Enhancing the Therapeutic Delivery of Oligonucleotides by Chemical Modification and Nanoparticle Encapsulation

 , and
, and
Abstract
:1. Introduction
2. Mechanism of Action and Major Application of ON Drugs
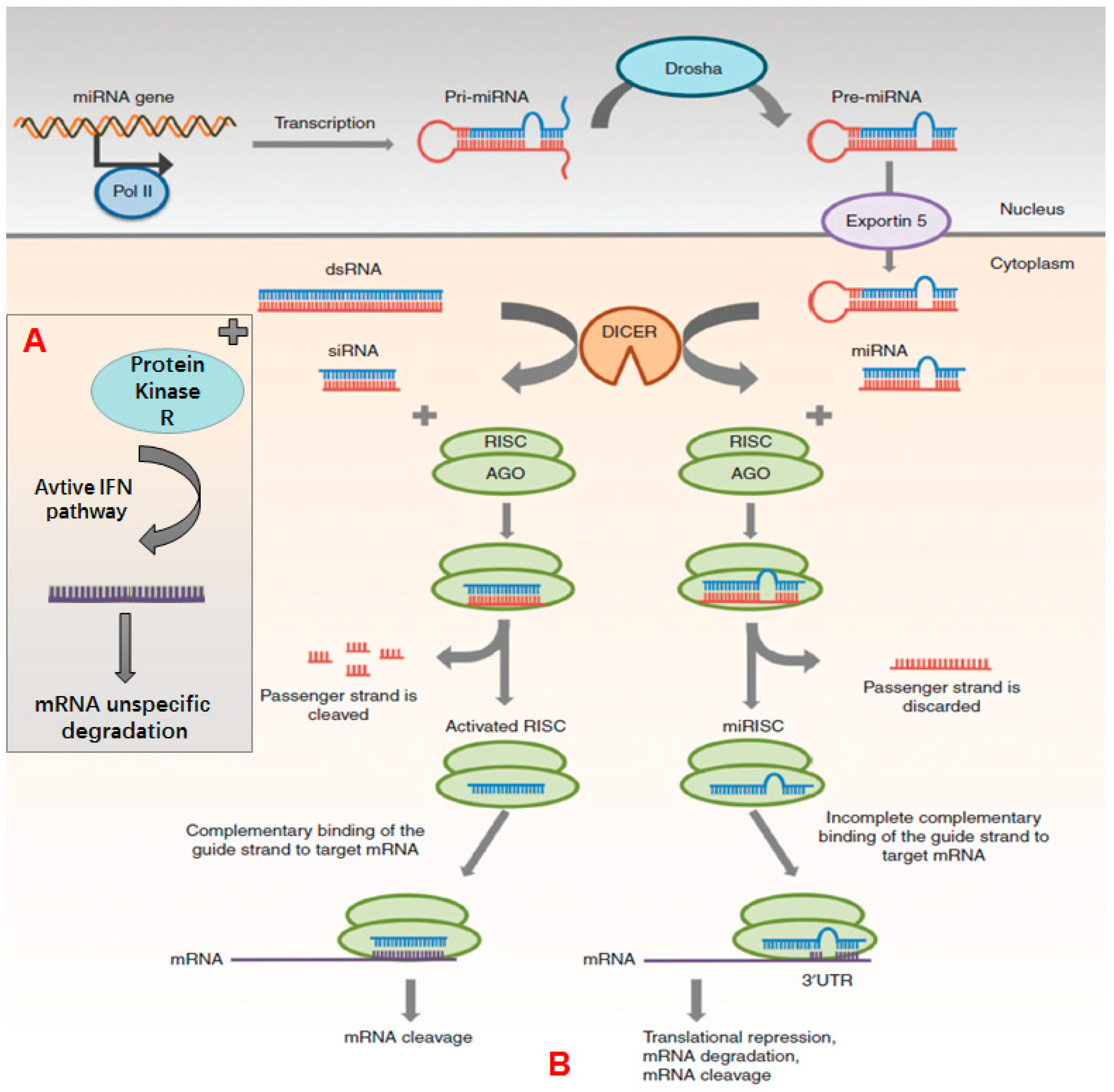
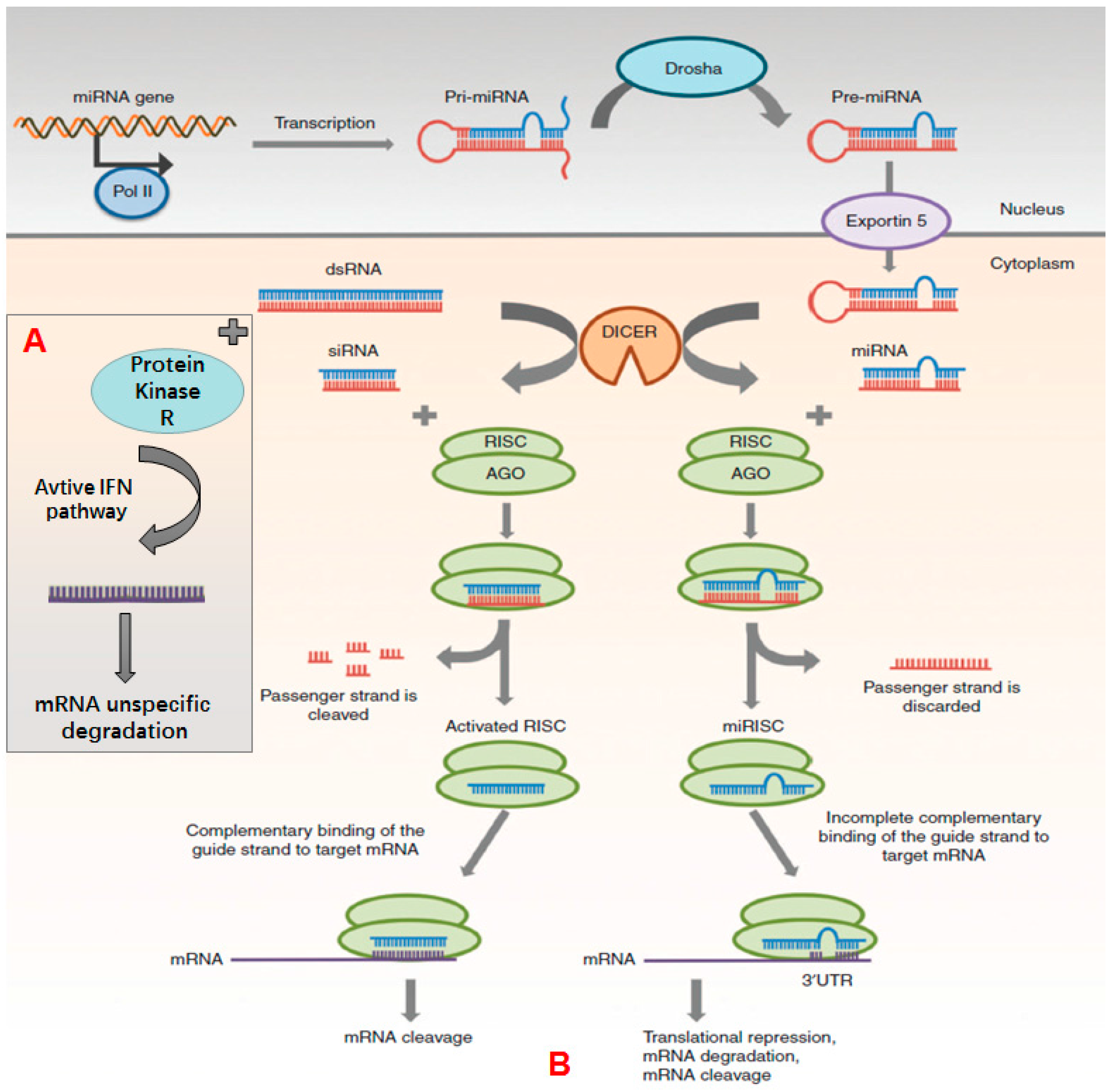
2.1. siRNA
2.2. miRNA
2.3. Anitsense ON and CpG ON
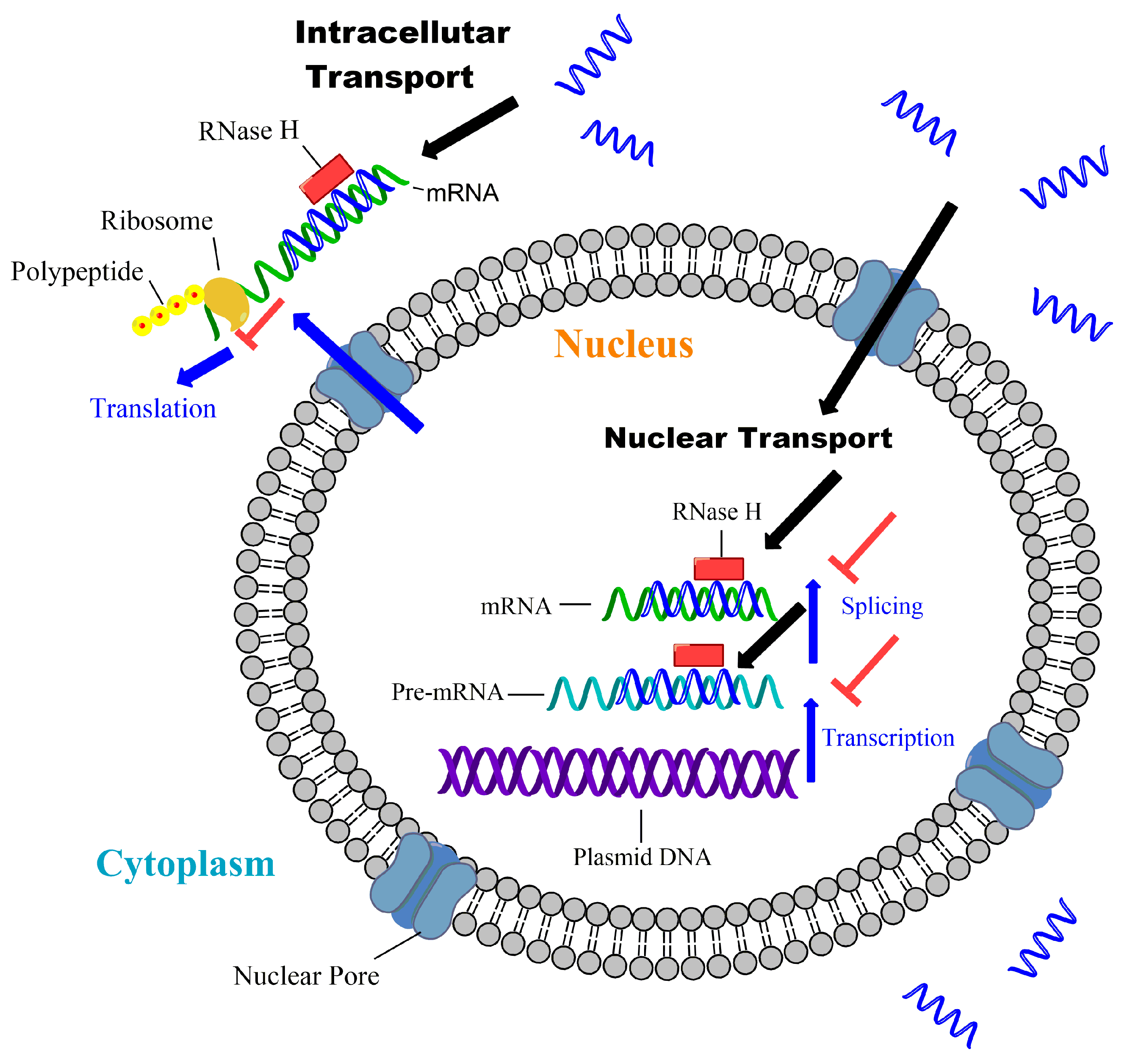
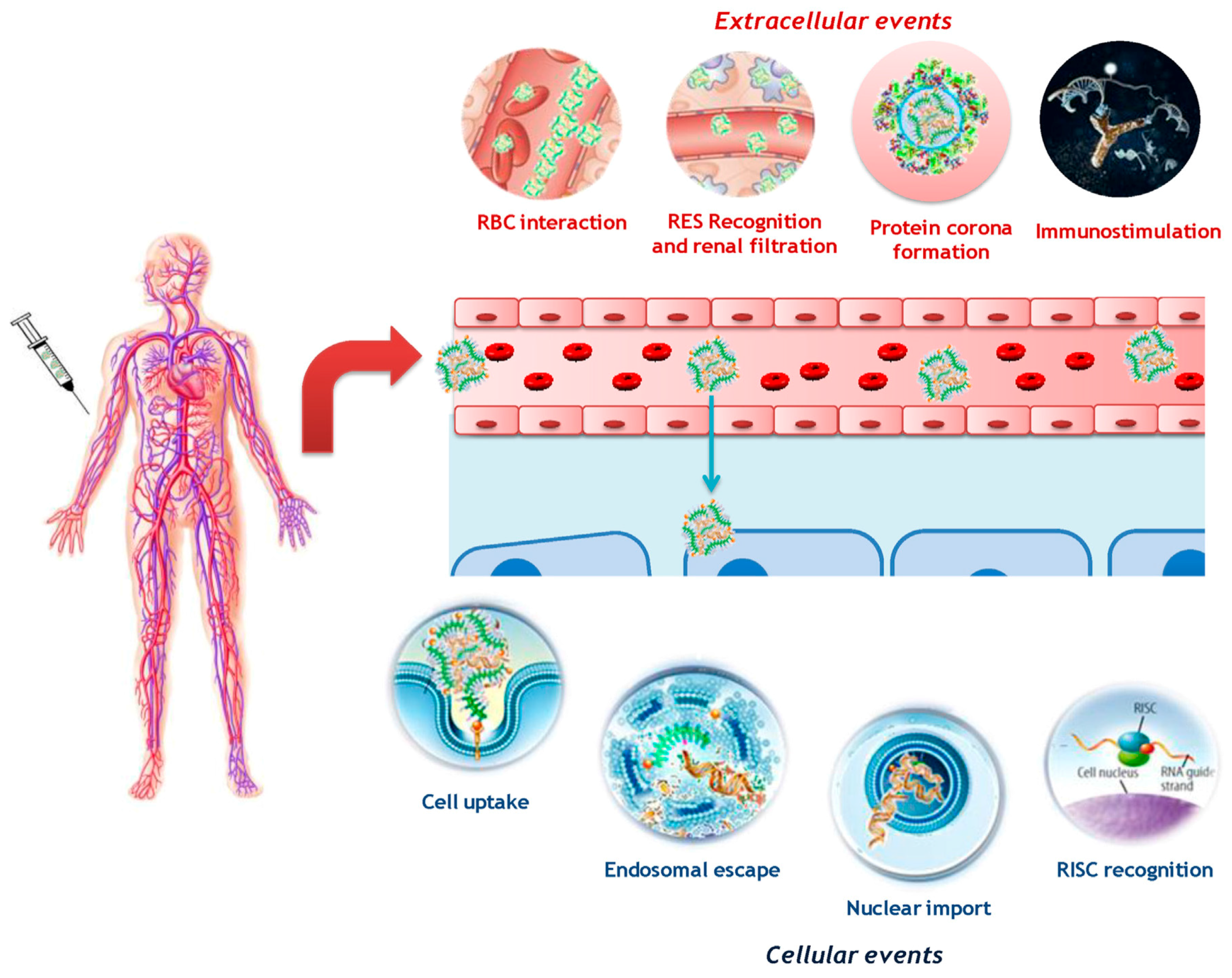
3. Barriers and Limitations for Systemic ON Drugs Delivery
4. Common Chemical Modification Strategies for ON Drugs
4.1. Structural Variants of ONs
4.1.1. The Modification of the Diester on the ON Skeleton
4.1.2. Modification of Ribose
4.1.3. Base Modification
4.1.4. ON Analogues Replace the ON Skeleton
4.2. CCPs and Ligands Conjugates
5. Drug Delivery Systems for ONs
5.1. Liposomes
5.1.1. Cationic Liposomes
5.1.2. Neutral Liposomes
5.1.3. Ionizable Liposomes
5.2. Micelles
5.3. Nanoparticles
5.3.1. Albumin-Based Nanoparticles
5.3.2. Metallic Nanoparticles
5.3.3. Other Common Nanoparticles
5.4. Targeting of Nanoparticle-Based Delivery Systems
6. Conclusions
Conflicts of Interest
Abbreviations
| 2′-O-Me | 2′-O-methylation |
| 2′-O-A | 2′-O-allylation |
| 2′-F | 2′-fluorization |
| DODMA | 1,2-dioleyloxy-N,N-dimethyl-3-aminopropane |
| DOTAP | 1,2-dioleyl-3-trimethylammonium propane |
| DOGS | dioctadecylamidoglycylspermine |
| DC-Chol | 3β-(N-(N′,N′-dimethylaminoethane)-carbamoyl) cholesterol |
| PC | phosphatidylcholine |
| PE | phosphatidylethanolamine |
| Chol | cholesterol |
| DOPE | 1,2-dioleoyl-sn-glycero-3-phosphoethanolamine |
| DODMA | 1,2-dioleyloxy-N,N-dimethyl-3-aminopropane |
| DOTAP | 1,2-dioleyl-3-trimethylammonium propane |
| DOGS | dioctadecylamidoglycylspermine |
References
- Laursen, M.B.; Pakula, M.M.; Gao, S.; Fluiter, K.; Mook, O.R.; Baas, F.; Langklaer, N.; Wengel, S.L.; Wengel, J.; Kjems, J.; et al. Utilization of unlocked nucleic acid (UNA) to enhance siRNA performance in vitro and in vivo. Mol. BioSyst. 2010, 6, 862–870. [Google Scholar] [CrossRef] [PubMed]
- Khorkova, O.; Wahlestedt, C. Oligonucleotide therapies for disorders of the nervous system. Nat. Biotechnol. 2017, 35, 249–263. [Google Scholar] [CrossRef] [PubMed]
- Rayburn, E.R.; Zhang, R. Antisense, RNAi, and gene silencing strategies for therapy: Mission possible or impossible? Drug Discov. Today 2008, 13, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Ng, E.W.; Shima, D.T.; Calias, P.; Cunningham, E.T., Jr.; Guyer, D.R.; Adamis, A.P. Pegaptanib, a targeted anti-VEGF aptamer for ocular vascular disease. Nat. Rev. Drug Discov. 2006, 5, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Geary, R.S.; Baker, B.F.; Crooke, S.T. Clinical and preclinical pharmacokinetics and pharmacodynamics of mipomersen (kynamro((R))): A second-generation antisense oligonucleotide inhibitor of apolipoprotein B. Clin. Pharmacokinet. 2015, 54, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, C. First approval in sight for Novartis’ CAR-T therapy after panel vote. Nat. Biotechnol. 2017, 35, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Miao, L.; Satterlee, A.; Huang, L. Delivery of oligonucleotides with lipid nanoparticles. Adv. Drug Deliv. Rev. 2015, 87, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.K.; Deleavey, G.F.; Damha, M.J. Chemically modified siRNA: Tools and applications. Drug Discov Today 2008, 13, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Choung, S.; Kim, Y.J.; Kim, S.; Park, H.-O.; Choi, Y.-C. Chemical modification of siRNAs to improve serum stability without loss of efficacy. Biochem. Biophys. Res. Commun. 2006, 342, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Yang, Z.; Teng, L. Nanomedicine based on nucleic acids: Pharmacokinetic and pharmacodynamic perspectives. Curr. Pharm. Biotechnol. 