
Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks




Abstract
:
1. Introduction
2. Definition and Chemical Structures
3. Natural Sources of Methylxanthines and Biosynthesis
4. Extraction, Identification and Quantification
5. Molecular Targets and Structure-Activity Relationships
6. Physiological and Health Benefits
7. Drawbacks and Toxicity
8. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | amyloid-β-peptide |
| AMP | adenosine monophosphate |
| cAMP | cyclic adenosine monophosphate |
| CE | capillary electrophoresis |
| CNS | central nervous system |
| COPD | chronic obstructive pulmonary disease |
| GC | gas chromatography |
| GC-MS | gas chromatography-mass spectrometry |
| GABA | gamma-aminobutyric acid |
| GMP | guanosine monophosphate |
| HDL | high-density lipoprotein |
| HPLC | high-performance liquid chromatography |
| LD50 | median lethal dose |
| LDL | low-density lipoprotein |
| MRM | multiple reaction monitoring |
| MS | mass spectrometry |
| PDE | phosphodiesterase |
| SAM | S-adenosyl-l-methionine |
| TLC | thin-layer chromatography |
References
- Stasolla, C.; Katahira, R.; Thorpe, T.A.; Ashihara, H. Purine and pyrimidine nucleotide metabolism in higher plants. J. Plant Physiol. 2003, 160, 1271–1295. [Google Scholar] [CrossRef] [PubMed]
- Zrenner, R.; Stitt, M.; Sonnewald, U.; Boldt, R. Pyrimidine and purine biosynthesis and degradation in plants. Annu. Rev. Plant Biol. 2006, 57, 805–836. [Google Scholar] [CrossRef] [PubMed]
- Glantzounis, G.K.; Tsimoyiannis, E.C.; Kappas, A.M.; Galaris, D.A. Uric Acid and Oxidative Stress. Curr. Pharm. Des. 2005, 11, 4145–4151. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, R.G.; Armstrong, J.W.; Campbell, E. Pest Control: Caffeine as a repellent for slugs and snails. Nature 2002, 417, 915–916. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, P.M.; Ulmer-Dufek, J.; Baumann, T.W. Purine alkaloid formation in buds and developing leaflets of Coffea arabica: Expression of an optimal defence strategy? Phytochemistry 1986, 25, 613–616. [Google Scholar] [CrossRef]
- Wolf, A.; Bray, G.A.; Popkin, B.M. A short history of beverages and how our body treats them. Obes. Rev. 2008, 9, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Sotelo, A.; Soleri, D.; Wacher, C.; Sánchez-Chinchillas, A.; Argote, R.M. Chemical and nutritional composition of Tejate, a traditional maize and cacao beverage from the central valleys of Oaxaca, Mexico. Plant Foods Hum. Nutr. 2012, 67, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Stavric, B. Methylxanthines: Toxicity to humans. 1. Theophylline. Food Chem. Toxicol. 1988, 26, 541–565. [Google Scholar] [CrossRef]
- Ashihara, H.; Sano, H.; Crozier, A. Caffeine and related purine alkaloids: Biosynthesis, catabolism, function and genetic engineering. Phytochemistry 2008, 69, 841–856. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A.; Daval, J.-L.; Debry, G. Caffeine and the central nervous system: Mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res. Rev. 1992, 17, 139–170. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar] [PubMed]
- Jacobson, B.H.; Weber, M.D.; Claypool, L.; Hunt, L.E. Effect of caffeine on maximal strength and power in élite male athletes. Br. J. Sports Med. 1992, 26, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Anselme, F.; Collomp, K.; Mercier, B.; Ahmaïdi, S.; Prefaut, C. Caffeine increases maximal anaerobic power and blood lactate concentration. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 65, 188–191. [Google Scholar] [CrossRef] [PubMed]
