In Vitro Innervation as an Experimental Model to Study the Expression and Functions of Acetylcholinesterase and Agrin in Human Skeletal Muscle


 ,
, 
Abstract
:1. Introduction
2. The Experimental Model of the in Vitro Innervated Human Skeletal Muscle Cells
2.1. The Development of Skeletal Muscle is More Nerve-Dependent in Humans than in Animal Species Used in Skeletal Muscle Research
2.1.1. Postsynaptic NMJ Components in Aneurally Cultured Human and Animal Skeletal Muscle Cells
2.1.2. Contractile Properties of Aneurally Cultured Human and Animal Skeletal Muscle Cells
2.1.3. Evolutionary Basis for Differences in Developmental Programme of Skeletal Muscle Cells?
2.2. Experimental Model of the in Vitro Innervated Human Skeletal Muscle Cells: Preparation and Description
2.2.1. Co-Culture Models to Study NMJ
2.2.2. Co-cultures of Primary Human Skeletal Muscle Cells and Rat Embryonic Spinal Cord: Basic Characteristics
2.2.3. Developmental Characteristics of the in Vitro Innervated Human Skeletal Muscle Cells
2.2.4. Challenges and Limitations of Using the Coventional Rat-Human Co-Culture Model
2.3. Characterization of the Neural Component of the Co-Cultures of Human Skeletal Muscle Cells and Rat Spinal Cord Explants
2.3.1. The Essential Role of Glial Cells and Sensory Neurons
2.3.2. Neural Networks and Spontaneous Neural Activity in Spinal Cord Explants
2.4. Formation of the Basal Lamina in the Synaptic Cleft coincides with the Transformation of Immature Neuromuscular Contact into Differentiated NMJ
3. The Expression of AChE during NMJ Formation in Co-Cultures of Human Skeletal Muscle Cells and Embryonic Rat Spinal Cord Explants.
4. The Role of Neural Agrin in the Formation of the NMJ in the in Vitro Innervated Human Skeletal Muscle Cells
5. Possible Alternative Roles of AChE and Neural Agrin in Human Skeletal Muscle
5.1. AChE and Apoptosis of Primary Human Myoblasts
5.2. Neural Agrin and Differentiation of Cultured Human Skeletal Muscle Cells
6. Conclusions and Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Burden, S.J.; Yumoto, N.; Zhang, W. The role of MuSK in synapse formation and neuromuscular disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a009167. [Google Scholar] [CrossRef] [PubMed]
- Rotundo, R.L.; Rossi, S.G.; Kimbell, L.M.; Ruiz, C.; Marrero, E. Targeting acetylcholinesterase to the neuromuscular synapse. Chem. Biol. Interact. 2005, 157–158, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Rotundo, R.L. Biogenesis, assembly and trafficking of acetylcholinesterase. J. Neurochem. 2017. [Google Scholar] [CrossRef] [PubMed]
- McMahan, U.J. The agrin hypothesis. Cold Spring Harb. Symp. Quant. Biol. 1990, 55, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Massoulie, J.; Millard, C.B. Cholinesterases and the basal lamina at vertebrate neuromuscular junctions. Curr. Opin. Pharmacol. 2009, 9, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Kummer, T.T.; Misgeld, T.; Sanes, J.R. Assembly of the postsynaptic membrane at the neuromuscular junction: Paradigm lost. Curr. Opin. Neurobiol. 2006, 16, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Bezakova, G.; Ruegg, M.A. New insights into the roles of agrin. Nat. Rev. Mol. Cell. Biol. 2003, 4, 295–308. [Google Scholar] [CrossRef]
- Mis, K.; Matkovic, U.; Pirkmajer, S.; Sciancalepore, M.; Lorenzon, P.; Mars, T.; Grubic, Z. Acetylcholinesterase and agrin: Different functions, similar expression patterns, multiple roles. Chem. Biol. Interact. 2013, 203, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Gros, K.; Parato, G.; Pirkmajer, S.; Mis, K.; Podbregar, M.; Grubic, Z.; Lorenzon, P.; Mars, T. Non-synaptic roles of acetylcholinesterase and agrin. J. Mol. Neurosci. 2014, 53, 454–460. [Google Scholar] [CrossRef]
- Campoy, F.J.; Vidal, C.J.; Munoz-Delgado, E.; Montenegro, M.F.; Cabezas-Herrera, J.; Nieto-Ceron, S. Cholinergic system and cell proliferation. Chem. Biol. Interact. 2016, 259, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Greenberg, D.S. Acetylcholinesterase involvement in apoptosis. Front. Mol. Neurosci. 2012, 5. [Google Scholar] [CrossRef] [PubMed]
- Rezonja, K.; Mars, T.; Jerin, A.; Kozelj, G.; Pozar-Lukanovic, N.; Sostaric, M. Dexamethasone does not diminish sugammadex reversal of neuromuscular block—Clinical study in surgical patients undergoing general anesthesia. BMC Anesthesiol. 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Krivoi, I.I. Mechanisms of the non-neurotransmitter actions of acetylcholine in the neuromuscular apparatus. Neurosci. Behav. Physiol. 2002, 32, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Heiny, J.A.; Kravtsova, V.V.; Mandel, F.; Radzyukevich, T.L.; Benziane, B.; Prokofiev, A.V.; Pedersen, S.E.; Chibalin, A.V.; Krivoi, I.I. The nicotinic acetylcholine receptor and the na,k-atpase alpha2 isoform interact to regulate membrane electrogenesis in skeletal muscle. J. Biol. Chem. 2010, 285, 28614–28626. [Google Scholar] [CrossRef]
- Kravtsova, V.V.; Petrov, A.M.; Matchkov, V.V.; Bouzinova, E.V.; Vasiliev, A.N.; Benziane, B.; Zefirov, A.L.; Chibalin, A.V.; Heiny, J.A.; Krivoi, I.I. Distinct alpha2 Na,K-ATPase membrane pools are differently involved in early skeletal muscle remodeling during disuse. J. Gen. Physiol. 2016, 147, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Zierath, J.R.; Galuska, D.; Engstrom, A.; Johnson, K.H.; Betsholtz, C.; Westermark, P.; Wallberg-Henriksson, H. Human islet amyloid polypeptide at pharmacological levels inhibits insulin and phorbol ester-stimulated glucose transport in in vitro incubated human muscle strips. Diabetologia 1992, 35, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, H.K.; Chibalin, A.V.; Koistinen, H.A.; Yang, J.; Koumanov, F.; Wallberg-Henriksson, H.; Zierath, J.R.; Holman, G.D. Kinetics of glut4 trafficking in rat and human skeletal muscle. Diabetes 2009, 58, 847–854. [Google Scholar] [CrossRef] [PubMed]
- Zierath, J.R.; Krook, A.; Wallberg-Henriksson, H. Insulin action and insulin resistance in human skeletal muscle. Diabetologia 2000, 43, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Dohm, G.L.; Tapscott, E.B.; Pories, W.J.; Dabbs, D.J.; Flickinger, E.G.; Meelheim, D.; Fushiki, T.; Atkinson, S.M.; Elton, C.W.; Caro, J.F. An in vitro human muscle preparation suitable for metabolic studies. Decreased insulin stimulation of glucose transport in muscle from morbidly obese and diabetic subjects. J. Clin. Investig. 1988, 82, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.E.; Dohm, G.L.; Leggett-Frazier, N.; Elton, C.W.; Tapscott, E.B.; Pories, W.P.; Caro, J.F. Restoration of insulin responsiveness in skeletal muscle of morbidly obese patients after weight loss. Effect on muscle glucose transport and glucose transporter glut4. J. Clin. Investig. 1992, 89, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Aas, V.; Bakke, S.S.; Feng, Y.Z.; Kase, E.T.; Jensen, J.; Bajpeyi, S.; Thoresen, G.H.; Rustan, A.C. Are cultured human myotubes far from home? Cell Tissue Res. 2013, 354, 671–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaporte, C.; Dautreaux, B.; Fardeau, M. Human myotube differentiation in vitro in different culture conditions. Biol. Cell 1986, 57, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Askanas, V. Acetylcholine receptors and acetylcholinesterase accumulate at the nerve-muscle contacts of de novo grown human monolayer muscle cocultured with fetal rat spinal cord. Exp. Neurol. 1985, 88, 327–335. [Google Scholar] [CrossRef]
- Askanas, V.; Kwan, H.; Alvarez, R.B.; Engel, W.K.; Kobayashi, T.; Martinuzzi, A.; Hawkins, E.F. De novo neuromuscular junction formation on human muscle fibres cultured in monolayer and innervated by foetal rat spinal cord: Ultrastructural and ultrastructural-cytochemical studies. J. Neurocytol. 1987, 16, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Askanas, V.; Engel, W.K. Human muscle cultured in monolayer and cocultured with fetal rat spinal cord: Importance of dorsal root ganglia for achieving successful functional innervation. J. Neurosci. 1987, 7, 3131–3141. [Google Scholar]