2014, 15, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, Y.; Yu, B.; Phelps, M.A.; Lee, L.J.; Lee, R.J. Comparative cellular pharmacokinetics and pharmacodynamics of siRNA delivery by SPANosomes and by cationic liposomes. Nanomedicine 2013, 9, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.S.; Dakwar, G.R.; Zagato, E.; Brans, T.; Figueiredo, C.; Raemdonck, K.; Azevedo, N.F.; De Smedt, S.C.; Braeckmans, K. Intracellular delivery of oligonucleotides in Helicobacter pylori by fusogenic liposomes in the presence of gastric mucus. Biomaterials 2017, 138, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Arranz, A.; Reinsch, C.; Papadakis, K.A.; Dieckmann, A.; Rauchhaus, U.; Androulidaki, A.; Zacharioudaki, V.; Margioris, A.N.; Tsatsanis, C.; Panzner, S. Treatment of experimental murine colitis with CD40 antisense oligonucleotides delivered in amphoteric liposomes. J. Control. Release Off. J. Control. Release Soc. 2013, 165, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Cavallaro, G.; Sardo, C.; Craparo, E.F.; Porsio, B.; Giammona, G. Polymeric nanoparticles for siRNA delivery: Production and applications. Int. J. Pharm. 2017, 525, 313–333. [Google Scholar] [CrossRef] [PubMed]
- Sadreddini, S.; Safaralizadeh, R.; Baradaran, B.; Aghebati-Maleki, L.; Hosseinpour-Feizi, M.A.; Shanehbandi, D.; Jadidi-Niaragh, F.; Sadreddini, S.; Kafil, H.S.; Younesi, V.; et al. Chitosan nanoparticles as a dual drug/siRNA delivery system for treatment of colorectal cancer. Immunol. Lett. 2017, 181, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Tang, L.; Xie, Y.; Xianyu, Y.; Zhang, L.; Wang, P.; Hamada, Y.; Jiang, K.; Zheng, W.; Jiang, X. Gold nanoclusters-assisted delivery of NGF siRNA for effective treatment of pancreatic cancer. Nat. Commun. 2017, 8, 15130. [Google Scholar] [CrossRef] [PubMed]
- Andrea, K.A.; Wang, L.; Carrier, A.J.; Campbell, M.; Buhariwalla, M.; Mutch, M.; MacQuarrie, S.L.; Bennett, C.; Mkandawire, M.; Oakes, K.; et al. Adsorption of Oligo-DNA on Magnesium Aluminum-Layered Double-Hydroxide Nanoparticle Surfaces: Mechanistic Implication in Gene Delivery. Langmuir ACS J. Surf. Colloids 2017, 33, 3926–3933. [Google Scholar] [CrossRef] [PubMed]
- Ragelle, H.; Colombo, S.; Pourcelle, V.; Vanvarenberg, K.; Vandermeulen, G.; Bouzin, C.; Marchand-Brynaert, J.; Feron, O.; Foged, C.; Preat, V. Intracellular siRNA delivery dynamics of integrin-targeted, PEGylated chitosan-poly(ethylene imine) hybrid nanoparticles: A mechanistic insight. J. Control. Release Off. J. Control. Release Soc. 2015, 211, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Freire, J.M.; Rego de Figueiredo, I.; Valle, J.; Veiga, A.S.; Andreu, D.; Enguita, F.J.; Castanho, M.A. siRNA-cell-penetrating peptides complexes as a combinatorial therapy against chronic myeloid leukemia using BV173 cell line as model. J. Control. Release Off. J. Control. Release Soc. 2017, 245, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Guo, X.Y.; Yang, T.; Yu, M.Z.; Chen, D.W.; Wang, J.C. Brain tumor-targeted therapy by systemic delivery of siRNA with Transferrin receptor-mediated core-shell nanoparticles. Int. J. Pharm. 2016, 510, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Aldrian, G.; Vaissiere, A.; Konate, K.; Seisel, Q.; Vives, E.; Fernandez, F.; Viguier, V.; Genevois, C.; Couillaud, F.; Demene, H.; et al. PEGylation rate influences peptide-based nanoparticles mediated siRNA delivery in vitro and in vivo. J. Control. Release Off. J. Control. Release Soc. 2017, 256, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Andrew, F.; SiQun, X.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in caenorhabditics elegans. Lett. Nat. 1998, 391, 806–811. [Google Scholar]
- Antimisiaris, S.; Mourtas, S.; Papadia, K. Targeted si-RNA with liposomes and exosomes (extracellular vesicles): How to unlock the potential. Int. J. Pharm. 2017, 525, 293–312. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.K.; Chow, M.Y.; Zhang, Y.; Leung, S.W. siRNA Versus miRNA as Therapeutics for Gene Silencing. Mol. Ther. Nucleic Acids 2015, 4, e252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritz, E. Phosphorothioates, Essential Components of Therapeutic Oligonucleotides. Nucleic Acid Ther. 2014, 24, 374–387. [Google Scholar]
