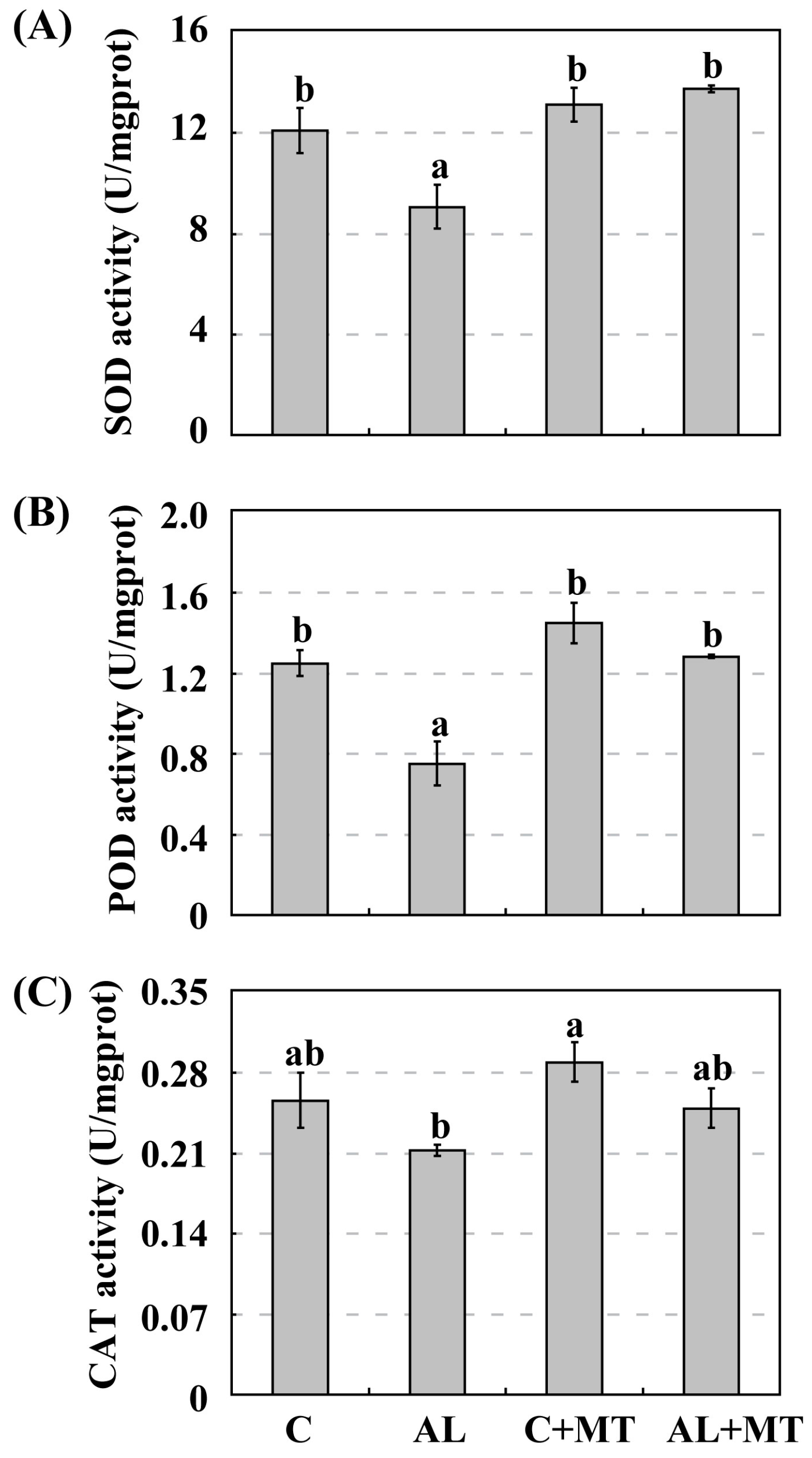
2.3. The Effect of Melatonin on Antioxidant System in Malus hupehensis Rehd. under Alkaline Stress
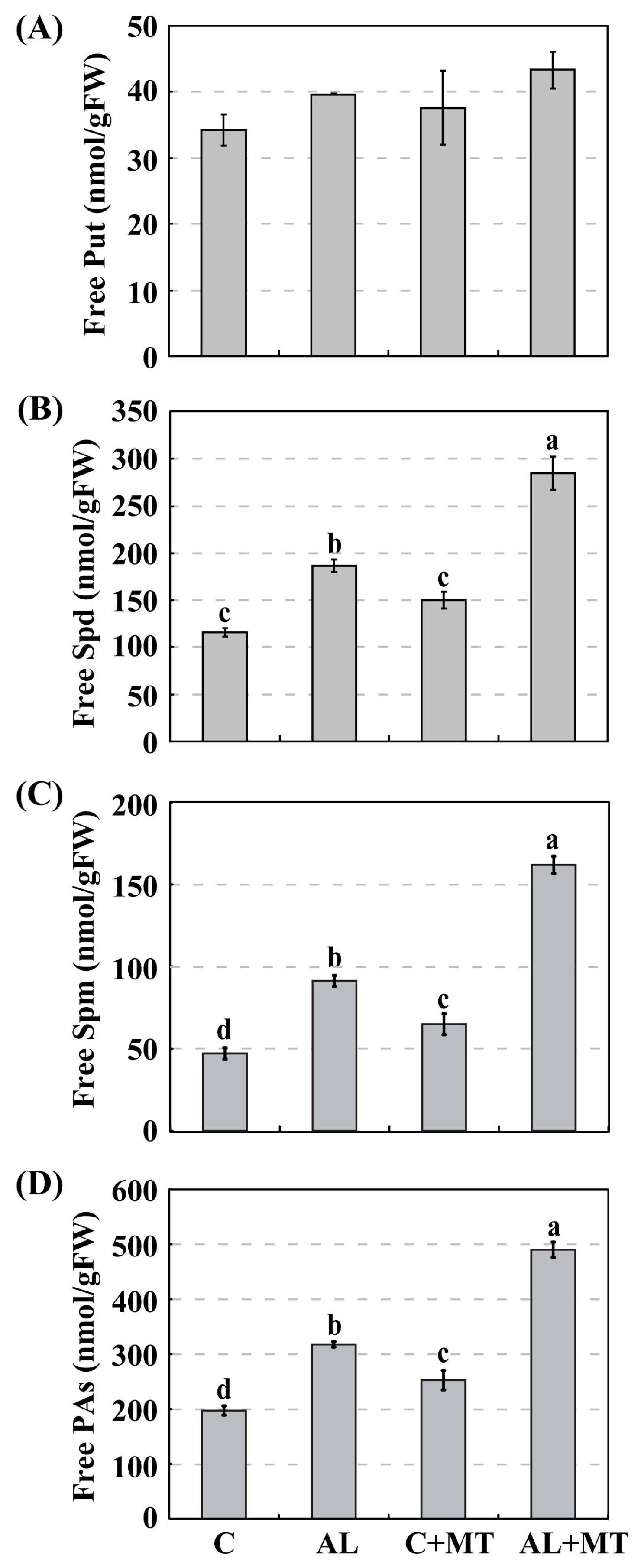
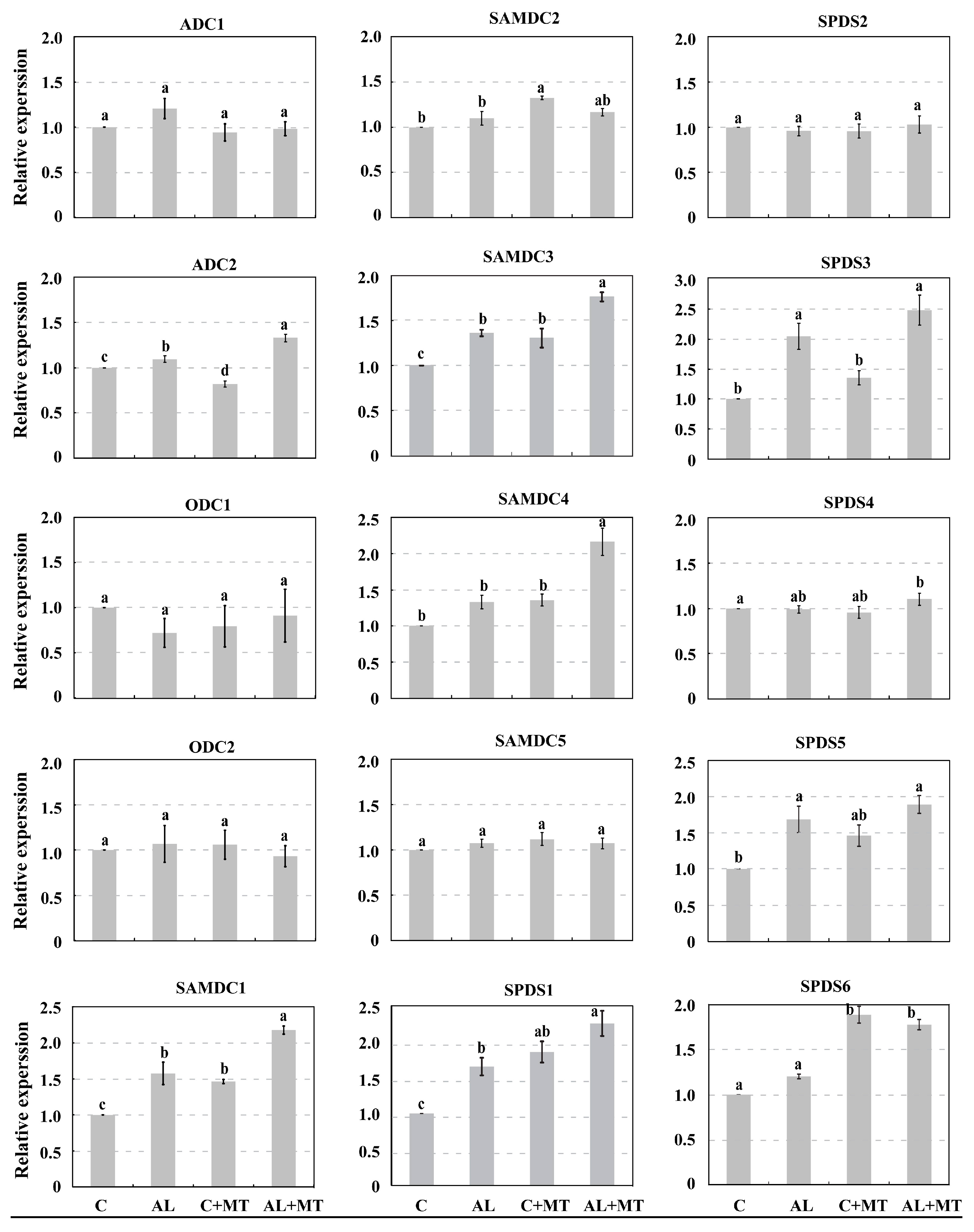
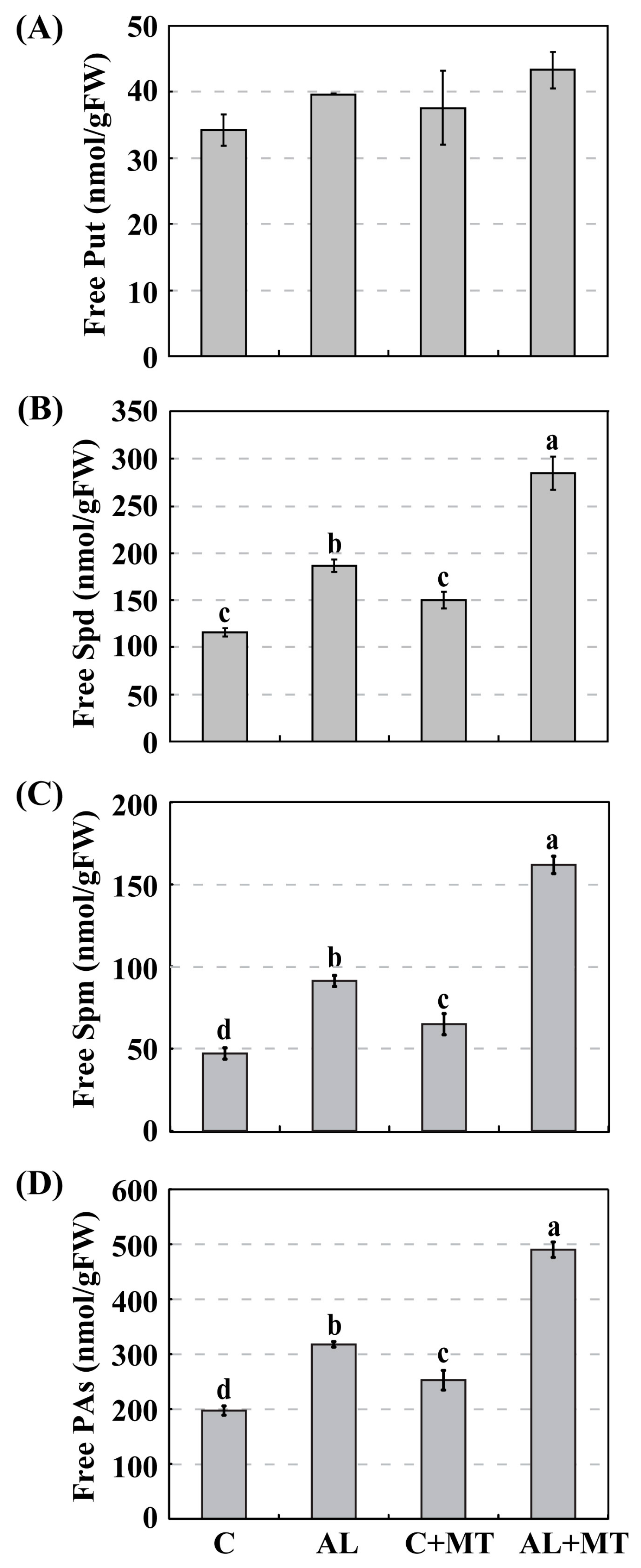
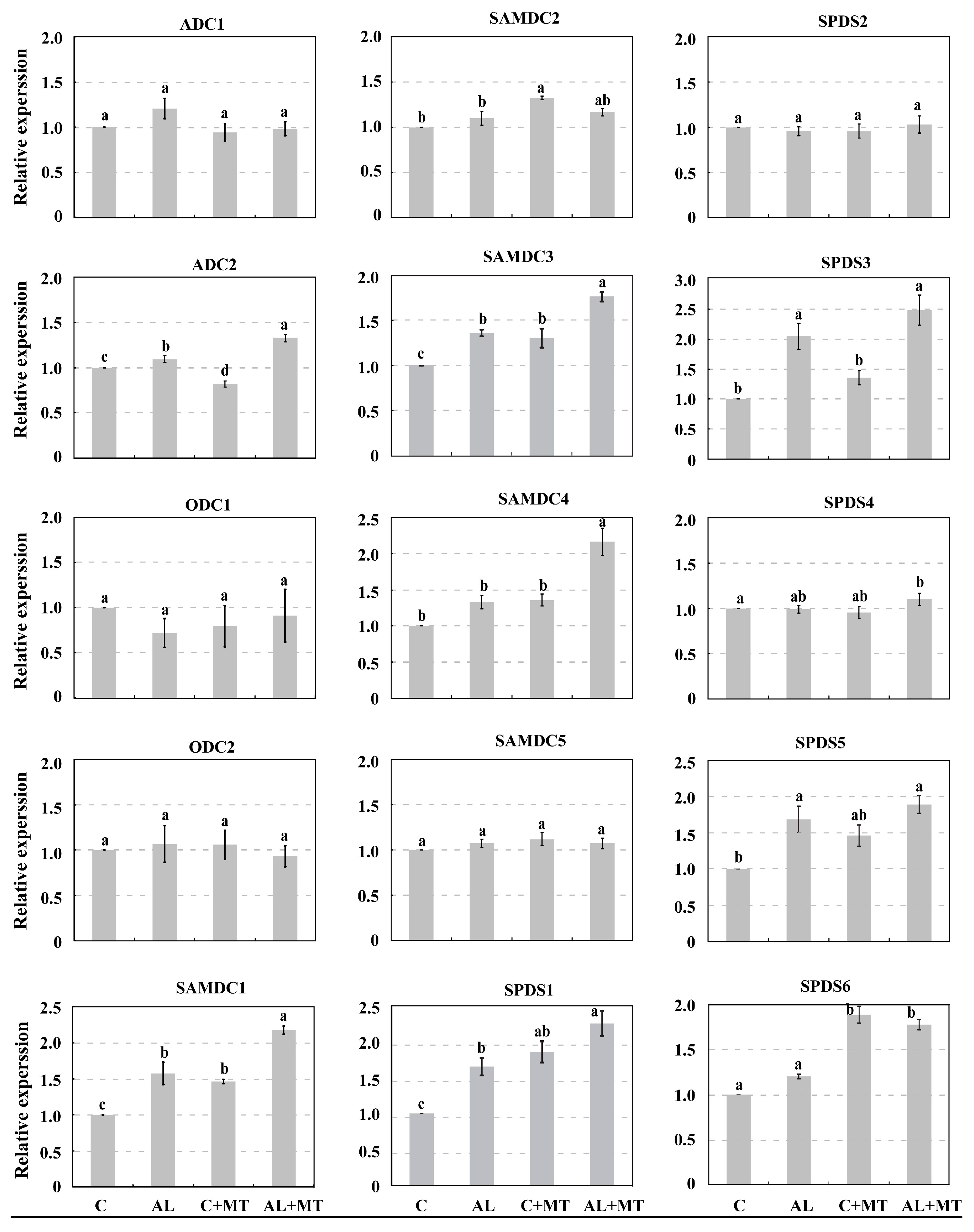
2.4. The Effect of Melatonin on Polyamines Biosynthesis in Malus hupehensis Rehd. under Alkaline Stress
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Melatonin Applications, and Alkaline Stress Treatment
4.2. Assessment of Growth Parameters
4.3. Measurements of REL, Chlorophyll, and Root Activity
4.4. Determination of MDA and ROS, and Activity of Antioxidant Enzymes
4.5. Quantification of Free Polyamines
4.6. Quantitative PCR Analysis
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, W.; Cui, P.; Sun, H.; Guo, W.; Yang, C.; Jin, H.; Fang, B.; Shi, D. Comparative effects of salt and alkali stresses on organic acid accumulation and ionic balance of seabuckthorn (Hippophae rhamnoides L.). Ind. Crops Prod. 2009, 30, 351–358. [Google Scholar] [CrossRef]
- Hu, G.; Liu, Y.; Zhang, X.; Yao, F.; Huang, Y.; Ervin, E.H.; Zhao, B. Physiological evaluation of alkali-salt tolerance of thirty switchgrass (Panicum virgatum) lines. PLoS ONE 2015, 10, e0125305. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Chen, S.; Wang, T.; Sun, G.; Dai, S. Comparative proteomic analysis of Puccinellia tenuiflora leaves under Na2CO3 stress. Int. J. Mol. Sci. 2013, 14, 1740–1762. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Huang, W.; Chen, H.; Wu, G.; Yuan, H.; Song, X.; Kang, Q.; Zhao, D.; Jiang, W.; Liu, Y.; et al. Identification of differentially expressed genes in flax (Linum usitatissimum L.) under saline-alkaline stress by digital gene expression. Gene 2014, 549, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Suo, J.; Chen, S.; Jin, Y.; Ma, X.; Yin, Z.; Zhang, Y.; Wang, T.; Luo, J.; Jin, W.; et al. Na2CO3-responsive mechanisms in halophyte puccinellia tenuiflora roots revealed by physiological and proteomic analyses. Sci. Rep. 2016, 6, 32717. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Genetic analysis of plant salt tolerance using arabidopsis. Plant Physiol. 2000, 124, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Bassil, E.; Coku, A.; Blumwald, E. Cellular ion homeostasis: Emerging roles of intracellular nhx Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 2012, 63, 5727–5740. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Reactive oxygen species as signals that modulate plant stress responses and programmed cell death. Bioessays 2006, 28, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Swanson, S.; Gilroy, S. Ros in plant development. Physiol. Plant. 2010, 138, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.-S.; Wang, W.; Zhang, Q.; Liu, J.-H. A basic helix-loop-helix transcription factor, PtrbHLH, of Poncirus trifoliata confers cold tolerance and modulates peroxidase-mediated scavenging of hydrogen peroxide. Plant Physiol. 2013, 162, 1178–1194. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wu, B.; Yin, H.; Fan, Z.; Li, X.; Ni, S.; He, L.; Li, J. Overexpression of caapx induces orchestrated reactive oxygen scavenging and enhances cold and heat tolerances in tobacco. BioMed Res. Int. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Shigeoka, S. Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 2011, 155, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Moschou, P.N.; Wu, J.; Cona, A.; Tavladoraki, P.; Angelini, R.; Roubelakis-Angelakis, K.A. The polyamines and their catabolic products are significant players in the turnover of nitrogenous molecules in plants. J. Exp. Bot. 2012, 63, 5003–5015. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.T.; Chan, Z.L. Improvement of plant abiotic stress tolerance through modulation of the polyamine pathway. J. Integr. Plant Biol. 2014, 56, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.Q.; Zhang, J.Y.; Hu, J.B.; Wang, W.; Wu, H.; Zhang, Q.H.; Liu, J.H. FcWRKY70, a WRKY protein of Fortunella crassifolia, functions in drought tolerance and modulates putrescine synthesis by regulating arginine decarboxylase gene. Plant Cell Environ. 2015, 38, 2248–2262. [Google Scholar] [CrossRef] [PubMed]
- Sang, Q.Q.; Shan, X.; An, Y.H.; Shu, S.; Sun, J.; Guo, S.R. Proteomic analysis reveals the positive effect of exogenous spermidine in tomato seedlings’ response to high-temperature stress. Front. Plant Sci. 2017, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Koc, E.; Islek, C.; Kasko Arici, Y. Spermine and its interaction with proline induce resistance to the root rot pathogen phytophthora capsici in pepper (Capsicum annuum). Hortic. Environ. Biotechnol. 2017, 58, 254–267. [Google Scholar] [CrossRef]
- Saha, J.; Giri, K. Molecular phylogenomic study and the role of exogenous spermidine in the metabolic adjustment of endogenous polyamine in two rice cultivars under salt stress. Gene 2017, 609, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin-a pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [PubMed]