- Couteaux, R. Early days in the research to localize skeletal muscle acetylcholinesterases. J. Physiol. Paris 1998, 92, 59–62. [Google Scholar] [CrossRef]
- Grubic, Z.; Komel, R.; Walker, W.F.; Miranda, A.F. Myoblast fusion and innervation with rat motor nerve alter distribution of acetylcholinesterase and its mrna in cultures of human muscle. Neuron 1995, 14, 317–327. [Google Scholar] [CrossRef]
- Sanes, J.R.; Lawrence, J.C., Jr. Activity-dependent accumulation of basal lamina by cultured rat myotubes. Dev. Biol. 1983, 97, 123–136. [Google Scholar] [CrossRef]
- Sanes, J.R. The basement membrane/basal lamina of skeletal muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef] [PubMed]
- Vogel, Z.; Sytkowski, A.J.; Nirenberg, M.W. Acetylcholine receptors of muscle grown in vitro. Proc. Natl. Acad. Sci. USA 1972, 69, 3180–3184. [Google Scholar] [CrossRef] [PubMed]
- Sytkowski, A.J.; Vogel, Z.; Nirenberg, M.W. Development of acetylcholine receptor clusters on cultured muscle cells. Proc. Natl. Acad. Sci. USA 1973, 70, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Axelrod, D.; Ravdin, P.; Koppel, D.E.; Schlessinger, J.; Webb, W.W.; Elson, E.L.; Podleski, T.R. Lateral motion of fluorescently labeled acetylcholine receptors in membranes of developing muscle fibers. Proc. Natl. Acad. Sci. USA 1976, 73, 4594–4598. [Google Scholar] [CrossRef] [PubMed]
- Christian, C.N.; Daniels, M.P.; Sugiyama, H.; Vogel, Z.; Jacques, L.; Nelson, P.G. A factor from neurons increases the number of acetylcholine receptor aggregates on cultured muscle cells. Proc. Natl. Acad. Sci. USA 1978, 75, 4011–4015. [Google Scholar] [CrossRef] [PubMed]
- Kummer, T.T.; Misgeld, T.; Lichtman, J.W.; Sanes, J.R. Nerve-independent formation of a topologically complex postsynaptic apparatus. J. Cell Biol. 2004, 164, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Gajsek, N.; Jevsek, M.; Mars, T.; Mis, K.; Pirkmajer, S.; Brecelj, J.; Grubic, Z. Synaptogenetic mechanisms controlling postsynaptic differentiation of the neuromuscular junction are nerve-dependent in human and nerve-independent in mouse C2C12 muscle cultures. Chem. Biol. Interact. 2008, 175, 50–57. [Google Scholar] [CrossRef]
- DeChiara, T.M.; Bowen, D.C.; Valenzuela, D.M.; Simmons, M.V.; Poueymirou, W.T.; Thomas, S.; Kinetz, E.; Compton, D.L.; Rojas, E.; Park, J.S.; et al. The receptor tyrosine kinase MuSK is required for neuromuscular junction formation in vivo. Cell 1996, 85, 501–512. [Google Scholar] [CrossRef]
- Sanes, J.R.; Lichtman, J.W. Induction, assembly, maturation and maintenance of a postsynaptic apparatus. Nat. Rev. Neurosci. 2001, 2, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Burden, S.J. Building the vertebrate neuromuscular synapse. J. Neurobiol. 2002, 53, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Watty, A.; Neubauer, G.; Dreger, M.; Zimmer, M.; Wilm, M.; Burden, S.J. The in vitro and in vivo phosphotyrosine map of activated MuSK. Proc. Natl. Acad. Sci. USA 2000, 97, 4585–4590. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.; Herbst, R. The formation of complex acetylcholine receptor clusters requires MuSK kinase activity and structural information from the MuSK extracellular domain. Mol. Cell. Neurosci. 2012, 49, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Apel, E.D.; Glass, D.J.; Moscoso, L.M.; Yancopoulos, G.D.; Sanes, J.R. Rapsyn is required for MuSK signaling and recruits synaptic components to a musk-containing scaffold. Neuron 1997, 18, 623–635. [Google Scholar] [CrossRef]
- Mars, T.; King, M.P.; Miranda, A.F.; Walker, W.F.; Mis, K.; Grubic, Z. Functional innervation of cultured human skeletal muscle proceeds by two modes with regard to agrin effects. Neuroscience 2003, 118, 87–97. [Google Scholar] [CrossRef]
- Kim, N.; Stiegler, A.L.; Cameron, T.O.; Hallock, P.T.; Gomez, A.M.; Huang, J.H.; Hubbard, S.R.; Dustin, M.L.; Burden, S.J. Lrp4 is a receptor for agrin and forms a complex with musk. Cell 2008, 135, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Luo, S.; Wang, Q.; Suzuki, T.; Xiong, W.C.; Mei, L. Lrp4 serves as a coreceptor of agrin. Neuron 2008, 60, 285–297. [Google Scholar] [CrossRef] [PubMed]
- Guettier-Sigrist, S.; Coupin, G.; Warter, J.M.; Poindron, P. Cell types required to efficiently innervate human muscle cells in vitro. Exp. Cell Res. 2000, 259, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mars, T.; Yu, K.J.; Tang, X.M.; Miranda, A.F.; Grubic, Z.; Cambi, F.; King, M.P. Differentiation of glial cells and motor neurons during the formation of neuromuscular junctions in cocultures of rat spinal cord explant and human muscle. J. Comp. Neurol. 2001, 438, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Nikolic, N.; Bakke, S.S.; Kase, E.T.; Rudberg, I.; Flo Halle, I.; Rustan, A.C.; Thoresen, G.H.; Aas, V. Electrical pulse stimulation of cultured human skeletal muscle cells as an in vitro model of exercise. PLoS ONE 2012, 7, e33203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolic, N.; Gorgens, S.W.; Thoresen, G.H.; Aas, V.; Eckel, J.; Eckardt, K. Electrical pulse stimulation of cultured skeletal muscle cells as a model for in vitro exercise—Possibilities and limitations. Acta Physiol. 2017, 220, 310–331. [Google Scholar] [CrossRef] [PubMed]
- Lambernd, S.; Taube, A.; Schober, A.; Platzbecker, B.; Gorgens, S.W.; Schlich, R.; Jeruschke, K.; Weiss, J.; Eckardt, K.; Eckel, J. Contractile activity of human skeletal muscle cells prevents insulin resistance by inhibiting pro-inflammatory signalling pathways. Diabetologia 2012, 55, 1128–1139. [Google Scholar] [CrossRef] [PubMed]
- Steinbeck, J.A.; Jaiswal, M.K.; Calder, E.L.; Kishinevsky, S.; Weishaupt, A.; Toyka, K.V.; Goldstein, P.A.; Studer, L. Functional connectivity under optogenetic control allows modeling of human neuromuscular disease. Cell Stem Cell 2016, 18, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Spector, I.; Prives, J.M. Development of electrophysiological and biochemical membrane properties during differentiation of embryonic skeletal muscle in culture. Proc. Natl. Acad. Sci. USA 1977, 74, 5166–5170. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Fischman, D.A.; Moscona, A.A. The fine structure of embryonic chick skeletal muscle cells differentiated in vitro. J. Cell Biol. 1967, 35, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ezerman, E.B.; Ishikawa, H. Differentiation of the sarcoplasmic reticulum and T system in developing chick skeletal muscle in vitro. J. Cell Biol. 1967, 35, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Konigsberg, I.R. Clonal analysis of myogenesis. Science 1963, 140, 1273–1284. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.G.; Rotundo, R.L. Cell surface acetylcholinesterase molecules on multinucleated myotubes are clustered over the nucleus of origin. J. Cell Biol. 1992, 119, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Valle, C.; Rotundo, R.L. Regulation of acetylcholinesterase synthesis and assembly by muscle activity. Effects of tetrodotoxin. J. Biol. Chem. 1989, 264, 14043–14049. [Google Scholar]
- Tang, H.; Sun, Z.; Goldman, D. Cam kinase II-dependent suppression of nicotinic acetylcholine receptor delta-subunit promoter activity. J. Biol. Chem. 2001, 276, 26057–26065. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D. Retention of differentiation potentialities during prolonged cultivation of myogenic cells. Proc. Natl. Acad. Sci. USA 1968, 61, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Brockman, S.K.; Przybylski, R.J.; Younkin, S.G. Cellular localization of the molecular forms of acetylcholinesterase in cultured embryonic rat myotubes. J. Neurosci. 1982, 2, 1775–1785. [Google Scholar] [PubMed]
- Brockman, S.K.; Younkin, L.H.; Younkin, S.G. The effect of spontaneous electromechanical activity on the metabolism of acetylcholinesterase in cultured embryonic rat myotubes. J. Neurosci. 1984, 4, 131–140. [Google Scholar] [PubMed]