- Han, L.; Tang, C.; Yin, C. Oral delivery of shRNA and siRNA via multifunctional polymeric nanoparticles for synergistic cancer therapy. Biomaterials 2014, 35, 4589–4600. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, J.E.; Barnes, C.; Khan, O.F.; Thiriot, A.; Jhunjunwala, S.; Shaw, T.E.; Xing, Y.; Sager, H.B.; Sahay, G.; Speciner, L. In vivo endothelial siRNA delivery using polymeric nanoparticles with low molecular weight. Nat. Nanotechnol. 2014, 9, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Aji Alex, M.R.; Nehate, C.; Veeranarayanan, S.; Kumar, D.S.; Kulshreshtha, R.; Koul, V. Self assembled dual responsive micelles stabilized with protein for co-delivery of drug and siRNA in cancer therapy. Biomaterials 2017, 133, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Yu-Wai-Man, C.; Tagalakis, A.D.; Manunta, M.D.; Hart, S.L.; Khaw, P.T. Receptor-targeted liposome-peptide-siRNA nanoparticles represent an efficient delivery system for MRTF silencing in conjunctival fibrosis. Sci. Rep. 2016, 6, 21881. [Google Scholar] [CrossRef] [PubMed]
- Rassu, G.; Soddu, E.; Posadino, A.M.; Pintus, G.; Sarmento, B.; Giunchedi, P.; Gavini, E. Nose-to-brain delivery of BACE1 siRNA loaded in solid lipid nanoparticles for Alzheimer’s therapy. Colloids Surf. B Biointerfaces 2017, 152, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.C.; Feinbaum, R.L.; Ambrost, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yi, H.; He, H.; Pan, H.; Cai, L.; Ma, Y. Tumor associated macrophage-targeted microRNA delivery with dual-responsive polypeptide nanovectors for anti-cancer therapy. Biomaterials 2017, 134, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-S.; Wang, Y.-H.; Zhu, X.-T.; Wu, C.-J. Genome-Wide Profiling of miRNA and mRNA Expression in Alzheimer’s Disease. Med. Sci. Monit. 2017, 23, 2721–2731. [Google Scholar] [CrossRef] [PubMed]
- Schnittert, J.; Kuninty, P.R.; Bystry, T.F.; Brock, R.; Storm, G.; Prakash, J. Anti-microRNA targeting using peptide-based nanocomplexes to inhibit differentiation of human pancreatic stellate cells. Nanomedicine 2017, 12, 1369–1384. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Deuse, T.; Stubbendorff, M.; Chernogubova, E.; Erben, R.G.; Eken, S.M.; Jin, H.; Li, Y.; Busch, A.; Heeger, C.H.; et al. Local MicroRNA Modulation Using a Novel Anti-miR-21-Eluting Stent Effectively Prevents Experimental In-Stent Restenosis Significance. Arterioscler. Thromb. Vasc. Biol. 2015, 35. [Google Scholar]
- Liu, W.; Zhang, B.; Chen, G.; Wu, W.; Zhou, L.; Shi, Y.; Zeng, Q.; Li, Y.; Sun, Y.; Deng, X.; et al. Targeting miR-21 with Sophocarpine Inhibits Tumor Progression and Reverses Epithelial-Mesenchymal Transition in Head and Neck Cancer. Mol. Ther. J. Am. Soc. Gene Ther. 2017, 25, 2129–2139. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, K.; Iseda, K.; Kawano, D.; Kawakami, J. Quantitative relationship between chemical properties and bioactivities of anti-microRNA oligonucleotides targeted to tumor-associated microRNA-21. Biochimie 2017, 137, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Li, W.M.; Chan, C.M.; Miller, A.L.; Lee, C.H. Dual Functional Roles of Molecular Beacon as a MicroRNA Detector and Inhibitor. J. Biol. Chem. 2017, 292, 3568–3580. [Google Scholar] [CrossRef] [PubMed]
- Azzouzi, S.; Mak, W.C.; Kor, K.; Turner, A.P.; Ali, M.B.; Beni, V. An integrated dual functional recognition/amplification bio-label for the one-step impedimetric detection of Micro-RNA-21. Biosens. Bioelectron. 2017, 92, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Zhao, H.; Tao, Z.; Wang, R.; Liu, P.; Han, Z.; Ma, S.; Luo, Y.; Jia, J. MicroRNA-181c Exacerbates Brain Injury in Acute Ischemic Stroke. Aging Dis. 2016, 7, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Agrawal, Y.K. Targeting nanocarriers containing antisense oligonucleotides to cancer cell. J. Drug Deliv. Sci. Technol. 2017, 37, 97–114. [Google Scholar] [CrossRef]