- Vantassel, D.L.; Roberts, N.J.; Oenill, S.D. Melatonin from higher-plants—Isolation and identification of n-acetyl 5-methoxytryptamine. Plant Physiol. 1995, 108, 101. [Google Scholar]
- Arnao, M.B.; Hernandez-Ruiz, J. Functions of melatonin in plants: A review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, P.; Wei, Z.W.; Liang, D.; Liu, C.H.; Yin, L.H.; Jia, D.F.; Fu, M.Y.; Ma, F.W. The mitigation effects of exogenous melatonin on salinity-induced stress in Malus hupehensis. J. Pineal Res. 2012, 53, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Li, X.N.; Tan, D.X.; Jiang, D.; Liu, F.L. Melatonin enhances cold tolerance in drought-primed wild-type and abscisic acid-deficient mutant barley. J. Pineal Res. 2016, 61, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, C.; Chatzimichail, G.; Xenofontos, R.; Pavlou, J.J.; Panagiotou, E.; Christou, A.; Fotopoulos, V. Melatonin systemically ameliorates drought stress-induced damage in Medicago sativa plants by modulating nitro-oxidative homeostasis and proline metabolism. J. Pineal Res. 2017, 62, e12401. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Z.P.; Yu, X.L.; Cui, W.T.; Pan, J.C.; Zhao, G.; Xu, S.; Wang, R.; Shen, W.B. Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microrna-mediated redox homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Jia, J.F.; Xu, Y.; Wang, Y.L.; Hao, J.G.; Li, T.K. Production of transgenic Nicotiana sylvestris plants expressing melatonin synthetase genes and their effect on UV-B-induced DNA damage. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 275–282. [Google Scholar] [CrossRef]
- Liu, N.; Gong, B.; Jin, Z.Y.; Wang, X.F.; Wei, M.; Yang, F.J.; Li, Y.; Shi, Q.H. Sodic alkaline stress mitigation by exogenous melatonin in tomato needs nitric oxide as a downstream signal. J. Plant Physiol. 2015, 186, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Jin, Z.Y.; Wang, S.S.; Gong, B.A.; Wen, D.; Wang, X.F.; Wei, M.; Shi, Q.H. Sodic alkaline stress mitigation with exogenous melatonin involves reactive oxygen metabolism and ion homeostasis in tomato. Sci. Hortic.-Amst. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- Li, C.; Tan, D.X.; Liang, D.; Chang, C.; Jia, D.F.; Ma, F.W. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 2015, 66, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L.L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.P.; Xie, Y.J.; Gu, Q.; Zhao, G.; Zhang, Y.H.; Cui, W.T.; Xu, S.; Wang, R.; Shen, W.B.A. The AtrbohF-dependent regulation of ROS signaling is required for melatonin-induced salinity tolerance in Arabidopsis. Free Radic. Bio. Med. 2017, 108, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Zhang, H.J.; Sun, Q.Q.; Cao, Y.Y.; Li, X.S.; Zhao, B.; Wu, P.; Guo, Y.D. Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production. Sci. Rep. 2017, 7, 503. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.T.; Wang, X.; Tan, D.X.; Reiter, R.J.; Chan, Z.L. Comparative physiological and proteomic analyses reveal the actions of melatonin in the reduction of oxidative stress in Bermuda grass (Cynodon dactylon (L.). Pers.). J. Pineal Res. 2015, 59, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Cuenca, M.R.; Forner-Giner, M.A.; Iglesias, D.J.; Primo-Millo, E.; Legaz, F. Strategy I responses to Fe-deficiency of two Citrus rootstocks differing in their tolerance to iron chlorosis. Sci. Hortic.-Amst. 2013, 153, 56–63. [Google Scholar] [CrossRef]
- Wang, X.P.; Jiang, P.; Ma, Y.; Geng, S.J.; Wang, S.C.; Shi, D.C. Physiological strategies of sunflower exposed to salt or alkali stresses: Restriction of ion transport in the cotyledon node zone and solute accumulation. Agron. J. 2015, 107, 2181–2192. [Google Scholar] [CrossRef]
- Shi, H.T.; Tan, D.X.; Reiter, R.J.; Ye, T.T.; Yang, F.; Chan, Z.L. Melatonin induces class A1 heat-shock factors (HSFA1s) and their possible involvement of thermotolerance in Arabidopsis. J. Pineal Res. 2015, 58, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lee, D.E.; Jang, H.; Byeon, Y.; Kim, Y.S.; Back, K. Melatonin-rich transgenic rice plants exhibit resistance to herbicide-induced oxidative stress. J. Pineal Res. 2013, 54, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.J.; Chen, H.J.; Wang, Z.Y.; Gu, X.R.; Wei, C.H.; Zhang, Y.; Ma, J.X.; Yang, J.Q.; Zhang, X. Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Front. Plant Sci. 2017, 8, 295. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; David, A.; Yadav, S.; Baluska, F.; Bhatla, S.C. Salt stress-induced seedling growth inhibition coincides with differential distribution of serotonin and melatonin in sunflower seedling roots and cotyledons. Physiol. Plant. 2014, 152, 714–728. [Google Scholar] [CrossRef] [PubMed]
- Osmont, K.S.; Sibout, R.; Hardtke, C.S. Hidden branches: Developments in root system architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef] [PubMed]
- Kuijken, R.C.; van Eeuwijk, F.A.; Marcelis, L.F.; Bouwmeester, H.J. Root phenotyping: From component trait in the lab to breeding. J. Exp. Bot. 2015, 66, 5389–5401. [Google Scholar] [CrossRef] [PubMed]