- Rieger, F.; Koenig, J.; Vigny, M. Spontaneous contractile activity and the presence of the 16 S form of acetylcholinesterase in rat muscle cells in culture: Reversible suppressive action of tetrodotoxin. Dev. Biol. 1980, 76, 358–365. [Google Scholar] [CrossRef]
- Dutton, E.K.; Simon, A.M.; Burden, S.J. Electrical activity-dependent regulation of the acetylcholine receptor delta-subunit gene, MyoD, and myogenin in primary myotubes. Proc. Natl. Acad. Sci. USA 1993, 90, 2040–2044. [Google Scholar] [CrossRef] [PubMed]
- Zahavi, E.E.; Ionescu, A.; Gluska, S.; Gradus, T.; Ben-Yaakov, K.; Perlson, E. A compartmentalized microfluidic neuromuscular co-culture system reveals spatial aspects of GDNF functions. J. Cell Sci. 2015, 128, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Smolina, N.; Kostareva, A.; Bruton, J.; Karpushev, A.; Sjoberg, G.; Sejersen, T. Primary murine myotubes as a model for investigating muscular dystrophy. Biomed. Res. Int. 2015, 2015, 594751. [Google Scholar] [CrossRef] [PubMed]
- Cohen, T.V.; Randall, W.R. The regulation of acetylcholinesterase by cis-elements within intron I in cultured contracting myotubes. J. Neurochem. 2006, 98, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Dutton, E.K.; Uhm, C.S.; Samuelsson, S.J.; Schaffner, A.E.; Fitzgerald, S.C.; Daniels, M.P. Acetylcholine receptor aggregation at nerve-muscle contacts in mammalian cultures: Induction by ventral spinal cord neurons is specific to axons. J. Neurosci. 1995, 15, 7401–7416. [Google Scholar] [PubMed]
- Rubin, L.L.; Chalfin, N.A.; Adamo, A.; Klymkowsky, M.W. Cellular and secreted forms of acetylcholinesterase in mouse muscle cultures. J. Neurochem. 1985, 45, 1932–1940. [Google Scholar] [CrossRef] [PubMed]
- Streit, J. Mechanisms of pattern generation in co-cultures of embryonic spinal cord and skeletal muscle. Int. J. Dev. Neurosci. 1996, 14, 137–148. [Google Scholar] [CrossRef]
- Chahine, K.G.; Baracchini, E.; Goldman, D. Coupling muscle electrical activity to gene expression via a camp-dependent second messenger system. J. Biol. Chem. 1993, 268, 2893–2898. [Google Scholar]
- Chahine, K.G.; Walke, W.; Goldman, D. A 102 base pair sequence of the nicotinic acetylcholine receptor delta-subunit gene confers regulation by muscle electrical activity. Development 1992, 115, 213–219. [Google Scholar] [PubMed]
- Hall, Z.W. Multiple forms of acetylcholinesterase and their distribution in endplate and non-endplate regions of rat diaphragm muscle. J. Neurobiol. 1973, 4, 343–361. [Google Scholar] [CrossRef]
- Bloch, R.J. Loss of acetylcholine receptor clusters induced by treatment of cultured rat myotubes with carbachol. J. Neurosci. 1986, 6, 691–700. [Google Scholar] [PubMed]
- Misgeld, T.; Kummer, T.T.; Lichtman, J.W.; Sanes, J.R. Agrin promotes synaptic differentiation by counteracting an inhibitory effect of neurotransmitter. Proc. Natl. Acad. Sci. USA 2005, 102, 11088–11093. [Google Scholar] [CrossRef] [PubMed]
- Thelen, M.H.; Simonides, W.S.; van Hardeveld, C. Electrical stimulation of C2C12 myotubes induces contractions and represses thyroid-hormone-dependent transcription of the fast-type sarcoplasmic-reticulum Ca2+-ATPase gene. Biochem. J. 1997, 321, 845–848. [Google Scholar] [CrossRef] [PubMed]
- McMahon, D.K.; Anderson, P.A.; Nassar, R.; Bunting, J.B.; Saba, Z.; Oakeley, A.E.; Malouf, N.N. C2C12 cells: Biophysical, biochemical, and immunocytochemical properties. Am. J. Physiol. 1994, 266, C1795–C1802. [Google Scholar]
- Lorenzon, P.; Giovannelli, A.; Ragozzino, D.; Eusebi, F.; Ruzzier, F. Spontaneous and repetitive calcium transients in C2C12 mouse myotubes during in vitro myogenesis. Eur. J. Neurosci. 1997, 9, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Asano, T.; Ishizuka, T.; Morishima, K.; Yawo, H. Optogenetic induction of contractile ability in immature C2C12 myotubes. Sci. Rep. 2015, 5, 8317. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Nedachi, T.; Kanzaki, M. Accelerated de novo sarcomere assembly by electric pulse stimulation in C2C12 myotubes. Exp. Cell Res. 2007, 313, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Niu, W.; Bilan, P.J.; Yu, J.; Gao, J.; Boguslavsky, S.; Schertzer, J.D.; Chu, G.; Yao, Z.; Klip, A. PKCε regulates contraction-stimulated GLUT4 traffic in skeletal muscle cells. J. Cell. Physiol 2011, 226, 173–180. [Google Scholar] [CrossRef]
- Niu, W.; Bilan, P.J.; Ishikura, S.; Schertzer, J.D.; Contreras-Ferrat, A.; Fu, Z.; Liu, J.; Boguslavsky, S.; Foley, K.P.; Liu, Z.; et al. Contraction-related stimuli regulate glut4 traffic in C2C12-GLUT4myc skeletal muscle cells. Am. J. Physiol. Endocrinol. Metab. 2010, 298, E1058–E1071. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Peng, H.B. Mechanism of acetylcholine receptor cluster formation induced by DC electric field. PLoS ONE 2011, 6, e26805. [Google Scholar] [CrossRef] [PubMed]
- Bandi, E.; Jevsek, M.; Mars, T.; Jurdana, M.; Formaggio, E.; Sciancalepore, M.; Fumagalli, G.; Grubic, Z.; Ruzzier, F.; Lorenzon, P. Neural agrin controls maturation of the excitation-contraction coupling mechanism in human myotubes developing in vitro. Am. J. Physiol. Cell Physiol. 2008, 294, C66–C73. [Google Scholar] [CrossRef]
- Arnold, A.S.; Gueye, M.; Guettier-Sigrist, S.; Courdier-Fruh, I.; Coupin, G.; Poindron, P.; Gies, J.P. Reduced expression of nicotinic achrs in myotubes from spinal muscular atrophy I patients. Lab. Investig. 2004, 84, 1271–1278. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. Principles of Neural Science, 5th ed.; McGraw-Hill: New York, NY, USA, 2013; 1709p. [Google Scholar]
- Lemon, R.N.; Griffiths, J. Comparing the function of the corticospinal system in different species: Organizational differences for motor specialization? Muscle Nerve 2005, 32, 261–279. [Google Scholar] [CrossRef]
- Maeda, H.; Fukuda, S.; Kameda, H.; Murabe, N.; Isoo, N.; Mizukami, H.; Ozawa, K.; Sakurai, M. Corticospinal axons make direct synaptic connections with spinal motoneurons innervating forearm muscles early during postnatal development in the rat. J. Physiol. 2016, 594, 189–205. [Google Scholar] [CrossRef]
- Burden, S.J. The formation of neuromuscular synapses. Genes Dev. 1998, 12, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Sanes, J.R.; Lichtman, J.W. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 1999, 22, 389–442. [Google Scholar] [CrossRef]
- Lupa, M.T.; Gordon, H.; Hall, Z.W. A specific effect of muscle cells on the distribution of presynaptic proteins in neurites and its absence in a C2 muscle cell variant. Dev. Biol. 1990, 142, 31–43. [Google Scholar] [CrossRef]
- Fischbach, G.D.; Berg, D.K.; Cohen, S.A.; Frank, E. Enrichment of nerve-muscle synapses in spinal cord-muscle cultures and identification of relative peaks of ach sensitivity at sites of transmitter release. Cold Spring Harb. Symp. Quant. Biol. 1976, 40, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Fischman, D.A.; Moscona, A.A. The development of nerve-muscle junctions in monolayer cultures of embryonic spinal cord and skeletal muscle cells. J. Cell Biol. 1969, 43, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Role, L.W.; Roufa, D.G.; Fischbach, G.D. The distribution of acetylcholine receptor clusters and sites of transmitter release along chick ciliary ganglion neurite-myotube contacts in culture. J. Cell Biol. 1987, 104, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Burgess, R.W.; Nguyen, Q.T.; Son, Y.J.; Lichtman, J.W.; Sanes, J.R. Alternatively spliced isoforms of nerve- and muscle-derived agrin: Their roles at the neuromuscular junction. Neuron 1999, 23, 33–44. [Google Scholar] [CrossRef]
- Nelson, P.; Christian, C.; Nirenberg, M. Synapse formation between clonal neuroblastoma x glioma hybrid cells and striated muscle cells. Proc. Natl. Acad. Sci. USA 1976, 73, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Kidokoro, Y.; Kravis, T.C.; Moser, K.M.; Taylor, J.C.; Crawford, I.P. Relationship of leukocyte elastase concentration to severity of emphysema in homozygous alpha1-antitrypsin-deficient persons. Am. Rev. Respir. Dis. 1977, 115, 793–803. [Google Scholar] [PubMed]