- Meissner, J.M.; Toporkiewicz, M.; Czogalla, A.; Matusewicz, L.; Kuliczkowski, K.; Sikorski, A.F. Novel antisense therapeutics delivery systems: In vitro and in vivo studies of liposomes targeted with anti-CD20 antibody. J. Control. Release Off. J. Control. Release Soc. 2015, 220, 515–528. [Google Scholar] [CrossRef] [PubMed]
- Kole, R.; Williams, T.; Cohen, L. RNA modulation, repair and remodeling by splice switching oligonucleotides. Acta Biochim. Pol. 2004, 51, 373–378. [Google Scholar] [PubMed]
- Smith, L.D.; Leme de Calais, F.; Raponi, M.; Mellone, M.; Buratti, E.; Blaydes, J.P.; Baralle, D. Novel splice-switching oligonucleotide promotes BRCA1 aberrant splicing and susceptibility to PARP inhibitor action. Int. J. Cancer 2017, 140, 1564–1570. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.Y.; Yang, Y.T.; Zhu, R.; Wang, Y.Q.; Tian, Y.; Li, X.H.; Wang, R.Q. CpG oligodeoxynucleotides discriminately enhance binding capacity of human naive B cells to Hepatitis B virus epitopes. Can. J. Microbiol. 2012, 58, 752–759. [Google Scholar] [CrossRef] [PubMed]
- Choong, O.K.; Mehrbod, P.; Tejo, B.A.; Omar, A.R. In vitro antiviral activity of circular triple helix forming oligonucleotide RNA towards Feline Infectious Peritonitis virus replication. BioMed Res. Int. 2014, 2014, 654712. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Temsamani, J.; Galbraith, W.; Jinyan, T. Pharmacokinetics of antisense oligonucleotides. Clin. Pharm. 1995, 28, 7–16. [Google Scholar] [CrossRef] [PubMed]
- GEARY, R.S.; WATANABE, T.A.; TRUONG, L.; FREIER, S.; LESNIK, E.A.; SIOUFI, N.B.; SASMOR, H.; MANOHARAN, M.; LEVIN, A.A. Pharmacokinetic Properties of 29-O-(2-Methoxyethyl)-Modified Oligonucleotide Analogs in Rats. J. Pharmacol. Exp. Ther. 2001, 296, 890–897. [Google Scholar] [PubMed]
- Zhang, R.W.; Robert, B.D.; Lu, Z.H.; Liu, T.P.; Jiang, Z.W.; Galbraith, W.M.; Agrawal, S. Pharmacokinetics and tissue distribution in rats of an oligodeoxynucleotide. Biochem. Pharmacol. 1995, 49, 929–939. [Google Scholar] [CrossRef]
- Liu, M.; Li, M.; Sun, S.; Li, B.; Du, D.; Sun, J.; Cao, F.; Li, H.; Jia, F.; Wang, T.; et al. The use of antibody modified liposomes loaded with AMO-1 to deliver oligonucleotides to ischemic myocardium for arrhythmia therapy. Biomaterials 2014, 35, 3697–3707. [Google Scholar] [CrossRef] [PubMed]
- Miteva, M.; Kirkbride, K.C.; Kilchrist, K.V.; Werfel, T.A.; Li, H.; Nelson, C.E.; Gupta, M.K.; Giorgio, T.D.; Duvall, C.L. Tuning PEGylation of mixed micelles to overcome intracellular and systemic siRNA delivery barriers. Biomaterials 2015, 38, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of nanoparticle design for overcoming biological barriers to drug delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef] [PubMed]
- Von Roemeling, C.; Jiang, W.; Chan, C.K.; Weissman, I.L.; Kim, B.Y. Breaking Down the Barriers to Precision Cancer Nanomedicine. Trends Biotechnol. 2017, 35, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.; Carver, K.; Wu, L. Albumin-based nanoconjugates for targeted delivery of therapeutic oligonucleotides. Biomaterials 2013, 34, 7939–7949. [Google Scholar] [CrossRef] [PubMed]
- Ming, X. Cellular delivery of siRNA and antisense oligonucleotides via receptor-mediated endocytosis. Expert Opin. Drug Deliv. 2011, 8, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Fattal, E.; Barratt, G. Nanotechnologies and controlled release systems for the delivery of antisense oligonucleotides and small interfering RNA. Br. J. Pharmacol. 2009, 157, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Dudgeon, N.; Shen, L.; Wang, J.H. Chemical modification of gene silencing oligonucleotides for drug discovery and development. Drug Discov. Today 2005, 10, 587–593. [Google Scholar] [CrossRef]