- Chapman, N.; Whalley, W.R.; Lindsey, K.; Miller, A.J. Water supply and not nitrate concentration determines primary root growth in Arabidopsis. Plant Cell Environ. 2011, 34, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Uga, Y.; Sugimoto, K.; Ogawa, S.; Rane, J.; Ishitani, M.; Hara, N.; Kitomi, Y.; Inukai, Y.; Ono, K.; Kanno, N.; et al. Control of root system architecture by deeper rooting 1 increases rice yield under drought conditions. Nat. Genet. 2013, 45, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yang, A.; Zhang, W.H. Efficient acquisition of iron confers greater tolerance to saline-alkaline stress in rice (Oryza sativa L.). J. Exp. Bot. 2016, 67, 6431–6444. [Google Scholar] [CrossRef] [PubMed]
- Campestre, M.P.; Antonelli, C.; Calzadilla, P.I.; Maiale, S.J.; Rodriguez, A.A.; Ruiz, O.A. The alkaline tolerance in Lotus japonicus is associated with mechanisms of iron acquisition and modification of the architectural pattern of the root. J. Plant Physiol. 2016, 206, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.M.; Zhang, Y.Q. Melatonin: A well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.T.; Zhao, H.L.; Cao, K.; Hu, L.P.; Du, T.H.; Baluska, F.; Zou, Z.R. Beneficial roles of melatonin on redox regulation of photosynthetic electron transport and synthesis of D1 protein in tomato seedlings under salt stress. Front. Plant Sci. 2016, 7, 1823. [Google Scholar] [CrossRef] [PubMed]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin as a natural ally against oxidative stress: A physicochemical examination. J. Pineal Res. 2011, 51, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Sun, Q.Q.; Zhang, H.J.; Cao, Y.Y.; Weeda, S.; Ren, S.X.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Back, K. 2-hydroxymelatonin promotes the resistance of rice plant to multiple simultaneous abiotic stresses (combined cold and drought). J. Pineal Res. 2016, 61, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.J.; Zheng, J.X.; Dong, Y.C.; Liu, Q.Y.; Yang, X.Z.; Wei, C.H.; Zhang, Y.; Ma, J.X.; Zhang, X. Local melatonin application induces cold tolerance in distant organs of Citrullus lanatus L. via long distance transport. Sci. Rep. 2017, 7, 40858. [Google Scholar] [CrossRef] [PubMed]