- Crain, S.M.; Alfei, L.; Peterson, E.R. Neuromuscular transmission in cultures of adult human and rodent skeletal muscle after innervation in vitro by fetal rodent spinal cord. J. Neurobiol. 1970, 1, 471–489. [Google Scholar] [CrossRef] [PubMed]
- Engel, A.; Franzini-Armstrong, C. Myology: Basic and Clinical, 2nd ed.; McGraw-Hill: New York, NY, USA, 1994. [Google Scholar]
- Eckle, V.S.; Drexler, B.; Grasshoff, C.; Seeger, T.; Thiermann, H.; Antkowiak, B. Spinal cord-skeletal muscle cocultures detect muscle-relaxant action of botulinum neurotoxin a. ALTEX 2014, 31, 433–440. [Google Scholar] [CrossRef]
- Drexler, B.; Thiermann, H.; Antkowiak, B.; Grasshoff, C. Effects of succinylcholine in an organotypic spinal cord-skeletal muscle coculture of embryonic mice. Chem. Biol. Interact. 2013, 206, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Drexler, B.; Seeger, T.; Grasshoff, C.; Thiermann, H.; Antkowiak, B. Long-term evaluation of organophosphate toxicity and antidotal therapy in co-cultures of spinal cord and muscle tissue. Toxicol. Lett. 2011, 206, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Uzel, S.G.; Platt, R.J.; Subramanian, V.; Pearl, T.M.; Rowlands, C.J.; Chan, V.; Boyer, L.A.; So, P.T.; Kamm, R.D. Microfluidic device for the formation of optically excitable, three-dimensional, compartmentalized motor units. Sci. Adv. 2016, 2, e1501429. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Heinemann, S.; Kidokoro, Y. Cholinergic metabolism and synapse formation by a rat nerve cell line. Proc. Natl. Acad. Sci. USA 1977, 74, 2579–2583. [Google Scholar] [CrossRef] [PubMed]
- Askanas, V.; Engel, W.K.; Kobayashi, T. Thyrotropin-releasing hormone enhances motor neuron-evoked contractions of cultured human muscle. Ann. Neurol. 1985, 18, 716–719. [Google Scholar] [CrossRef] [PubMed]
- Martinuzzi, A.; Askanas, V.; Kobayashi, T.; Engel, W.K.; Gorsky, J.E. Developmental expression of the muscle-specific isozyme of phosphoglycerate mutase in human muscle cultured in monolayer and innervated by fetal rat spinal cord. Exp. Neurol. 1987, 96, 365–375. [Google Scholar] [CrossRef]
- Vita, G.; Askanas, V.; Martinuzzi, A.; Engel, W.K. Histoenzymatic profile of human muscle cultured in monolayer and innervated de novo by fetal rat spinal cord. Muscle Nerve 1988, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Martinuzzi, A.; Askanas, V.; Engel, W.K. Paralysis of innervated cultured human muscle fibers affects enzymes differentially. J. Neurochem. 1990, 54, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Rezonja, K.; Lorenzon, P.; Mars, T. Opposing effects of dexamethasone, agrin and sugammadex on functional innervation and constitutive secretion of IL-6 in in vitro innervated primary human muscle cells. Neurosci. Lett. 2013, 549, 186–190. [Google Scholar] [CrossRef]
- Prpar Mihevc, S.; Pavlin, M.; Darovic, S.; Zivin, M.; Podbregar, M.; Rogelj, B.; Mars, T. Modelling fus mislocalisation in an in vitro model of innervated human muscle. J. Mol. Neurosci. 2017, 62, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.S.; Christe, M.; Handschin, C. A functional motor unit in the culture dish: Co-culture of spinal cord explants and muscle cells. J. Vis. Exp. 2012. [Google Scholar] [CrossRef] [PubMed]
- Ecob-Prince, M.S.; Jenkison, M.; Butler-Browne, G.S.; Whalen, R.G. Neonatal and adult myosin heavy chain isoforms in a nerve-muscle culture system. J. Cell Biol. 1986, 103, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Askanas, V.; Engel, W.K.; Vitadello, M.; Sartore, S. Myosin isoenzymes in cultured human muscle. Arch. Neurol. 1982, 39, 347–349. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Gonzalez, M.; Stancescu, M.; Vandenburgh, H.H.; Hickman, J.J. Neuromuscular junction formation between human stem cell-derived motoneurons and human skeletal muscle in a defined system. Biomaterials 2011, 32, 9602–9611. [Google Scholar] [CrossRef]
- Demestre, M.; Orth, M.; Fohr, K.J.; Achberger, K.; Ludolph, A.C.; Liebau, S.; Boeckers, T.M. Formation and characterisation of neuromuscular junctions between hipsc derived motoneurons and myotubes. Stem Cell Res. 2015, 15, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, G.; Laine, J.; Darabi, R.; Fournier, E.; Perlingeiro, R.; Tabti, N. Physiological and ultrastructural features of human induced pluripotent and embryonic stem cell-derived skeletal myocytes in vitro. Proc. Natl. Acad. Sci. USA 2014, 111, 8275–8280. [Google Scholar] [CrossRef] [PubMed]
- Abujarour, R.; Valamehr, B. Generation of skeletal muscle cells from pluripotent stem cells: Advances and challenges. Front. Cell Dev. Biol. 2015, 3, 29. [Google Scholar] [CrossRef]
- Michikawa, M.; Kobayashi, T.; Tsukagoshi, H. Early events of chemical transmission of newly formed neuromuscular junctions in monolayers of human muscle cells co-cultured with fetal rat spinal cord explants. Brain Res. 1991, 538, 79–85. [Google Scholar] [CrossRef]
- Saito, K.; Kobayashi, T.; Askanas, V.; Engel, W.K.; Ishikawa, K. Electrical properties of human muscle cultured in monolayer aneurally and co-cultured with fetal rat spinal cord. Biomed. Res. 1990, 11, 19–28. [Google Scholar] [CrossRef]
- Rezonja, K.; Sostaric, M.; Vidmar, G.; Mars, T. Dexamethasone produces dose-dependent inhibition of sugammadex reversal in in vitro innervated primary human muscle cells. Anesth. Analg. 2014, 118, 755–763. [Google Scholar] [CrossRef] [PubMed]
- Eckle, V.S.; Balk, M.; Thiermann, H.; Antkowiak, B.; Grasshoff, C. Botulinum toxin b increases intrinsic muscle activity in organotypic spinal cord-skeletal muscle co-cultures. Toxicol. Lett. 2016, 244, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Weimer, I.; Worek, F.; Seeger, T.; Thiermann, H.; Eckle, V.S.; Grasshoff, C.; Antkowiak, B. Self-regeneration of neuromuscular function following soman and vx poisoning in spinal cord-skeletal muscle cocultures. Toxicol. Lett. 2016, 244, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Magloire, V.; Streit, J. Intrinsic activity and positive feedback in motor circuits in organotypic spinal cord slice cultures. Eur. J. Neurosci. 2009, 30, 1487–1497. [Google Scholar] [CrossRef]
- Jevsek, M.; Mars, T.; Mis, K.; Grubic, Z. Origin of acetylcholinesterase in the neuromuscular junction formed in the in vitro innervated human muscle. Eur. J. Neurosci. 2004, 20, 2865–2871. [Google Scholar] [CrossRef] [PubMed]
- Lieth, E.; Fallon, J.R. Muscle agrin: Neural regulation and localization at nerve-induced acetylcholine receptor clusters. J. Neurosci. 1993, 13, 2509–2514. [Google Scholar] [PubMed]
- Martinuzzi, A.; Vergani, L.; Carrozzo, R.; Fanin, M.; Bartoloni, L.; Angelini, C.; Askanas, V.; Engel, W.K. Expression of muscle-type phosphorylase in innervated and aneural cultured muscle of patients with myophosphorylase deficiency. J. Clin. Investig. 1993, 92, 1774–1780. [Google Scholar] [CrossRef] [PubMed]
- Park-Matsumoto, Y.C.; Kameda, N.; Kobayashi, T.; Tsukagoshi, H. Developmental study of the expression of dystrophin in cultured human muscle aneurally and innervated with fetal rat spinal cord. Brain Res. 1991, 565, 280–289. [Google Scholar] [CrossRef]
- Kobayashi, T.; Askanas, V.; Saito, K.; Engel, W.K.; Ishikawa, K. Abnormalities of aneural and innervated cultured muscle fibers from patients with myotonic atrophy (dystrophy). Arch. Neurol. 1990, 47, 893–896. [Google Scholar] [CrossRef]
- Braun, S.; Croizat, B.; Lagrange, M.C.; Warter, J.M.; Poindron, P. Constitutive muscular abnormalities in culture in spinal muscular atrophy. Lancet 1995, 345, 694–695. [Google Scholar] [CrossRef]
- Guettier-Sigrist, S.; Hugel, B.; Coupin, G.; Freyssinet, J.M.; Poindron, P.; Warter, J.M. Possible pathogenic role of muscle cell dysfunction in motor neuron death in spinal muscular atrophy. Muscle Nerve 2002, 25, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Dorchies, O.M.; Laporte, J.; Wagner, S.; Hindelang, C.; Warter, J.M.; Mandel, J.L.; Poindron, P. Normal innervation and differentiation of X-linked myotubular myopathy muscle cells in a nerve-muscle coculture system. Neuromuscul. Disord. 2001, 11, 736–746. [Google Scholar] [CrossRef]