- Boisguerin, P.; Deshayes, S.; Gait, M.J.; O’Donovan, L.; Godfrey, C.; Betts, C.A.; Wood, M.J.; Lebleu, B. Delivery of therapeutic oligonucleotides with cell penetrating peptides. Adv. Drug Deliv. Rev. 2015, 87, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Amarzguioui, M.; Holen, T.; Babaie, E.; Prydz, H. Tolerance for mutations and chemical modifications in a siRNA. Nucleic Acid Res. 2003, 31, 589–595. [Google Scholar] [CrossRef] [PubMed]
- YI, C.Q.; Pan, T. Cellular dynamics of RNA modification. Acc. Chem. Res. 2011, 44, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Mook, O.R.; Baas, F.; De Wissel, F.B.; Fluiter, K. Evaluation of locked nucleic acid-modified small interfering RNA in vitro and in vivo. Mol. Cancer Ther. 2007, 6, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Braasch, D.A.; Paroo, Z.; Constantinescu, A.; Ren, G.; Oz, O.K.; Mason, R.P.; Corey, D.R. Biodistribution of phosphodiester and phosphorothioate siRNA. Bioorg. Med. Chem. Lett. 2004, 14, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Soutschek, J.; Akinc, A.; Bramlage, B.; Charisse, K.; Constien, R.; Donoghue, M.; Elbashir, S.; Vornlocher, H.P. Therapeutic silencing of an endogenous gene by systemic administration of modified siRNAs. Nature 2004, 432, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Allerson, C.R.; Sioufi, N.; Jarres, R.; Prakash, T.P.; Naik, N.; Berdeja, A.; Wanders, L.; Griffey, R.H.; Swayze, E.E.; Bhat, B. Fully2′-modified oligonucleotide duplexes with improved in vitro potency and stability compared to unmodified small interfering RNA. J. Med. Chem. 2005, 48, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Simeon, F.; Morris, M.C.; Heitz, F.; Divita, G. Insight into the mechanism of the peptide-based gene delivery system MPG Implications for delivery of siRNA into mammalian cells. Nucleic Acid Res. 2003, 31, 2717–2724. [Google Scholar] [CrossRef]
- Jing, H.; Cheng, W.; Li, S.; Wu, B.; Leng, X.; Xu, S.; Tian, J. Novel cell-penetrating peptide-loaded nanobubbles synergized with ultrasound irradiation enhance EGFR siRNA delivery for triple negative Breast cancer therapy. Colloids Surf. B Biointerfaces 2016, 146, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Lin, W.; Li, M.; Yang, Y.; Deng, J.; Liu, H.; Chen, Y.; Fu, X.; Liu, H.; Yang, Y. Efficient siRNA Delivery Using Novel Cell-Penetrating Peptide-siRNA Conjugate-Loaded Nanobubbles and Ultrasound. Ultrasound Med. Biol. 2016, 42, 1362–1374. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, B.; Tripathi, S.; Pandey, V.N. Single acute-dose and repeat-doses toxicity of anti-HIV-1 PNA TAR-penetratin conjugate after intraperitoneal administration to mice. Oligonucleotides 2008, 18, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Yuba, E.; Nakajima, Y.; Tsukamoto, K.; Iwashita, S.; Kojima, C.; Harada, A.; Kono, K. Effect of unsaturated alkyl chains on transfection activity of poly(amidoamine) dendron-bearing lipids. J. Control. Release Off. J. Control. Release Soc. 2012, 160, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yang, Y.; Lin, W.; Liu, H.; Liu, H.; Yang, Y.; Chen, Y.; Fu, X.; Deng, J. Cell-penetrating peptide-siRNA conjugate loaded YSA-modified nanobubbles for ultrasound triggered siRNA delivery. Colloids Surf. B Biointerfaces 2015, 136, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Arami, S.; Mahdavi, M.; Rashidi, M.R.; Yekta, R.; Rahnamay, M.; Molavi, L.; Hejazi, M.S.; Samadi, N. Apoptosis induction activity and molecular docking studies of survivin siRNA carried by Fe3O4-PEG-LAC-chitosan-PEI nanoparticles in MCF-7 human breast cancer cells. J. Pharm. Biomed. Anal. 2017, 142, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, A.; Yang, Z.; Wang, X.; Chang, L.; Chen, Z.; James Lee, L. Application of DODMA and Derivatives in Cationic Nanocarriers for Gene Delivery. Curr. Org. Chem. 2016, 20, 1813–1819. [Google Scholar] [CrossRef]