- Dalton, H.L.; Blomstedt, C.K.; Neale, A.D.; Gleadow, R.; DeBoer, K.D.; Hamill, J.D. Effects of down-regulating ornithine decarboxylase upon putrescine-associated metabolism and growth in Nicotiana tabacum L. J. Exp. Bot. 2016, 67, 3367–3381. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Fu, B.; Sun, P.P.; Xiao, C.; Liu, J.H. A NAC transcription factor represses putrescine biosynthesis and affects drought tolerance. Plant Physiol. 2016, 172, 1532–1547. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Li, X.; Bloszies, S.; Wen, D.; Sun, S.S.; Wei, M.; Li, Y.; Yang, F.J.; Shi, Q.H.; Wang, X.F. Sodic alkaline stress mitigation by interaction of nitric oxide and polyamines involves antioxidants and physiological strategies in Solanum lycopersicum. Free Radic. Biol. Med. 2014, 71, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, Z.; Zhu, L.; Ma, Z.Y.; Wang, J.F.; Zhu, J. Exogenous melatonin improves plant iron deficiency tolerance via increased accumulation of polyamine-mediated nitric oxide. Int. J. Mol. Sci. 2016, 17, 1777. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.; Gong, B.; Sun, S.; Liu, S.; Wang, X.; Wei, M.; Yang, F.; Li, Y.; Shi, Q. Promoting roles of melatonin in adventitious root development of Solanum lycopersicum L. by regulating auxin and nitric oxide signaling. Front. Plant Sci. 2016, 7, 718. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.Y.; Zhu, R.Y.; Zhang, G.Y.; Dai, Y.R. Attenuation of cold-induced apoptosis by exogenous melatonin in carrot suspension cells: The possible involvement of polyamines. J. Pineal Res. 2004, 36, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhang, K.; Zhou, X.; Xi, L.; Wang, Y.; Xu, H.; Pan, T.; Zou, Z. Melatonin alleviates chilling stress in cucumber seedlings by up-regulation of CsZat12 and modulation of polyamine and abscisic acid metabolism. Sci. Rep. 2017, 7, 4998. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Song, C.; Shao, J.; Bian, K.; Chen, W.; Yang, Z. Exogenous melatonin treatment increases chilling tolerance and induces defense response in harvested peach fruit during cold storage. J. Agric. Food Chem. 2016, 64, 5215–5222. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Sun, P.P.; Chen, C.L.; Wang, Y.; Fu, X.Z.; Liu, J.H. An arginine decarboxylase gene PtADC from Poncirus trifoliata confers abiotic stress tolerance and promotes primary root growth in Arabidopsis. J. Exp. Bot. 2011, 62, 2899–2914. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Measurement of plant root activity (ttc). In Principles and Techniques of Plant Physiological Biochemical Experiment, 1st ed.; Li, H.S., Ed.; Higher Education Press: Beijing, China, 2000; pp. 119–120. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Gong, X.; Liu, J.H. Detection of free polyamines in plants subjected to abiotic stresses by high-performance liquid chromatography (HPLC). Methods Mol. Biol. 2017, 1631, 305–311. [Google Scholar] [PubMed]