- Martinuzzi, A.; Askanas, V.; Kobayashi, T.; Engel, W.K.; Di Mauro, S. Expression of muscle-gene-specific isozymes of phosphorylase and creatine kinase in innervated cultured human muscle. J. Cell Biol. 1986, 103, 1423–1429. [Google Scholar] [CrossRef] [PubMed]
- Martinuzzi, A.; Askanas, V.; Kobayashi, T.; Engel, W.K. Asynchronous regulation of muscle specific isozymes of creatine kinase, glycogen phosphorylase, lactic dehydrogenase and phosphoglycerate mutase in innervated and non-innervated cultured human muscle. Neurosci. Lett. 1988, 89, 216–222. [Google Scholar] [CrossRef]
- Mars, T.; Mis, K.; Pirkmajer, S.; Katalinic, K.; Grubic, Z. The effects of organophosphates in the early stages of skeletal muscle regeneration. In Handbook of Toxicology of Chemical Warfare Agents, 2nd ed.; Gupta, R.C., Ed.; Elsevier/Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2015; pp. 751–759. [Google Scholar]
- Schiaffino, S.; Reggiani, C. Fiber types in mammalian skeletal muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed]
- Ecob-Prince, M.; Hill, M.; Brown, W. Myosin heavy chain expression in human muscle cocultured with mouse spinal cord. J. Neurol. Sci. 1989, 90, 167–177. [Google Scholar] [CrossRef]
- Kameda, N.; Kobayashi, T.; Park-Matsumoto, Y.C.; Tsukagoshi, H.; Shimizu, T. Developmental studies of the expression of myosin heavy chain isoforms in cultured human muscle aneurally and innervated with fetal rat spinal cord. J. Neurol. Sci. 1993, 114, 85–98. [Google Scholar] [CrossRef]
- Tanaka, H.; Furuya, T.; Kameda, N.; Kobayashi, T.; Mizusawa, H. Triad proteins and intracellular Ca2+ transients during development of human skeletal muscle cells in aneural and innervated cultures. J. Muscle Res. Cell Motil. 2000, 21, 507–526. [Google Scholar] [CrossRef]
- Armstrong, C.M.; Bezanilla, F.M.; Horowicz, P. Twitches in the presence of ethylene glycol bis(-aminoethyl ether)-N,N′-tetracetic acid. Biochim. Biophys. Acta 1972, 267, 605–608. [Google Scholar] [CrossRef]
- Inoue, I.; Tsutsui, I.; Bone, Q. Excitation-contraction coupling in skeletal and caudal heart muscle of the hagfish eptatretus burgeri girard. J. Exp. Biol. 2002, 205, 3535–3541. [Google Scholar] [PubMed]
- Askanas, V.; McFerrin, J.; Park-Matsumoto, Y.C.; Lee, C.S.; Engel, W.K. Glucocorticoid increases acetylcholinesterase and organization of the postsynaptic membrane in innervated cultured human muscle. Exp. Neurol. 1992, 115, 368–375. [Google Scholar] [CrossRef]
- Rubin, L.L.; Schuetze, S.M.; Weill, C.L.; Fischbach, G.D. Regulation of acetylcholinesterase appearance at neuromuscular junctions in vitro. Nature 1980, 283, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Bach, A.D.; Beier, J.P.; Stark, G.B. Expression of trisk 51, agrin and nicotinic-acetycholine receptor epsilon-subunit during muscle development in a novel three-dimensional muscle-neuronal co-culture system. Cell Tissue Res. 2003, 314, 263–274. [Google Scholar] [CrossRef]
- Hartley, R.S.; Yablonka-Reuveni, Z. Long-term maintenance of primary myogenic cultures on a reconstituted basement membrane. In Vitro Cell Dev. Biol. 1990, 26, 955–961. [Google Scholar] [CrossRef]
- Happe, C.L.; Tenerelli, K.P.; Gromova, A.K.; Kolb, F.; Engler, A.J. Mechanically patterned neuromuscular junctions-in-a-dish have improved functional maturation. Mol. Biol. Cell 2017, 28, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- Goetsch, K.P.; Myburgh, K.H.; Niesler, C.U. In vitro myoblast motility models: Investigating migration dynamics for the study of skeletal muscle repair. J. Muscle Res. Cell Motil. 2013, 34, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Goetsch, K.P.; Snyman, C.; Myburgh, K.H.; Niesler, C.U. Simultaneous isolation of enriched myoblasts and fibroblasts for migration analysis within a novel co-culture assay. Biotechniques 2015, 58, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Daniels, M.P.; Lowe, B.T.; Shah, S.; Ma, J.; Samuelsson, S.J.; Lugo, B.; Parakh, T.; Uhm, C.S. Rodent nerve-muscle cell culture system for studies of neuromuscular junction development: Refinements and applications. Microsc Res. Tech. 2000, 49, 26–37. [Google Scholar] [CrossRef]
- Smith, M.A.; O’Dowd, D.K. Cell-specific regulation of agrin RNA splicing in the chick ciliary ganglion. Neuron 1994, 12, 795–804. [Google Scholar] [CrossRef]
- Yumoto, N.; Kim, N.; Burden, S.J. Lrp4 is a retrograde signal for presynaptic differentiation at neuromuscular synapses. Nature 2012, 489, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Lenzi, J.; Pagani, F.; De Santis, R.; Limatola, C.; Bozzoni, I.; Di Angelantonio, S.; Rosa, A. Differentiation of control and ALS mutant human iPSCs into functional skeletal muscle cells, a tool for the study of neuromuscolar diseases. Stem Cell Res. 2016, 17, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Sancho, S.; Mongini, T.; Tanji, K.; Tapscott, S.J.; Walker, W.F.; Weintraub, H.; Miller, A.D.; Miranda, A.F. Analysis of dystrophin expression after activation of myogenesis in amniocytes, chorionic-villus cells, and fibroblasts. A new method for diagnosing Duchenne’s muscular dystrophy. N. Engl. J. Med. 1993, 329, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Berg, D.K.; Fischbach, G.D. Enrichment of spinal cord cell cultures with motoneurons. J. Cell Biol. 1978, 77, 83–98. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, R.J.; Fischbach, G.D. Isolation of embryonic chick motoneurons and their survival in vitro. J. Neurosci. 1986, 6, 3265–3274. [Google Scholar] [PubMed]
- Okado, N.; Oppenheim, R.W. Cell death of motoneurons in the chick embryo spinal cord. IX. The loss of motoneurons following removal of afferent inputs. J. Neurosci. 1984, 4, 1639–1652. [Google Scholar]
- Riethmacher, D.; Sonnenberg-Riethmacher, E.; Brinkmann, V.; Yamaai, T.; Lewin, G.R.; Birchmeier, C. Severe neuropathies in mice with targeted mutations in the ErbB3 receptor. Nature 1997, 389, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W.; Barres, B.A. New views on synapse-glia interactions. Curr. Opin. Neurobiol. 1996, 6, 615–621. [Google Scholar] [CrossRef]
- Mirsky, R.; Stewart, H.J.; Tabernero, A.; Bradke, F.; Brennan, A.; Dong, Z.; Jessen, K.R. Development and differentiation of Schwann cells. Rev. Neurol. 1996, 152, 308–313. [Google Scholar] [PubMed]
- Jessen, K.R.; Mirsky, R. Developmental regulation in the Schwann cell lineage. Adv. Exp. Med. Biol. 1999, 468, 3–12. [Google Scholar] [PubMed]
- Peng, H.B.; Yang, J.F.; Dai, Z.; Lee, C.W.; Hung, H.W.; Feng, Z.H.; Ko, C.P. Differential effects of neurotrophins and Schwann cell-derived signals on neuronal survival/growth and synaptogenesis. J. Neurosci. 2003, 23, 5050–5060. [Google Scholar] [PubMed]
- Feng, Z.; Ko, C.P. Schwann cells promote synaptogenesis at the neuromuscular junction via transforming growth factor-β1. J. Neurosci. 2008, 28, 9599–9609. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Francisco, G.E. New insights into the pathophysiology of post-stroke spasticity. Front. Hum. Neurosci. 2015, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Czarnecki, A.; Magloire, V.; Streit, J. Modulation of intrinsic spiking in spinal cord neurons. J. Neurophysiol. 2009, 102, 2441–2452. [Google Scholar] [CrossRef] [PubMed]
- Galante, M.; Nistri, A.; Ballerini, L. Opposite changes in synaptic activity of organotypic rat spinal cord cultures after chronic block of AMPA/kainate or glycine and GABAA receptors. J. Physiol. 2000, 523, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Ballerini, L.; Galante, M. Network bursting by organotypic spinal slice cultures in the presence of bicuculline and/or strychnine is developmentally regulated. Eur. J. Neurosci. 1998, 10, 2871–2879. [Google Scholar] [CrossRef] [PubMed]