- Rabbani, P.S.; Zhou, A.; Borab, Z.M.; Frezzo, J.A.; Srivastava, N.; More, H.T.; Rifkin, W.J.; David, J.A.; Berens, S.J.; Chen, R.; et al. Novel lipoproteoplex delivers Keap1 siRNA based gene therapy to accelerate diabetic wound healing. Biomaterials 2017, 132, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Munye, M.M.; Tagalakis, A.D.; Manunta, M.D.; Hart, S.L. The role of the helper lipid on the DNA transfection efficiency of lipopolyplex formulations. Sci. Rep. 2014, 4, 7107. [Google Scholar] [CrossRef] [PubMed]
- Remaut, K.; Lucas, B.; Braeckmans, K.; Demeester, J.; De Smedt, S.C. Pegylation of liposomes favours the endosomal degradation of the delivered phosphodiester oligonucleotides. J. Control. Release Off. J. Control. Release Soc. 2007, 117, 256–266. [Google Scholar] [CrossRef] [PubMed]
- Themis, M.; Forbes, S.J.; Chan, L.; Cooper, R.G.; Etheridge, C.J.; Miller, A.D.; Hodgson, H.J.F.; Coutelle, C. Enhanced in vitro and in vivo gene delivery using cationic agent complexed retrovirus vectors. Gene Ther. 1998, 5, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Gwak, S.J.; Yun, Y.; Yoon, D.H.; Kim, K.N.; Ha, Y. Therapeutic Use of 3beta-[N-(N′,N′-Dimethylaminoethane) Carbamoyl] Cholesterol-Modified PLGA Nanospheres as Gene Delivery Vehicles for Spinal Cord Injury. PLoS ONE 2016, 11, e0147389. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Zhu, P.; Lee, S.K.; Chowdhury, D.; Kussman, S.; Dykxhoorn, D.M.; Feng, Y.; Palliser, D.; Weiner, D.B.; Shankar, P.; et al. Antibody mediated in vivo delivery of small interfering RNAs via cell-surface receptors. Nat. Biotechnol. 2005, 23, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, E.; Freitas, J.; Loureiro, A.; Nogueira, P.; Gomes, A.C.; Preto, A.; Carmo, A.M.; Moreira, A.; Cavaco-Paulo, A. Neutral PEGylated liposomal formulation for efficient folate-mediated delivery of MCL1 siRNA to activated macrophages. Colloids Surf. B Biointerfaces 2017, 155, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Fan, H.; Levorse, D.A.; Crocker, L.S. Ionization behavior of amino lipids for siRNA delivery: Determination of ionization constants, SAR, and the impact of lipid pKa on cationic lipid-biomembrane interactions. Langmuir ACS J. Surf. Colloids 2011, 27, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.M.; Zhang, Y.F.; Xie, Y.D.; Cai, Y.F.; Li, B.Y.; Li, W.; Zeng, L.Y.; Li, Y.L.; Yu, R.T. Hypoxia-responsive ionizable liposome delivery siRNA for glioma therapy. Int. J. Nanomed. 2017, 12, 1065–1083. [Google Scholar] [CrossRef] [PubMed]
- Habrant, D.; Peuziat, P.; Colombani, T.; Dallet, L.; Gehin, J.; Goudeau, E.; Evrard, B.; Lambert, O.; Haudebourg, T.; Pitard, B. Design of Ionizable Lipids To Overcome the Limiting Step of Endosomal Escape: Application in the Intracellular Delivery of mRNA, DNA, and siRNA. J. Med. Chem 2016, 59, 3046–3062. [Google Scholar] [CrossRef] [PubMed]
- Amjad, M.W.; Kesharwani, P.; Mohd Amin, M.C.I.; Iyer, A.K. Recent advances in the design, development, and targeting mechanisms of polymeric micelles for delivery of siRNA in cancer therapy. Progr. Polym. Sci. 2017, 64, 154–181. [Google Scholar] [CrossRef]
- Yin, T.; Wang, L.; Yin, L.; Zhou, J.; Huo, M. Co-delivery of hydrophobic paclitaxel and hydrophilic AURKA specific siRNA by redox-sensitive micelles for effective treatment of breast cancer. Biomaterials 2015, 61, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Balk, M.; Deng, Z.; Wischke, C.; Gossen, M.; Behl, M.; Ma, N.; Lendlein, A. Engineering biodegradable micelles of polyethylenimine-based amphiphilic block copolymers for efficient DNA and siRNA delivery. J. Control. Release Off. J. Control. Release Soc. 2016, 242, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zheng, M.; Gong, P.; Deng, J.; Yi, H.; Zhang, P.; Zhang, Y.; Liu, P.; Ma, Y.; Cai, L. Polypeptide cationic micelles mediated co-delivery of docetaxel and siRNA for synergistic tumor therapy. Biomaterials 2013, 34, 3431–3438. [Google Scholar] [CrossRef] [PubMed]