- Perini, P.; Pasquali, G.; Margis-Pinheiro, M.; de Oliviera, P.R.; Revers, L. Reference genes for transcriptional analysis of flowering and fruit ripening stages in apple (Malus × domestica Borkh.). Mol. Breed. 2014, 34, 829–842. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shoot Height (M ± SD, cm) | Leaf Number (M ± SD) | Stem Diameter (M ± SD, mm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | |
| C | 10.77 ± 0.23 bc (100%) | 13.02 ± 0.28 a (100%) | 20.9% | 10.67 ± 0.35 c (100%) | 13.87 ± 0.19 a (100%) | 30.0% | 1.88 ± 0.04 b (100%) | 2.13 ± 0.04 a (100%) | 13.3% |
| AL | 10.45 ± 0.30 c (97.0%) | 11.57 ± 0.27 b (88.7%) | 10.7% | 10.67 ± 0.35 c (100%) | 12.20 ± 0.24 b (88.0%) | 14.3% | 1.88 ± 0.03 b (100%) | 1.90 ± 0.05 b (87.3%) | 1.6% |
| C + MT | 10.97 ± 0.48 bc (100%) | 13.15 ± 0.32 a (100%) | 19.9% | 11.33 ± 0.33 c (100%) | 13.93 ± 0.21 a (100%) | 23.0% | 1.90 ± 0.07 b (100%) | 2.16 ± 0.04 a (100%) | 13.7% |
| AL + MT | 10.95 ± 0.39 bc (99.8%) | 13.09 ± 0.43 a (99.5%) | 19.5% | 11.20 ± 0.47 c (98.9%) | 13.80 ± 0.17 a (99.1%) | 23.2% | 1.89 ± 0.04 b (99.5%) | 2.11 ± 0.03 a (97.7%) | 11.6% |
| Root (M ± SD) | Stem (M ± SD) | Leaf (M ± SD) | Total (M ± SD) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | |
| C | 0.5018 ± 0.0248 b (100%) | 0.8124 ± 0.03 a (100%) | 61.9% | 0.2486 ± 0.0097 c (100%) | 0.3382 ± 0.0159 ab (100%) | 36.0% | 0.7468 ± 0.0332 c (100%) | 1.2612 ± 0.0523 ab (100%) | 68.9% | 1.4972 (100%) | 2.4118 (100%) | 61.1% |
| AL | 0.5091 ± 0.0364 b (101.5%) | 0.5794 ± 0.0119 b (71.3%) | 13.8% | 0.2429 ± 0.0145 c (97.7%) | 0.2460 ± 0.0127 c (72.7%) | 1.3% | 0.7401 ± 0.0491 c (99.1%) | 0.8650 ± 0.0278 c (68.6%) | 16.9% | 1.4921 (99.7%) | 1.6903 (70.1%) | 13.3% |
| C + MT | 0.5329 ± 0.0302 b (100%) | 0.8571 ± 0.0234 a (100%) | 60.8% | 0.2581 ± 0.0201 c (100%) | 0.3538 ± 0.0124 a (100%) | 37.1% | 0.7830 ± 0.0470 c (100%) | 1.3722 ± 0.0629 a (100%) | 75.2% | 1.5740 (100%) | 2.5831 (100%) | 64.1% |
| AL + MT | 0.5431 ± 0.0385 b (101.9%) | 0.8337 ± 0.0225 a (97.3%) | 53.5% | 0.2538 ± 0.0182 c (98.3%) | 0.3048 ± 0.0043 b (86.2%) | 20.1% | 0.7939 ± 0.0502 c (101.4%) | 1.1581 ± 0.0296 b (84.4%) | 45.9% | 1.5906 (101.1%) | 2.2966 (88.9%) | 44.4% |
| Root (M ± SD) | Stem (M ± SD) | Leaf (M ± SD) | Total (M ± SD) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | 0 Day | 15 Days | Relative Growth | |