- Tscherter, A.; Heuschkel, M.O.; Renaud, P.; Streit, J. Spatiotemporal characterization of rhythmic activity in rat spinal cord slice cultures. Eur. J. Neurosci. 2001, 14, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Rosato-Siri, M.; Grandolfo, M.; Ballerini, L. Activity-dependent modulation of GABAergic synapses in developing rat spinal networks in vitro. Eur. J. Neurosci. 2002, 16, 2123–2135. [Google Scholar] [CrossRef] [PubMed]
- Grillner, S. Biological pattern generation: The cellular and computational logic of networks in motion. Neuron 2006, 52, 751–766. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, M.J. The origin of spontaneous activity in developing networks of the vertebrate nervous system. Curr. Opin. Neurobiol. 1999, 9, 94–104. [Google Scholar] [CrossRef]
- Carbonetto, S.; Lindenbaum, M. The basement membrane at the neuromuscular junction: A synaptic mediatrix. Curr. Opin. Neurobiol. 1995, 5, 596–605. [Google Scholar] [CrossRef]
- Anglister, L.; McMahan, U.J. Basal lamina directs acetylcholinesterase accumulation at synaptic sites in regenerating muscle. J. Cell Biol. 1985, 101, 735–743. [Google Scholar] [CrossRef]
- Mis, K.; Mars, T.; Jevsek, M.; Strasek, H.; Golicnik, M.; Brecelj, J.; Komel, R.; King, M.P.; Miranda, A.F.; Grubic, Z. Expression and distribution of acetylcholinesterase among the cellular components of the neuromuscular junction formed in human myotube in vitro. Chem. Biol. Interact. 2005, 157–158, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Bernard, V.; Girard, E.; Hrabovska, A.; Camp, S.; Taylor, P.; Plaud, B.; Krejci, E. Distinct localization of collagen q and prima forms of acetylcholinesterase at the neuromuscular junction. Mol. Cell. Neurosci. 2011, 46, 272–281. [Google Scholar] [CrossRef]
- Gaspersic, R.; Koritnik, B.; Erzen, I.; Sketelj, J. Muscle activity-resistant acetylcholine receptor accumulation is induced in places of former motor endplates in ectopically innervated regenerating rat muscles. Int. J. Dev. Neurosci. 2001, 19, 339–346. [Google Scholar] [CrossRef]
- Peng, H.B.; Xie, H.; Rossi, S.G.; Rotundo, R.L. Acetylcholinesterase clustering at the neuromuscular junction involves perlecan and dystroglycan. J. Cell Biol. 1999, 145, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Arikawa-Hirasawa, E.; Rossi, S.G.; Rotundo, R.L.; Yamada, Y. Absence of acetylcholinesterase at the neuromuscular junctions of perlecan-null mice. Nat. Neurosci. 2002, 5, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, B.J.; Lee, R.K.; Rotundo, R.L. Compartmentalization of acetylcholinesterase mRNA and enzyme at the vertebrate neuromuscular junction. Neuron 1993, 11, 467–477. [Google Scholar] [CrossRef]
- Krejci, E.; Legay, C.; Thomine, S.; Sketelj, J.; Massoulie, J. Differences in expression of acetylcholinesterase and collagen q control the distribution and oligomerization of the collagen-tailed forms in fast and slow muscles. J. Neurosci. 1999, 19, 10672–10679. [Google Scholar] [PubMed]
- Perrier, A.L.; Massoulie, J.; Krejci, E. PRiMA: The membrane anchor of acetylcholinesterase in the brain. Neuron 2002, 33, 275–285. [Google Scholar] [CrossRef]
- Xie, H.Q.; Leung, K.W.; Chen, V.P.; Lau, F.T.; Liu, L.S.; Choi, R.C.; Tsim, K.W. The promoter activity of proline-rich membrane anchor (PRiMA) of globular form acetylcholinesterase in muscle: Suppressive roles of myogenesis and innervating nerve. Chem. Biol. Interact. 2008, 175, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Boudreau-Lariviere, C.; Gisiger, V.; Michel, R.N.; Hubatsch, D.A.; Jasmin, B.J. Fast and slow skeletal muscles express a common basic profile of acetylcholinesterase molecular forms. Am. J. Physiol. 1997, 272, C68–C76. [Google Scholar] [PubMed]
- Gisiger, V.; Belisle, M.; Gardiner, P.F. Acetylcholinesterase adaptation to voluntary wheel running is proportional to the volume of activity in fast, but not slow, rat hindlimb muscles. Eur. J. Neurosci. 1994, 6, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Lomo, T.; Massoulie, J.; Vigny, M. Stimulation of denervated rat soleus muscle with fast and slow activity patterns induces different expression of acetylcholinesterase molecular forms. J. Neurosci. 1985, 5, 1180–1187. [Google Scholar] [PubMed]
- Blotnick, E.; Anglister, L. Exercise modulates synaptic acetylcholinesterase at neuromuscular junctions. Neuroscience 2016, 319, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Younkin, S.G.; Rosenstein, C.; Collins, P.L.; Rosenberry, T.L. Cellular localization of the molecular forms of acetylcholinesterase in rat diaphragm. J. Biol. Chem. 1982, 257, 13630–13637. [Google Scholar] [PubMed]
- Silver, A. The Biology of Cholinesterases; North-Holland Pub. Co.: Amsterdam, The Netherlands; American Elsevier Pub. Co.: New York, NY, USA, 1974; pp. p. xiv, p. 596. [Google Scholar]
- Anglister, L. Acetylcholinesterase from the motor nerve terminal accumulates on the synaptic basal lamina of the myofiber. J. Cell Biol. 1991, 115, 755–764. [Google Scholar] [CrossRef]
- Schatz, C.R.; Veh, R.W. High-resolution localization of acetylcholinesterase at the rat neuromuscular junction. J. Histochem. Cytochem. 1987, 35, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Mis, K.; Mars, T.; Jevsek, M.; Brank, M.; Zajc-Kreft, K.; Grubic, Z. Localization of mrnas encoding acetylcholinesterase and butyrylcholinesterase in the rat spinal cord by nonradioactive in situ hybridization. J. Histochem. Cytochem. 2003, 51, 1633–1644. [Google Scholar] [CrossRef] [PubMed]
- Legay, C. Why so many forms of acetylcholinesterase? Microsc. Res. Tech. 2000, 49, 56–72. [Google Scholar] [CrossRef]
- Koelle, G.B.; Friedenwald, J.A. A histochemical method for localizing cholinesterase activity. Proc. Soc. Exp. Biol. Med. 1949, 70, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Brzin, M.; Pucihar, S. Iodide, thiocyanate and cyanide ions as capturing reagents in one-step copper-thiocholine method for cytochemical localization of cholinesterase activity. Histochemistry 1976, 48, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Di Giamberardino, L.; Couraud, J.Y. Rapid accumulation of high molecular weight acetylcholinesterase in transected sciatic nerve. Nature 1978, 271, 170–172. [Google Scholar] [CrossRef] [PubMed]
- Brimijoin, S. Axonal transport and subcellular distribution of molecular forms of acetylcholinesterase in rabbit sciatic nerve. Mol. Pharmacol. 1979, 15, 641–648. [Google Scholar] [PubMed]
- Couraud, J.Y.; Di Giamberardino, L.; Hassig, R.; Mira, J.C. Axonal transport of the molecular forms of acetylcholinesterase in developing and regenerating peripheral nerve. Exp. Neurol. 1983, 80, 94–110. [Google Scholar] [CrossRef]
- De la Porte, S.; Courbin, P.; Chapron, J.; Hantaz-Ambroise, D.; Koenig, J. Simultaneous demonstration of 3 synaptic markers: Nerve endings, localization of ache and achr clusters at neuromuscular junctions of the rat in vitro. Biol. Cell 1986, 56, 181–183. [Google Scholar] [CrossRef]
- Dobbertin, A.; Hrabovska, A.; Dembele, K.; Camp, S.; Taylor, P.; Krejci, E.; Bernard, V. Targeting of acetylcholinesterase in neurons in vivo: A dual processing function for the proline-rich membrane anchor subunit and the attachment domain on the catalytic subunit. J. Neurosci. 2009, 29, 4519–4530. [Google Scholar] [CrossRef]
- Camp, S.; De Jaco, A.; Zhang, L.; Marquez, M.; De la Torre, B.; Taylor, P. Acetylcholinesterase expression in muscle is specifically controlled by a promoter-selective enhancesome in the first intron. J. Neurosci. 2008, 28, 2459–2470. [Google Scholar] [CrossRef]
- Rotundo, R.L.; Rossi, S.G.; Anglister, L. Transplantation of quail collagen-tailed acetylcholinesterase molecules onto the frog neuromuscular synapse. J. Cell Biol. 1997, 136, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Girard, E.; Barbier, J.; Chatonnet, A.; Krejci, E.; Molgo, J. Synaptic remodeling at the skeletal neuromuscular junction of acetylcholinesterase knockout mice and its physiological relevance. Chem. Biol. Interact. 2005, 157–158, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Stribley, J.A.; Chatonnet, A.; Wilder, P.J.; Rizzino, A.; McComb, R.D.; Taylor, P.; Hinrichs, S.H.; Lockridge, O. Postnatal developmental delay and supersensitivity to organophosphate in gene-targeted mice lacking acetylcholinesterase. J. Pharmacol. Exp. Ther. 2000, 293, 896–902. [Google Scholar] [PubMed]