- Gwak, S.J.; Nice, J.; Zhang, J.; Green, B.; Macks, C.; Bae, S.; Webb, K.; Lee, J.S. Cationic, amphiphilic copolymer micelles as nucleic acid carriers for enhanced transfection in rat spinal cord. Acta Biomater. 2016, 35, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Q.; Hu, Q.; Li, Y.; Tang, G.; Chu, P.K. Restoration of chemosensitivity by multifunctional micelles mediated by P-gp siRNA to reverse MDR. Biomaterials 2014, 35, 8621–8634. [Google Scholar] [CrossRef] [PubMed]
- Son, S.; Song, S.; Lee, S.J.; Min, S.; Kim, S.A.; Yhee, J.Y.; Huh, M.S.; Chan Kwon, I.; Jeong, S.Y.; Byun, Y.; et al. Self-crosslinked human serum albumin nanocarriers for systemic delivery of polymerized siRNA to tumors. Biomaterials 2013, 34, 9475–9485. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Yin, Y.; Huang, C.; Pan, W.; Liang, D. Encapsulation of RNA by negatively charged human serum albumin via physical interactions. Sci. China Chem. 2016, 60, 130–135. [Google Scholar] [CrossRef]
- Shukla, A.K.; Iravani, S. Metallic nanoparticles: Green synthesis and spectroscopic characterization. Environ. Chem. Lett. 2017, 15, 223–231. [Google Scholar] [CrossRef]
- Vimbela, G.V.; Ngo, S.M.; Fraze, C.; Yang, L.; Stout, D.A. Antibacterial properties and toxicity from metallic nanomaterials. Int. J. Nanomed. 2017, 12, 3941–3965. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, M.; Douziech Eyrolles, L.; Alkarib, S.Y.; Herve-Aubert, K.; Ben Djemaa, S.; Marchais, H.; Chourpa, I.; David, S. siRNA delivery system based on magnetic nanovectors: Characterization and stability evaluation. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2017, 106, 287–293. [Google Scholar] [CrossRef] [PubMed]
- Rahme, K.; Guo, J.; Holmes, J.D.; O’Driscoll, C.M. Evaluation of the physicochemical properties and the biocompatibility of polyethylene glycol-conjugated gold nanoparticles: A formulation strategy for siRNA delivery. Colloids Surf. B Biointerfaces 2015, 135, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Vinhas, R.; Fernandes, A.R.; Baptista, P.V. Gold Nanoparticles for BCR-ABL1 Gene Silencing: Improving Tyrosine Kinase Inhibitor Efficacy in Chronic Myeloid Leukemia. Mol. Ther. Nucleic Acids 2017, 7, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, W.; Li, N.; Zhao, Z.; Mou, Z.; Yang, E.; Wang, W. Silver nanoparticles-quercetin conjugation to siRNA against drug-resistant Bacillus subtilis for effective gene silencing: In vitro and in vivo. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 63, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Yhee, J.Y.; Kim, S.H.; Kwon, I.C.; Kim, K. Biocompatible gelatin nanoparticles for tumor-targeted delivery of polymerized siRNA in tumor-bearing mice. J. Control. Release Off. J. Control. Release Soc. 2013, 172, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Srikar, R.; Suresh, D.; Zambre, A.; Taylor, K.; Chapman, S.; Leevy, M.; Upendran, A.; Kannan, R. Targeted nanoconjugate co-delivering siRNA and tyrosine kinase inhibitor to KRAS mutant NSCLC dissociates GAB1-SHP2 post oncogene knockdown. Sci. Rep. 2016, 6, 30245. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Uludag, H. Nucleic-acid based gene therapeutics: Delivery challenges and modular design of nonviral gene carriers and expression cassettes to overcome intracellular barriers for sustained targeted expression. J. Drug Target. 2012, 20, 301–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tang, J.; Fu, L.; Ran, R.; Liu, Y.; Yuan, M.; He, Q. A pH-responsive alpha-helical cell penetrating peptide-mediated liposomal delivery system. Biomaterials 2013, 34, 7980–7993. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.Y.; Hendzel, M.; Uludag, H. Improved transfection efficiency of an aliphatic lipid substituted 2 kDa polyethylenimine is attributed to enhanced nuclear association and uptake in rat bone marrow stromal cell. J. Gene Med. 2011, 13, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Lam, J.K.; Armes, S.P.; Lewis, A.L.; Stolnik, S. Folate conjugated phosphorylcholine-based polycations for specific targeting in nucleic acids delivery. J. Drug Target. 2009, 17, 512–523. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