- Jackman, M.; Wendling, P.; Friars, D.; Graham, T.E. Metabolic, catecholamine, and endurance responses to caffeine during intense exercise. J. Appl. Physiol. 1996, 81, 1658–1663. [Google Scholar] [PubMed]
- Graham, T.E.; Spriet, L.L. Performance and metabolic responses to a high caffeine dose during prolonged exercise. J. Appl. Physiol. 1991, 71, 2292–2298. [Google Scholar] [PubMed]
- Kovacs, E.M.R.; Stegen, J.H.C.H.; Brouns, F. Effect of caffeinated drinks on substrate metabolism, caffeine excretion, and performance. J. Appl. Physiol. 1998, 85, 709–715. [Google Scholar] [PubMed]
- Greer, F.; Friars, D.; Graham, T.E. Comparison of caffeine and theophylline ingestion: Exercise metabolism and endurance. J. Appl. Physiol. 2000, 89, 1837–1844. [Google Scholar] [PubMed]
- Chapman, R.F.; Mickleborough, T.D. The effects of caffeine on ventilation and pulmonary function during exercise: An often-overlooked response. Phys. Sportsmed. 2009, 37, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Theophylline. Am. J. Respir. Crit. Care Med. 2013, 188, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.; Jackson, E.; Mi, Z.; McCombs, J.; Carcillo, J. Low-dose theophylline increases urine output in diuretic-dependent critically ill children. Intensive Care Med. 1998, 24, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Salihović, M.; Huseinović, S.; Špirtović-Halilović, S.; Osmanović, A.; Dedić, A.; Ašimović, Z.; Završnik, D. DFT study and biological activity of some methylxanthines. Bull. Chem. Technol. Bosnia Herzeg. 2014, 42, 31–36. [Google Scholar]
- Talik, P.; Krzek, J.; Ekiert, R.J. Analytical techniques used for determination of methylxanthines and their analogues—Recent advances. Sep. Purif. Rev. 2012, 41, 1–61. [Google Scholar] [CrossRef]
- McCall, A.L.; Millington, W.R.; Wurtman, R.J. Blood-brain barrier transport of caffeine: Dose-related restriction of adenine transport. Life Sci. 1982, 31, 2709–2715. [Google Scholar] [CrossRef]
- Beale, J.M., Jr. Wilson and Gisvold’s Textbook of Organic Medicinal and Pharmaceutical Chemistry, 12th ed.; Lippincott Williams & Wilkins, Wolters Kuwer: Philadelphia, PA, USA, 2011. [Google Scholar]
- Pavia, D.L. Coffee, Tea, or Cocoa. A trio of experiments including the isolation of theobromine from cocoa. J. Chem. Educ. 1973, 50, 791. [Google Scholar] [CrossRef] [PubMed]
- Gardenhire, D.S. Rau’s Respiratory Care Pharmacology, 9th ed.; Elsevier Health Sciences: St. Louis, MO, USA, 2016; p. 140. [Google Scholar]
- Ashihara, H.; Suzuki, T. Distribution and biosynthesis of caffeine in plants. Front. Biosci. 2004, 9, 1864–1876. [Google Scholar] [CrossRef] [PubMed]
- Ashihara, H.; Crozier, A. Biosynthesis and metabolism of caffeine and related purine alkaloids in plants. In Advances in Botanical Research; Callow, J.A., Ed.; Academic Press: New York, NY, USA, 1999; Volume 30, pp. 117–205. [Google Scholar]
- Mazzafera, P.; Carvalho, A. Breeding for low seed caffeine content of coffee (Coffea L.) by interspecific hybridization. Euphytica 1991, 59, 55–60. [Google Scholar]
- Lin, Y.-S.; Tsai, Y.-J.; Tsay, J.-S.; Lin, J.-K. Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J. Agric. Food Chem. 2003, 51, 1864–1873. [Google Scholar] [CrossRef]
- Fernandez, P.L.; Martin, M.J.; Gonzalez, A.G.; Pablos, F. HPLC determination of catechins and caffeine in tea. Differentiation of green, black and instant teas. Analyst 2000, 125, 421–425. [Google Scholar] [CrossRef]
- Stavric, B. Methylxanthines: Toxicity to humans. 3. Theobromine, paraxanthine and the combined effects of methylxanthines. Food Chem. Toxicol. 1988, 26, 725–733. [Google Scholar] [CrossRef]