- Xie, W.; Wilder, P.J.; Stribley, J.; Chatonnet, A.; Rizzino, A.; Taylor, P.; Hinrichs, S.H.; Lockridge, O. Knockout of one acetylcholinesterase allele in the mouse. Chem. Biol. Interact. 1999, 119–120, 289–299. [Google Scholar] [CrossRef]
- Engel, A.G.; Sine, S.M. Current understanding of congenital myasthenic syndromes. Curr. Opin. Pharmacol. 2005, 5, 308–321. [Google Scholar] [CrossRef] [PubMed]
- Ohno, K.; Brengman, J.; Tsujino, A.; Engel, A.G. Human endplate acetylcholinesterase deficiency caused by mutations in the collagen-like tail subunit (colq) of the asymmetric enzyme. Proc. Natl. Acad. Sci. USA 1998, 95, 9654–9659. [Google Scholar] [CrossRef] [PubMed]
- Frank, E.; Fischbach, G.D. Early events in neuromuscular junction formation in vitro: Induction of acetylcholine receptor clusters in the postsynaptic membrane and morphology of newly formed synapses. J. Cell Biol. 1979, 83, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Cohen, M.W.; Zorychta, E. Effects of innervation on the distribution of acetylcholine receptors on cultured muscle cells. J. Physiol. 1977, 268, 731–756. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Cohen, M.W. Nerve-induced and spontaneous redistribution of acetylcholine receptors on cultured muscle cells. J. Physiol. 1977, 268, 757–773. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.W.; Weldon, P.R. Localization of acetylcholine receptors and synaptic ultrastructure at nerve-muscle contacts in culture: Dependence on nerve type. J. Cell Biol. 1980, 86, 388–401. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, A.E.; Daniels, M.P. Conditioned medium from cultures of embryonic neurons contains a high molecular weight factor which induces acetylcholine receptor aggregation on cultured myotubes. J. Neurosci. 1982, 2, 623–632. [Google Scholar] [PubMed]
- Nitkin, R.M.; Smith, M.A.; Magill, C.; Fallon, J.R.; Yao, Y.M.; Wallace, B.G.; McMahan, U.J. Identification of agrin, a synaptic organizing protein from torpedo electric organ. J. Cell Biol. 1987, 105, 2471–2478. [Google Scholar] [CrossRef] [PubMed]
- Reist, N.E.; Magill, C.; McMahan, U.J. Agrin-like molecules at synaptic sites in normal, denervated, and damaged skeletal muscles. J. Cell Biol. 1987, 105, 2457–2469. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Yao, Y.M.; Reist, N.E.; Magill, C.; Wallace, B.G.; McMahan, U.J. Identification of agrin in electric organ extracts and localization of agrin-like molecules in muscle and central nervous system. J. Exp. Biol. 1987, 132, 223–230. [Google Scholar] [PubMed]
- Magill-Solc, C.; McMahan, U.J. Synthesis and transport of agrin-like molecules in motor neurons. J. Exp. Biol. 1990, 153, 1–10. [Google Scholar] [PubMed]
- Magill-Solc, C.; McMahan, U.J. Motor neurons contain agrin-like molecules. J. Cell Biol. 1988, 107, 1825–1833. [Google Scholar] [CrossRef] [PubMed]
- Magill, C.; Reist, N.E.; Fallon, J.R.; Nitkin, R.M.; Wallace, B.G.; McMahan, U.J. Agrin. Prog. Brain Res. 1987, 71, 391–396. [Google Scholar] [PubMed]
- Gautam, M.; Noakes, P.G.; Moscoso, L.; Rupp, F.; Scheller, R.H.; Merlie, J.P.; Sanes, J.R. Defective neuromuscular synaptogenesis in agrin-deficient mutant mice. Cell 1996, 85, 525–535. [Google Scholar] [CrossRef]
- Magill-Solc, C.; McMahan, U.J. Agrin-like molecules in motor neurons. J. Physiol. 1990, 84, 78–81. [Google Scholar] [CrossRef]
- McMahan, U.J.; Horton, S.E.; Werle, M.J.; Honig, L.S.; Kroger, S.; Ruegg, M.A.; Escher, G. Agrin isoforms and their role in synaptogenesis. Curr. Opin. Cell Biol. 1992, 4, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Ruegg, M.A.; Tsim, K.W.; Horton, S.E.; Kroger, S.; Escher, G.; Gensch, E.M.; McMahan, U.J. The agrin gene codes for a family of basal lamina proteins that differ in function and distribution. Neuron 1992, 8, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Gesemann, M.; Denzer, A.J.; Ruegg, M.A. Acetylcholine receptor-aggregating activity of agrin isoforms and mapping of the active site. J. Cell Biol. 1995, 128, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Ferns, M.; Hoch, W.; Campanelli, J.T.; Rupp, F.; Hall, Z.W.; Scheller, R.H. RNA splicing regulates agrin-mediated acetylcholine receptor clustering activity on cultured myotubes. Neuron 1992, 8, 1079–1086. [Google Scholar] [CrossRef]
- Ferns, M.J.; Campanelli, J.T.; Hoch, W.; Scheller, R.H.; Hall, Z. The ability of agrin to cluster AChRs depends on alternative splicing and on cell surface proteoglycans. Neuron 1993, 11, 491–502. [Google Scholar] [CrossRef]
- Reist, N.E.; Werle, M.J.; McMahan, U.J. Agrin released by motor neurons induces the aggregation of acetylcholine receptors at neuromuscular junctions. Neuron 1992, 8, 865–868. [Google Scholar] [CrossRef]
- Cohen, M.W.; Godfrey, E.W. Early appearance of and neuronal contribution to agrin-like molecules at embryonic frog nerve-muscle synapses formed in culture. J. Neurosci. 1992, 12, 2982–2992. [Google Scholar] [PubMed]
- Cohen, M.W.; Moody-Corbett, F.; Godfrey, E.W. Neuritic deposition of agrin on culture substrate: Implications for nerve-muscle synaptogenesis. J. Neurosci. 1994, 14, 3293–3303. [Google Scholar] [PubMed]
- Anderson, M.J.; Shi, Z.Q.; Grawel, R.; Zackson, S.L. Erratic deposition of agrin during the formation of xenopus neuromuscular junctions in culture. Dev. Biol. 1995, 170, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Campagna, J.A.; Ruegg, M.A.; Bixby, J.L. Evidence that agrin directly influences presynaptic differentiation at neuromuscular junctions in vitro. Eur. J. Neurosci. 1997, 9, 2269–2283. [Google Scholar] [CrossRef] [PubMed]
- Gesemann, M.; Cavalli, V.; Denzer, A.J.; Brancaccio, A.; Schumacher, B.; Ruegg, M.A. Alternative splicing of agrin alters its binding to heparin, dystroglycan, and the putative agrin receptor. Neuron 1996, 16, 755–767. [Google Scholar] [CrossRef]
- Gee, S.H.; Montanaro, F.; Lindenbaum, M.H.; Carbonetto, S. Dystroglycan-alpha, a dystrophin-associated glycoprotein, is a functional agrin receptor. Cell 1994, 77, 675–686. [Google Scholar] [CrossRef]
- Sugiyama, J.; Bowen, D.C.; Hall, Z.W. Dystroglycan binds nerve and muscle agrin. Neuron 1994, 13, 103–115. [Google Scholar] [CrossRef]
- Campanelli, J.T.; Roberds, S.L.; Campbell, K.P.; Scheller, R.H. A role for dystrophin-associated glycoproteins and utrophin in agrin-induced AChRclustering. Cell 1994, 77, 663–674. [Google Scholar] [CrossRef]
- Bowe, M.A.; Deyst, K.A.; Leszyk, J.D.; Fallon, J.R. Identification and purification of an agrin receptor from torpedo postsynaptic membranes: A heteromeric complex related to the dystroglycans. Neuron 1994, 12, 1173–1180. [Google Scholar] [CrossRef]
- Scotton, P.; Bleckmann, D.; Stebler, M.; Sciandra, F.; Brancaccio, A.; Meier, T.; Stetefeld, J.; Ruegg, M.A. Activation of muscle-specific receptor tyrosine kinase and binding to dystroglycan are regulated by alternative mRNA splicing of agrin. J. Biol. Chem. 2006, 281, 36835–36845. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.W.; Jacobson, C.; Godfrey, E.W.; Campbell, K.P.; Carbonetto, S. Distribution of alpha-dystroglycan during embryonic nerve-muscle synaptogenesis. J. Cell Biol. 1995, 129, 1093–1101. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Burgess, R.W.; Dominguez, B.; Pfaff, S.L.; Sanes, J.R.; Lee, K.F. Distinct roles of nerve and muscle in postsynaptic differentiation of the neuromuscular synapse. Nature 2001, 410, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Arber, S.; William, C.; Li, L.; Tanabe, Y.; Jessell, T.M.; Birchmeier, C.; Burden, S.J. Patterning of muscle acetylcholine receptor gene expression in the absence of motor innervation. Neuron 2001, 30, 399–410. [Google Scholar] [CrossRef]