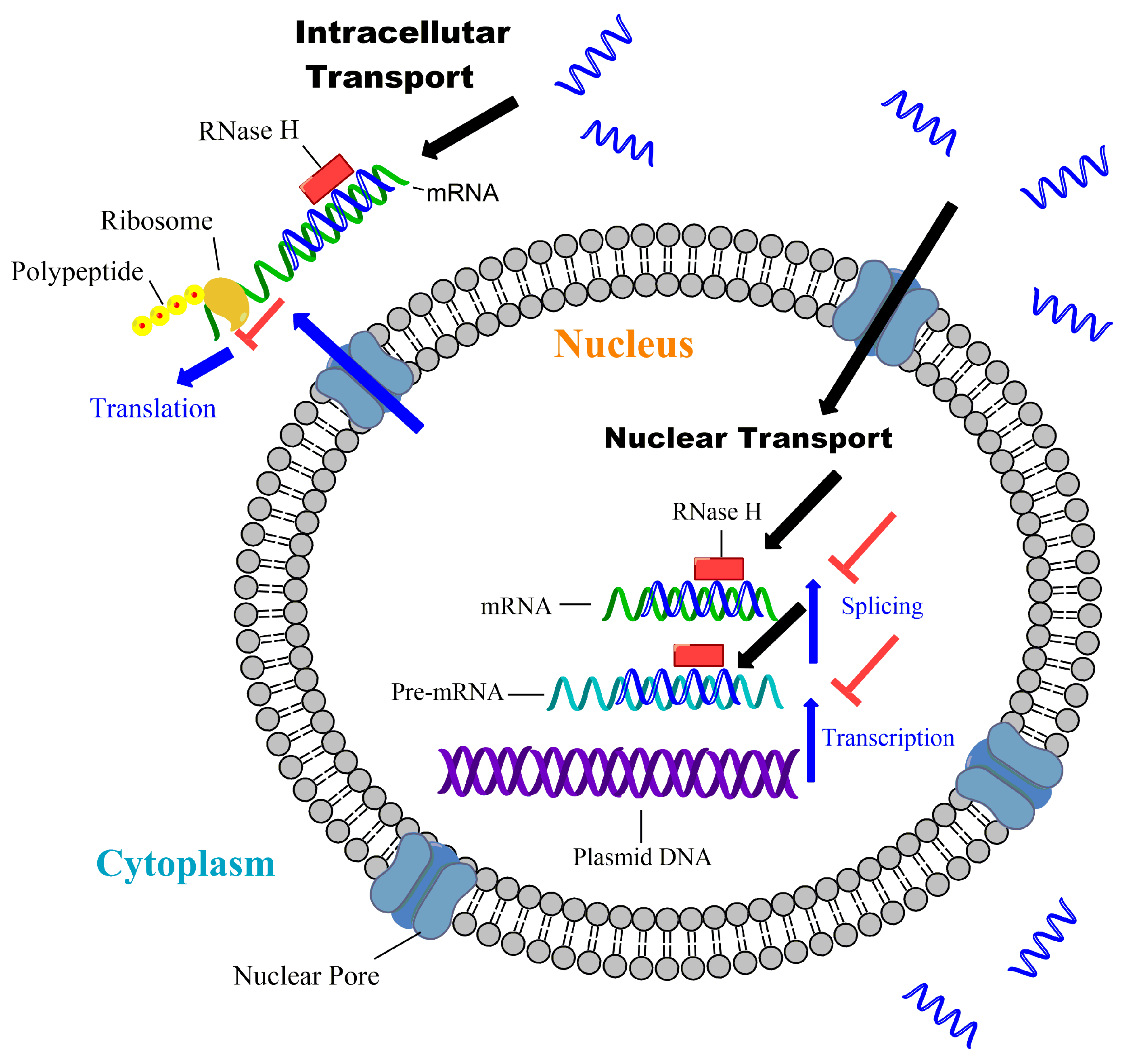{kind=link}
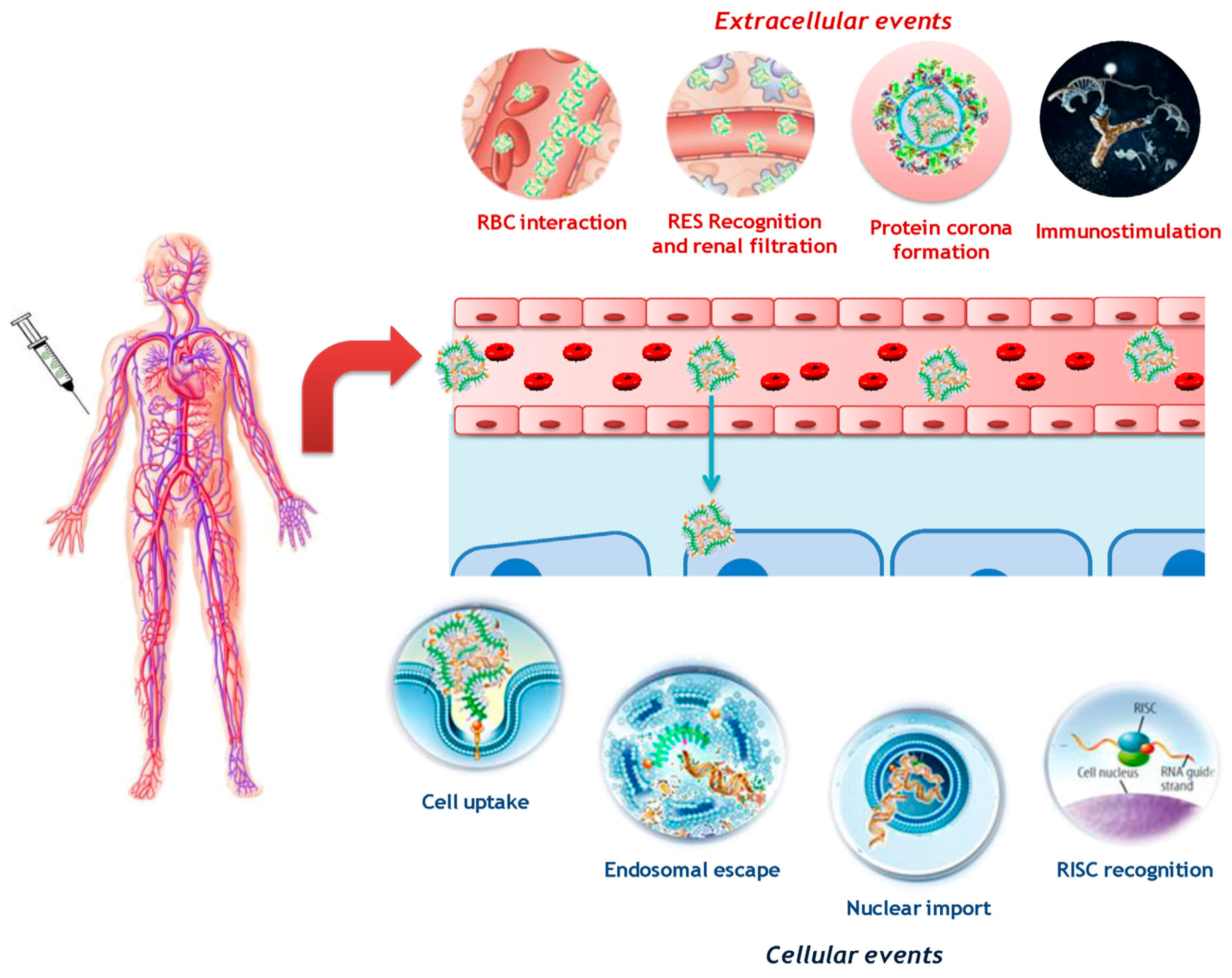{kind=link}
| Trade Name | Time to Market | Company | Indication |
|---|---|---|---|
| Vitravene | 1998 | Isis (Ionis) | Cytomegalovirus-induced retinitis |
| Macugen | 2004 | Pfizer/Eyetech | Age-related macular degeneration |
| Kynamro | 2013 | Sanofi/Isis (Ionis) | Familial hypercholesterolemia |
| Exondys 51 | 2016 | Sarepta Therapeutics | Duchenne muscular dystrophy |
| Spinraza | 2016 | Biogen/Ionis | Spinal muscular atrophy |
| Defibrotide | 2016 | Jazz Pharma | Severe hepatic veno-occlusive disease |
| Category | Mechanism of Action | Mainly Applications | Reference |
|---|---|---|---|
| Antisense | Hybridizes with mRNA and inhibit ribosome’s binding | Cancer treatment congenital genetic disease and acquired immune disease treatment | [43] |
| ON | Coupled with RNase H and promote targeting nucleic acid’s degradation | ||
| Splice switching ON | Inhibits or promotes exon insertion to modify pre-mRNA’s splicing pattern | RNA repairing and modulation | [44,45] |
| CpG-ON | Triggers cells to express toll-like receptor 9, and induces innate immune response | Vaccine adjuvants | [46] |
| Triple-helix-forming ON | Inserts into double stranded DNA to inhibit mRNA transcription | Virus infection treatment cancer treatment | [47] |
| Category | Contents | Strength | Shortage | Reference |
|---|---|---|---|---|
| Diester modification | Phosphorothioate | Increase cellular uptake, bioavailability and resistance to nucleases | Cytotoxicity increases, gene silencing effect decreases | [58,59] |
| Ribose modification | 2′-O-Me, 2′-O-A, 2′-F | Enhanced stability | Gene silencing effect decreases | [24,60] |
| Base modification | Adenine methylation and deamination. cytosine methylation, hydroxy methylation and carboxy substitution, Guanine oxidation | Improved gene silencing effect | Functional groups change easily through modification | [61] |
| ON analogues replacement | Peptide nucleic acid, locked nucleic acid, morpholino phosphamide | Good targeting effect, nuclease resistance | Binding affinity decreases | [57] |
| Groups | Materials | Strength | Limitations | Normal Method | Reference |
|---|---|---|---|---|---|
| Cationic LPs | DOTAP | Positive charge, high encapsulation efficiency, easily access to cells, endosomal escape | Adsorption of anionic serum proteins, fast clearance by RES | Neutral lipid and pegylated modification | [73,74,77,78] |
| DODMA | |||||
| DOGS | |||||
| DC-Chol | |||||
| Neutral LPs | PC | Good biocompatibility and pharmacokinetic characteristics | Low encapsulation efficiency | Adding cationic materials | [7,75,79,80] |
| Chol | |||||
| DOPE | |||||
| Ionizable LPs | DODMA | Transformable charge, high transfection efficiency, broad prospects | Improve design ideas | [7,81,82,83] | |
| DODAP |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Zhao, Y.; Zhao, X.; Lee, R.J.; Teng, L.; Zhou, C. Enhancing the Therapeutic Delivery of Oligonucleotides by Chemical Modification and Nanoparticle Encapsulation. Molecules 2017, 22, 1724. https://doi.org/10.3390/molecules22101724
Sun Y, Zhao Y, Zhao X, Lee RJ, Teng L, Zhou C. Enhancing the Therapeutic Delivery of Oligonucleotides by Chemical Modification and Nanoparticle Encapsulation. Molecules. 2017; 22(10):1724. https://doi.org/10.3390/molecules22101724
Chicago/Turabian StyleSun, Yating, Yarong Zhao, Xiuting Zhao, Robert J. Lee, Lesheng Teng, and Chenguang Zhou. 2017. "Enhancing the Therapeutic Delivery of Oligonucleotides by Chemical Modification and Nanoparticle Encapsulation" Molecules 22, no. 10: 1724. https://doi.org/10.3390/molecules22101724