- Baumann, T.W.; Schulthess, B.H.; Hänni, K. Guaraná (Paullinia cupana) rewards seed dispersers without intoxicating them by caffeine. Phytochemistry 1995, 39, 1063–1070. [Google Scholar] [CrossRef]
- Weckerle, C.S.; Stutz, M.A.; Baumann, T.W. Purine alkaloids in Paullinia. Phytochemistry 2003, 64, 735–742. [Google Scholar] [CrossRef]
- Kretschmar, J.A.; Baumann, T.W. Caffeine in Citrus flowers. Phytochemistry 1999, 52, 19–23. [Google Scholar] [CrossRef]
- Atawodi, S.E.-O.; Pfundstein, B.; Haubner, R.; Spiegelhalder, B.; Bartsch, H.; Owen, R.W. Content of polyphenolic compounds in the nigerian stimulants Cola nitida ssp. alba, Cola nitida ssp. rubra A. Chev, and Cola acuminata Schott & Endl and their antioxidant capacity. J. Agric. Food Chem. 2007, 55, 9824–9828. [Google Scholar] [PubMed]
- Frary, C.D.; Johnson, R.K.; Wang, M.Q. Food sources and intakes of caffeine in the diets of persons in the United States. J. Am. Diet. Assoc. 2005, 105, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.M.; Marshman, J.A.; Schwieder, M.; Berg, R. Caffeine content of beverages as consumed. Can. Med. Assoc. J. 1976, 114, 205–208. [Google Scholar] [PubMed]
- Moderno, P.M.; Carvalho, M.; Silva, B.M. Recent patents on Camellia sinensis: Source of health promoting compounds. Recent Pat. Food Nutr. Agric. 2009, 1, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P.; Henning, S.M.; Niu, Y.; Lee, R.; Scheuller, H.S.; Heber, D. Catechin and caffeine content of green tea dietary supplements and correlation with antioxidant capacity. J. Agric. Food Chem. 2006, 54, 1599–1603. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Jerónimo, C.; Valentão, P.; Andrade, P.B.; Silva, B.M. Green tea: A promising anticancer agent for renal cell carcinoma. Food Chem. 2010, 122, 49–54. [Google Scholar] [CrossRef]
- Dias, T.R.; Tomás, G.; Teixeira, N.F.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. White tea (Camellia sinensis L.): Antioxidant properties and beneficial health effects. Int. J. Food Sci. Nutr. Diet. 2013, 11, 1–15. [Google Scholar]
- Martins, A.D.; Alves, M.G.; Bernardino, R.L.; Dias, T.R.; Silva, B.M.; Oliveira, P.F. Effect of white tea (Camellia sinensis (L.)) extract in the glycolytic profile of Sertoli cell. Eur. J. Nutr. 2013, 53, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Unachukwu, U.J.; Ahmed, S.; Kavalier, A.; Lyles, J.T.; Kennelly, E.J. White and green teas (Camellia sinensis var. sinensis): Variation in phenolic, methylxanthine, and antioxidant profiles. J. Food Sci. 2010, 75, C541–C548. [Google Scholar] [CrossRef] [PubMed]
- Boros, K.; Jedlinszki, N.; Csupor, D. Theanine and caffeine content of infusions prepared from commercial tea samples. Pharmacogn. Mag. 2016, 12, 75–79. [Google Scholar] [PubMed]
- Crozier, T.W.M.; Stalmach, A.; Lean, M.E.J.; Crozier, A. Espresso coffees, caffeine and chlorogenic acid intake: Potential health implications. Food Funct. 2012, 3, 30–33. [Google Scholar] [CrossRef] [PubMed]
- McCusker, R.R.; Fuehrlein, B.; Goldberger, B.A.; Gold, M.S.; Cone, E.J. Caffeine content of decaffeinated coffee. J. Anal. Toxicol. 2006, 30, 611–613. [Google Scholar] [CrossRef] [PubMed]
- Astill, C.; Birch, M.R.; Dacombe, C.; Humphrey, P.G.; Martin, P.T. Factors affecting the caffeine and polyphenol contents of black and green tea infusions. J. Agric. Food Chem. 2001, 49, 5340–5347. [Google Scholar] [CrossRef] [PubMed]
- Blauch, J.L.; Tarka, S.M. HPLC determination of caffeine and theobromine in coffee, tea, and instant hot cocoa Mixes. J. Food Sci. 1983, 48, 745–747. [Google Scholar] [CrossRef]