- Kim, N.; Burden, S.J. MuSK controls where motor axons grow and form synapses. Nat. Neurosci. 2008, 11, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Brady, S.T.; Siegel, G.J.; Albers, R.W.; Price, D.L.; Benjamins, J. Basic Neurochemistry: Principles of Molecular, Cellular, and Medical Neurobiology, 8th ed.; Elsevier/Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2012; p. xxiv, p. 1096. [Google Scholar]
- Grisaru, D.; Sternfeld, M.; Eldor, A.; Glick, D.; Soreq, H. Structural roles of acetylcholinesterase variants in biology and pathology. Eur. J. Biochem. 1999, 264, 672–686. [Google Scholar] [CrossRef] [PubMed]
- Pegan, K.; Matkovic, U.; Mars, T.; Mis, K.; Pirkmajer, S.; Brecelj, J.; Grubic, Z. Acetylcholinesterase is involved in apoptosis in the precursors of human muscle regeneration. Chem. Biol. Interact. 2010, 187, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Jurdana, M.; Fumagalli, G.; Grubic, Z.; Lorenzon, P.; Mars, T.; Sciancalepore, M. Neural agrin changes the electrical properties of developing human skeletal muscle cells. Cell. Mol. Neurobiol. 2009, 29, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.B. Apoptosis in the pathogenesis and treatment of disease. Science 1995, 267, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Jejurikar, S.S.; Kuzon, W.M., Jr. Satellite cell depletion in degenerative skeletal muscle. Apoptosis 2003, 8, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Sciorati, C.; Rigamonti, E.; Manfredi, A.A.; Rovere-Querini, P. Cell death, clearance and immunity in the skeletal muscle. Cell Death Differ. 2016, 23, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Soreq, H.; Patinkin, D.; Lev-Lehman, E.; Grifman, M.; Ginzberg, D.; Eckstein, F.; Zakut, H. Antisense oligonucleotide inhibition of acetylcholinesterase gene expression induces progenitor cell expansion and suppresses hematopoietic apoptosis ex vivo. Proc. Natl. Acad. Sci. USA 1994, 91, 7907–7911. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Yang, L.; Zhao, Q.; Caen, J.P.; He, H.Y.; Jin, Q.H.; Guo, L.H.; Alemany, M.; Zhang, L.Y.; Shi, Y.F. Induction of acetylcholinesterase expression during apoptosis in various cell types. Cell Death Differ. 2002, 9, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Toiber, D.; Berson, A.; Greenberg, D.; Melamed-Book, N.; Diamant, S.; Soreq, H. N-Acetylcholinesterase-induced apoptosis in Alzheimer’s disease. PLoS ONE 2008, 3, e3108. [Google Scholar] [CrossRef] [PubMed]
- Wessler, I.; Kirkpatrick, C.J. Acetylcholine beyond neurons: The non-neuronal cholinergic system in humans. Br. J. Pharmacol. 2008, 154, 1558–1571. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lee, B.; Johnson, G.; Naleway, J.; Guzikowski, A.; Dai, W.; Darzynkiewicz, Z. Novel assay utilizing fluorochrome-tagged physostigmine (ph-f) to in situ detect active acetylcholinesterase (AChE) induced during apoptosis. Cell Cycle 2005, 4, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Day, T.; Greenfield, S.A. Bioactivity of a peptide derived from acetylcholinesterase in hippocampal organotypic cultures. Exp. Brain Res. 2004, 155, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Du, A.; Xie, J.; Guo, K.; Yang, L.; Wan, Y.; OuYang, Q.; Zhang, X.; Niu, X.; Lu, L.; Wu, J.; et al. A novel role for synaptic acetylcholinesterase as an apoptotic deoxyribonuclease. Cell Discov. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.; Sklan, E.H.; Birikh, K.; Shapira, M.; Trejo, L.; Eldor, A.; Soreq, H. Complex regulation of acetylcholinesterase gene expression in human brain tumors. Oncogene 2002, 21, 8428–8441. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Jeong, S.H.; Yee, S.B.; Kim, T.H.; Soung, Y.H.; Ha, N.C.; Kim, N.D.; Park, J.Y.; Bae, H.R.; Park, B.S.; et al. Interactions of acetylcholinesterase with caveolin-1 and subsequently with cytochrome c are required for apoptosome formation. Carcinogenesis 2008, 29, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Kim, N.D.; Yoo, Y.H. Acetylcholinesterase plays a pivotal role in apoptosome formation. Cancer Res. 2004, 64, 2652–2655. [Google Scholar] [CrossRef] [PubMed]
- Boukouris, A.E.; Zervopoulos, S.D.; Michelakis, E.D. Metabolic enzymes moonlighting in the nucleus: Metabolic regulation of gene transcription. Trends Biochem. Sci. 2016, 41, 712–730. [Google Scholar] [CrossRef]
- Huberts, D.H.; van der Klei, I.J. Moonlighting proteins: An intriguing mode of multitasking. Biochim. Biophys. Acta 2010, 1803, 520–525. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, W.; Mulders, J.W.; Bibby, M.A.; Slingsby, C.; Bloemendal, H.; de Jong, W.W. Duck lens epsilon-crystallin and lactate dehydrogenase b4 are identical: A single–copy gene product with two distinct functions. Proc. Natl. Acad. Sci. USA 1988, 85, 7114–7118. [Google Scholar] [CrossRef] [PubMed]
- Philpott, C.C.; Klausner, R.D.; Rouault, T.A. The bifunctional iron-responsive element binding protein/cytosolic aconitase: The role of active-site residues in ligand binding and regulation. Proc. Natl. Acad. Sci. USA 1994, 91, 7321–7325. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Lakshmanan, M.; Swa, H.L.; Chen, J.; Zhang, X.; Ong, Y.S.; Loo, L.S.; Akincilar, S.C.; Gunaratne, J.; Tergaonkar, V.; et al. An oncogenic role of agrin in regulating focal adhesion integrity in hepatocellular carcinoma. Nat. Commun. 2015, 6, 6184. [Google Scholar] [CrossRef] [PubMed]
- Bassat, E.; Mutlak, Y.E.; Genzelinakh, A.; Shadrin, I.Y.; Baruch-Umansky, K.; Yifa, O.; Kain, D.; Rajchman, D.; Leach, J.; Bassat, D.R.; et al. The extracellular matrix protein agrin promotes heart regeneration in mice. Nature 2017, 547, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Burgess, R.W.; Dickman, D.K.; Nunez, L.; Glass, D.J.; Sanes, J.R. Mapping sites responsible for interactions of agrin with neurons. J. Neurochem. 2002, 83, 271–284. [Google Scholar] [CrossRef] [PubMed]
- Pirkmajer, S.; Chibalin, A.V. Na,K-ATPase regulation in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E1–E31. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T. Na+-K+ pump regulation and skeletal muscle contractility. Physiol. Rev. 2003, 83, 1269–1324. [Google Scholar] [CrossRef] [PubMed]
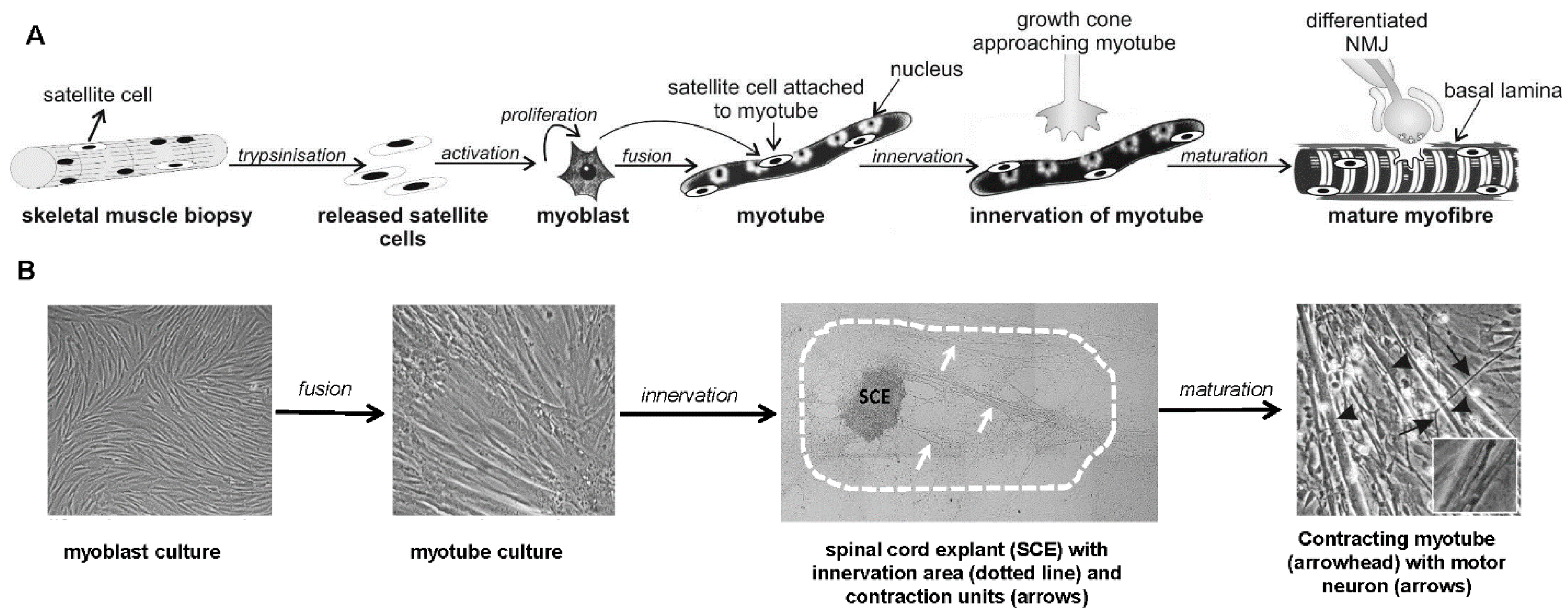

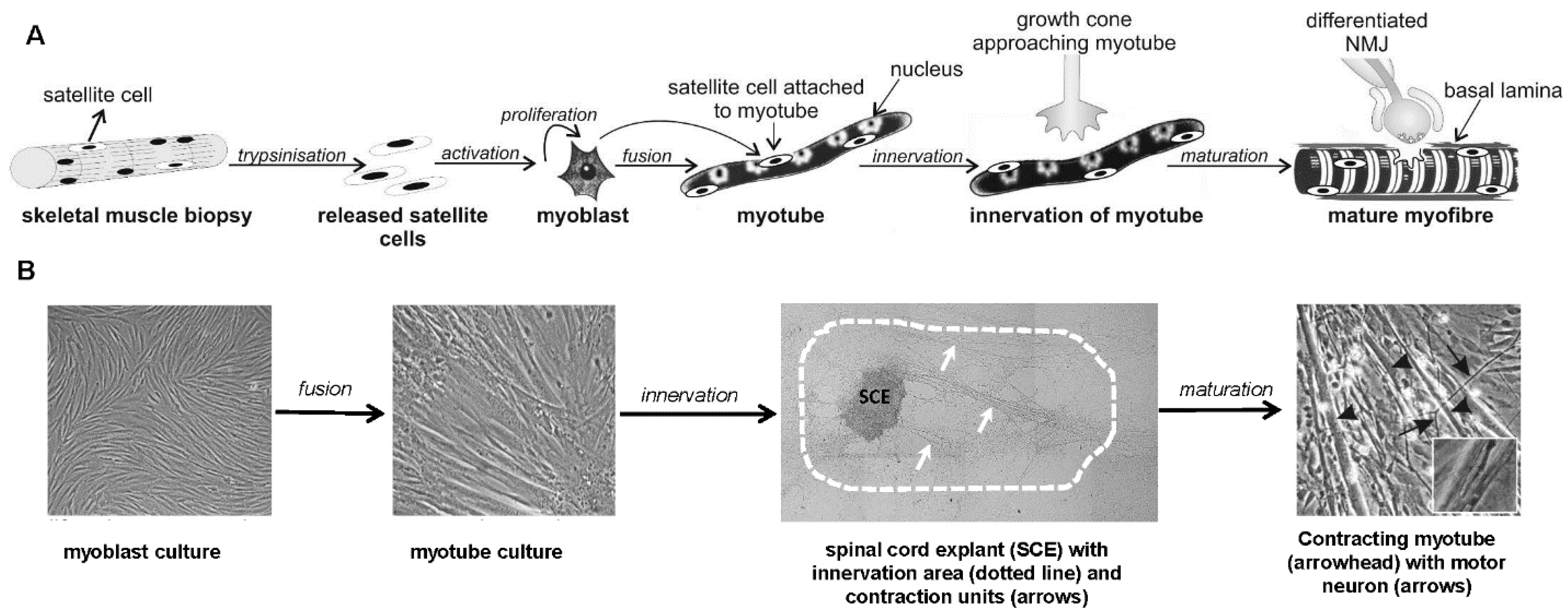{kind=link}
| Co-Culture Stage | Basal Lamina | Formation of Functional Neuromuscular Junctions | Acetylcholinesterase and nAChR |
|---|---|---|---|
| Stage I (Day 1–9) | Not formed | First neurite-myotube contact (Day 3) | Diffuse AChE staining in myotubes and neurites extending from the spinal cord explant |
| First α-bungarotoxin-sensitive contractions occur (Day 7) | AChE expressed in all myonuclei and present along the whole myotube length | ||
| No visible cross-striations in myotubes | Immature nAChR clusters at neuromuscular contacts | ||
| Stage II (Day 10–21) | Formed | Number of contraction-positive explants attains plateau (Day 10) | AChE expressed predominantely at junctional myonuclei |
| Number of contracting units attains plateau (Day 17) | Discrete patches of AChE originating in motor neurons and myotubes | ||
| Cross-striations visible in contracting myotubes | Mature nAChR cluster at neuromuscular contacts AChE and nAChR co-localize in discrete patches |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mis, K.; Grubic, Z.; Lorenzon, P.; Sciancalepore, M.; Mars, T.; Pirkmajer, S. In Vitro Innervation as an Experimental Model to Study the Expression and Functions of Acetylcholinesterase and Agrin in Human Skeletal Muscle. Molecules 2017, 22, 1418. https://doi.org/10.3390/molecules22091418
Mis K, Grubic Z, Lorenzon P, Sciancalepore M, Mars T, Pirkmajer S. In Vitro Innervation as an Experimental Model to Study the Expression and Functions of Acetylcholinesterase and Agrin in Human Skeletal Muscle. Molecules. 2017; 22(9):1418. https://doi.org/10.3390/molecules22091418
Chicago/Turabian StyleMis, Katarina, Zoran Grubic, Paola Lorenzon, Marina Sciancalepore, Tomaz Mars, and Sergej Pirkmajer. 2017. "In Vitro Innervation as an Experimental Model to Study the Expression and Functions of Acetylcholinesterase and Agrin in Human Skeletal Muscle" Molecules 22, no. 9: 1418. https://doi.org/10.3390/molecules22091418