| C | 0.1175 ± 0.005 b (100%) | 0.1801 ± 0.0092 a (100%) | 53.3% | 0.1042 ± 0.0035 b (100%) | 0.1265 ± 0.0067 a (100%) | 21.4% | 0.2488 ± 0.0118 b (100%) | 0.3950 ± 0.0192 a (100%) | 58.8% | 0.4705 (100%) | 0.7017 (100%) | 49.1% |
| AL | 0.1163 ± 0.007 b (99.0%) | 0.1360 ± 0.0043 b (75.5%) | 16.9% | 0.1036 ± 0.0060 b (99.4%) | 0.0989 ± 0.0047 b (78.2%) | −5.5% | 0.2404 ± 0.0159 b (96.6%) | 0.2792 ± 0.0092 b (70.1%) | 16.1% | 0.4603 (97.8%) | 0.5141 (73.3%) | 11.7% |
| C + MT | 0.1150 ± 0.007 b (100%) | 0.1908 ± 0.0083 a (100%) | 65.9% | 0.1032 ± 0.0077 b (100%) | 0.1427 ± 0.0066 a (100%) | 38.3% | 0.2544 ± 0.0177 b (100%) | 0.3974 ± 0.0109 a (100%) | 56.2% | 0.4726 (100%) | 0.7309 (100%) | 54.7% |
| AL + MT | 0.1160 ± 0.008 b (100.9%) | 0.1796 ± 0.0047 a (94.1%) | 54.8% | 0.1060 ± 0.0074 b (102.7%) | 0.1257 ± 0.0022 a (88.1%) | 18.6% | 0.2568 ± 0.0204 b (100.9%) | 0.3579 ± 0.0070 a (90.1%) | 39.4% | 0.4787 (101.3%) | 0.6631 (90.7%) | 38.5% |
| Length (cm) | Surface Area (cm2) | Avg. Diam (mm) | Root Volume (cm3) | No. of Tips * | No. of Forks * | |
|---|---|---|---|---|---|---|
| C | 244.30 ± 14.93 a (100%) | 31.63 ± 2.25 ab (100%) | 0.44 ± 0.02 ab (100%) | 0.36 ± 0.04 ab (100%) | 1577 ± 155 ab (100%) | 2275 ± 144 ab (100%) |
| AL | 220.46 ± 12.94 a (90.2%) | 27.13 ± 1.69 b (85.8%) | 0.38 ± 0.01 b (86.4%) | 0.29 ± 0.02 b (80.6%) | 1179 ± 100 b (74.8%) | 2002 ± 140 b (88.0%) |
| C + MT | 267.66 ± 21.53 a (100%) | 35.75 ± 1.36 a (100%) | 0.50 ± 0.02 a (100%) | 0.45 ± 0.01 ab (100%) | 1944 ± 186 a (100%) | 2956 ± 301 a (100%) |
| AL + MT | 241.41 ± 20.45 a (90.2%) | 31.96 ± 2.21 ab (89.4%) | 0.43 ± 0.03 ab (86.0%) | 0.46 ± 0.07 a (102.2%) | 1822 ± 196 a (93.7%) | 2773 ± 253 ab (93.8%) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Shi, S.; Dou, F.; Song, Y.; Ma, F. Exogenous Melatonin Alleviates Alkaline Stress in Malus hupehensis Rehd. by Regulating the Biosynthesis of Polyamines. Molecules 2017, 22, 1542. https://doi.org/10.3390/molecules22091542
Gong X, Shi S, Dou F, Song Y, Ma F. Exogenous Melatonin Alleviates Alkaline Stress in Malus hupehensis Rehd. by Regulating the Biosynthesis of Polyamines. Molecules. 2017; 22(9):1542. https://doi.org/10.3390/molecules22091542
Chicago/Turabian StyleGong, Xiaoqing, Shuting Shi, Fangfang Dou, Yi Song, and Fengwang Ma. 2017. "Exogenous Melatonin Alleviates Alkaline Stress in Malus hupehensis Rehd. by Regulating the Biosynthesis of Polyamines" Molecules 22, no. 9: 1542. https://doi.org/10.3390/molecules22091542