- Bispo, M.S.; Veloso, M.C.C.; Pinheiro, H.L.C.; de Oliveira, R.F.S.; Reis, J.O.N.; de Andrade, J.B. Simultaneous determination of caffeine, theobromine, and theophylline by high-performance liquid chromatography. J. Chromatogr. Sci. 2002, 40, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Hicks, M.B.; Hsieh, Y.H.P.; Bell, L.N. Tea preparation and its influence on methylxanthine concentration. Food Res. Int. 1996, 29, 325–330. [Google Scholar] [CrossRef]
- Alañón, M.E.; Castle, S.M.; Siswanto, P.J.; Cifuentes-Gómez, T.; Spencer, J.P.E. Assessment of flavanol stereoisomers and caffeine and theobromine content in commercial chocolates. Food Chem. 2016, 208, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.A.; Campos-Giménez, E.; Jiménez Alvarez, D.; Rytz, A.; Nagy, K.; Williamson, G. Predictive relationship between polyphenol and nonfat cocoa solids content of chocolate. J. Agric. Food Chem. 2008, 56, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Tokusoglu, Ö.; Ünal, K.M. Optimized method for simultaneous determination of catechin, gallic acid, and methylxanthine compounds in chocolate using RP-HPLC. Eur. Food Res. Technol. 2002, 215, 340–346. [Google Scholar] [CrossRef]
- Ashihara, H.; Kato, M.; Crozier, A. Distribution, biosynthesis and catabolism of methylxanthines in plants. In Methylxanthines; Springer Berlin Heidelberg: Berlin, Germany; Heidelberg, Germany, 2011; pp. 11–31. [Google Scholar]
- Misako, K.; Kouichi, M. Caffeine synthase and related methyltransferases in plants. Front. Biosci. 2004, 9, 1833–1842. [Google Scholar] [PubMed]
- Koshiishi, C.; Kato, A.; Yama, S.; Crozier, A.; Ashihara, H. A new caffeine biosynthetic pathway in tea leaves: Utilisation of adenosine released from the S-adenosyl-l-methionine cycle. FEBS Lett. 2001, 499, 50–54. [Google Scholar] [CrossRef]
- Zajac, M.A.; Zarkzewski, A.G.; Kowal, M.G.; Narayan, S. A Novel Method of Caffeine Synthesis from Uracil; Taylor & Francis: Philadelphia, PA, USA, 2003; Volume 33, p. 7. [Google Scholar]
- Kumar, N.S.; Chandran, T.B.; Namratha, S.; Bharat, B.R.; Kumar, C.H.P.; Kishore, V. Production of theobromine from Pseudomonas sp. Int. Res. J. Pharm. 2013, 4. [Google Scholar]
- Nanjundaiah, S.; Bhatt, P.; Rastogi, N.K.; Thakur, M.S. Response surface optimization for decaffeination and theophylline production by Fusarium solani. Appl. Biochem. Biotechnol. 2015, 178, 58–75. [Google Scholar] [CrossRef] [PubMed]
- Begas, E.; Kouvaras, E.; Tsakalof, A.; Papakosta, S.; Asprodini, E.K. In vivo evaluation of CYP1A2, CYP2A6, NAT-2 and xanthine oxidase activities in a Greek population sample by the RP-HPLC monitoring of caffeine metabolic ratios. Biomed. Chromatogr. 2007, 21, 190–200. [Google Scholar] [CrossRef] [PubMed]
- El-Yazigi, A.; Shabib, S.; Al-Rawithi, S.; Yusuf, A.; Legayada, E.S.; Al-Humidan, A. Salivary clearance and urinary metabolic pattern of caffeine in healthy children and in pediatric patients with hepatocellular diseases. J. Clin. Pharmacol. 1999, 39, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Krul, C.; Hageman, G. Analysis of urinary caffeine metabolites to assess biotransformation enzyme activities by reversed-phase high-performance liquid chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1998, 709, 27–34. [Google Scholar] [CrossRef]
- Bendriss, E.-K.; Markoglou, N.; Wainer, I.W. Liquid chromatographic method for the simultaneous determination of caffeine and fourteen caffeine metabolites in urine. J. Chromatogr. B Biomed. Sci. Appl. 2000, 746, 331–338. [Google Scholar] [CrossRef]
- Van Soeren, M.; Mohr, T.; Kjaer, M.; Graham, T.E. Acute effects of caffeine ingestion at rest in humans with impaired epinephrine responses. J. Appl. Physiol. 1996, 80, 999–1005. [Google Scholar] [PubMed]
- Newton, R.; Broughton, L.J.; Lind, M.J.; Morrison, P.J.; Rogers, H.J.; Bradbrook, I.D. Plasma and salivary pharmacokinetics of caffeine in man. Eur. J. Clin. Pharmacol. 1981, 21, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Georga, K.A.; Samanidou, V.F.; Papadoyannis, I.N. IImproved micro-method for the HPLC analysis of caffeine and its demethylated metabolites in human biological fluids after SPE. J. Liq. Chromatogr. Relat. Technol. 2000, 23, 1523–1537. [Google Scholar] [CrossRef]
- Fenske, M. Caffeine determination in human saliva and urine by TLC and ultraviolet absorption densitometry. Chromatographia 2006, 65, 233–238. [Google Scholar] [CrossRef]
- Emara, S. Simultaneous determination of caffeine, theophylline and theobromine in human plasma by on-line solid-phase extraction coupled to reversed-phase chromatography. Biomed. Chromatogr. 2004, 18, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Beth Welsh, M.; Klooster, M.J.; Balistreri, W.F.; Lim, C.K. Rapid high-performance liquid chromatography assay for salivary and serum caffeine following an oral load. J. Chromatogr. A 1987, 385, 267–274. [Google Scholar] [CrossRef]
- Srdjenovic, B.; Djordjevic-Milic, V.; Grujic, N.; Injac, R.; Lepojevic, Z. Simultaneous HPLC determination of caffeine, theobromine, and theophylline in food, drinks, and herbal products. J. Chromatogr. Sci. 2008, 46, 144–149. [Google Scholar] [CrossRef] [PubMed]
- González-Nuñez, L.N.; Cañizares-Macías, M.P. Focused microwaves-assisted extraction of theobromine and caffeine from cacao. Food Chem. 2011, 129, 1819–1824. [Google Scholar] [CrossRef]
- Saldaña, M.D.A.; Zetzl, C.; Mohamed, R.S.; Brunner, G. Extraction of Methylxanthines from Guaraná Seeds, Maté Leaves, and Cocoa Beans Using Supercritical Carbon Dioxide and Ethanol. J. Agric. Food Chem. 2002, 50, 4820–4826. [Google Scholar] [CrossRef] [PubMed]
- Caudle, A.G.; Gu, Y.; Bell, L.N. Improved analysis of theobromine and caffeine in chocolate food products formulated with cocoa powder. Food Res. Int. 2001, 34, 599–603. [Google Scholar] [CrossRef]
- Hulbert, G.J.; Biswal, R.N.; Mehr, C.B.; Walker, T.H.; Collins, J.L. Solid/liquid extraction of caffeine from guaraná with methylene chloride/Extracción solido-liquido de cafeina de guarana con cloruro de metileno. Food Sci. Technol. Int. 1998, 4, 53–58. [Google Scholar] [CrossRef]
- Belščak, A.; Komes, D.; Horžić, D.; Ganić, K.K.; Karlović, D. Comparative study of commercially available cocoa products in terms of their bioactive composition. Food Res. Int. 2009, 42, 707–716. [Google Scholar] [CrossRef]
- Xia, Z.; Ni, Y.; Kokot, S. Simultaneous determination of caffeine, theophylline and theobromine in food samples by a kinetic spectrophotometric method. Food Chem. 2013, 141, 4087–4093. [Google Scholar] [CrossRef] [PubMed]
- De Aragão, N.M.; Veloso, M.C.C.; Bispo, M.S.; Ferreira, S.L.C.; de Andrade, J.B. Multivariate optimisation of the experimental conditions for determination of three methylxanthines by reversed-phase high-performance liquid chromatography. Talanta 2005, 67, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Zacharis, C.K.; Kika, F.S.; Tzanavaras, P.D.; Fytianos, K. Development and validation of a rapid ultra high pressure liquid chromatographic method for the determination of methylxanthines in herbal infusions. J. Chromatogr. B 2013, 927, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Takahashi, K.; Saijo, M.; Ishii, T.; Nagata, T. Rapid determination of theophylline, theobromine and caffeine in dietary supplements containing guarana by ultra-performance liquid chromatography. Shokuhin Eiseigaku Zasshi 2009, 50, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Kumazawa, T.; Seno, H.; Lee, X.-P.; Ishii, A.; Watanabe-Suzuki, K.; Sato, K.; Suzuki, O. Extraction of methylxanthines from human body fluids by solid-phase microextraction. Anal. Chim. Acta 1999, 387, 53–60. [Google Scholar] [CrossRef]
- Shrivas, K.; Wu, H.-F. Rapid determination of caffeine in one drop of beverages and foods using drop-to-drop solvent microextraction with gas chromatography/mass spectrometry. J. Chromatogr. A 2007, 1170, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Tserng, K.Y. Gas chromatographic-mass spectrometric quantitation of theophylline and its metabolites in biological fluids. J. Pharm. Sci. 1983, 72, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Khanchi, A.R.; Mahani, M.K.; Hajihosseini, M.; Maragheh, M.G.; Chaloosi, M.; Bani, F. Simultaneous spectrophotometric determination of caffeine and theobromine in Iranian tea by artificial neural networks and its comparison with PLS. Food Chem. 2007, 103, 1062–1068. [Google Scholar] [CrossRef]
- Khoshayand, M.R.; Abdollahi, H.; Shariatpanahi, M.; Saadatfard, A.; Mohammadi, A. Simultaneous spectrophotometric determination of paracetamol, ibuprofen and caffeine in pharmaceuticals by chemometric methods. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2008, 70, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Sahu, A. Spectrophotometric determination of caffeine and theophylline in pure alkaloids and its application in pharmaceutical formulations. Anal. Biochem. 2006, 349, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Armenta, S.; Garrigues, S.; de la Guardia, M. Solid-phase FT-Raman determination of caffeine in energy drinks. Anal. Chim. Acta 2005, 547, 197–203. [Google Scholar] [CrossRef]
- Moreira, A.B.; Dias, I.L.T.; Neto, G.O.; Zagatto, E.A.G.; Ferreira, M.M.C.; Kubota, L.T. Solid-phase spectrofluorimetric determination of acetylsalicylic acid and caffeine in pharmaceutical preparations using partial least-squares multivariate calibration. Talanta 2005, 67, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.C.L.; Poppi, R.J. Simultaneous determination of acetylsalicylic acid, paracetamol and caffeine using solid-phase molecular fluorescence and parallel factor analysis. Anal. Chim. Acta 2009, 642, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, M.M.; Irudayaraj, J. Rapid determination of caffeine content in soft drinks using FTIR-ATR spectroscopy. Food Chem. 2002, 78, 261–266. [Google Scholar] [CrossRef]
- Meinhart, A.D.; Bizzotto, C.S.; Ballus, C.A.; Prado, M.A.; Bruns, R.E.; Filho, J.T.; Godoy, H.T. Optimisation of a CE method for caffeine analysis in decaffeinated coffee. Food Chem. 2010, 120, 1155–1161. [Google Scholar] [CrossRef]
- Dilek Uysal, U.; Aturki, Z.; Raggi, M.A.; Fanali, S. Separation of catechins and methylxanthines in tea samples by capillary electrochromatography. J. Sep. Sci. 2009, 32, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Horie, H.; Mukai, T.; Kohata, K. Simultaneous determination of qualitatively important components in green tea infusions using capillary electrophoresis. J. Chromatogr. A 1997, 758, 332–335. [Google Scholar] [CrossRef]
- Thormann, W.; Minger, A.; Molteni, S.; Caslavska, J.; Gebauer, P. Determination of substituted purines in body fluids by micellar electrokinetic capillary chromatography with direct sample injection. J. Chromatogr. 1992, 593, 275–288. [Google Scholar] [CrossRef]
- Zhao, Y.; Lunte, C.E. Determination of caffeine and its metabolites by micellar electrokinetic capillary electrophoresis. J. Chromatogr. B Biomed. Sci. Appl. 1997, 688, 265–274. [Google Scholar] [CrossRef]
- Bonoli, M.; Colabufalo, P.; Pelillo, M.; Gallina Toschi, T.; Lercker, G. Fast determination of catechins and xanthines in tea beverages by micellar electrokinetic chromatography. J. Agric. Food Chem. 2003, 51, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Sanghavi, B.J.; Srivastava, A.K. Simultaneous voltammetric determination of acetaminophen, aspirin and caffeine using an in situ surfactant-modified multiwalled carbon nanotube paste electrode. Electrochim. Acta 2010, 55, 8638–8648. [Google Scholar] [CrossRef]
- Yang, S.; Yang, R.; Li, G.; Qu, L.; Li, J.; Yu, L. Nafion/multi-wall carbon nanotubes composite film coated glassy carbon electrode for sensitive determination of caffeine. J. Electroanal. Chem. 2010, 639, 77–82. [Google Scholar] [CrossRef]
- Cook, C.E.; Tallent, C.R.; Amerson, E.W.; Myers, M.W.; Kepler, J.A.; Taylor, G.F.; Christensen, H.D. Caffeine in plasma and saliva by a radioimmunoassay procedure. J. Pharmacol. Exp. Ther. 1976, 199, 679–686. [Google Scholar] [PubMed]
- Van Aerde, P.; Moerman, E.; Van Severen, R.; Braeckman, P. Determination of plasma theophylline by straight-phase high-performance liquid chromatography: Elimination of interfering caffeine metabolites. J. Chromatogr. 1981, 222, 467–474. [Google Scholar] [CrossRef]
- Aranda, M.; Morlock, G. Simultaneous determination of caffeine, ergotamine, and metamizol in solid pharmaceutical formulation by HPTLC-UV-FLD with mass confirmation by online HPTLC-ESI-MS. J. Chromatogr. Sci. 2007, 45, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Ford, M.J.; Deibel, M.A.; Tomkins, B.A.; van Berkel, G.J. Quantitative thin-layer chromatography/mass spectrometry analysis of caffeine using a surface sampling probe electrospray ionization tandem mass spectrometry system. Anal. Chem. 2005, 77, 4385–4389. [Google Scholar] [CrossRef] [PubMed]
- Khasanov, V.V.; Dychko, K.A.; Kuryaeva, T.T.; Ryzhova, G.L.; Mal’tseva, E.V. A new procedure for caffeine determination. Russ. J. Appl. Chem. 2005, 78, 1427–1429. [Google Scholar] [CrossRef]
- Llorent-Martínez, E.J.; García-Reyes, J.F.; Ortega-Barrales, P.; Molina-Díaz, A. Solid-phase ultraviolet sensing system for determination of methylxanthines. Anal. Bioanal. Chem. 2005, 382, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.F.; Vandemark, F.L.; Schmidt, G.J. More sensitive high-pressure liquid-chromatographic determiantion of theophylline in serum. Clin. Chem. 1976, 22, 1903–1906. [Google Scholar] [PubMed]
- Meyer, A.; Ngiruwonsanga, T.; Henze, G. Determination of adenine, caffeine, theophylline and theobromine by HPLC with amperometric detection. Anal. Bioanal. Chem. 1996, 356, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Chen, B.; Ma, M.; Luo, X.; Zhang, F.; Yao, S.; Wan, Z.; Yang, D.; Hang, H. Simultaneous analysis of theanine, chlorogenic acid, purine alkaloids and catechins in tea samples with the help of multi-dimension information of on-line high performance liquid chromatography/electrospray-mass spectrometry. J. Pharm. Biomed. Anal. 2004, 34, 695–704. [Google Scholar] [CrossRef]
- Marchei, E.; Pellegrini, M.; Pacifici, R.; Palmi, I.; Pichini, S. Development and validation of a high-performance liquid chromatography-mass spectrometry assay for methylxanthines and taurine in dietary supplements. J. Pharm. Biomed. Anal. 2005, 37, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Ptolemy, A.S.; Tzioumis, E.; Thomke, A.; Rifai, S.; Kellogg, M. Quantification of theobromine and caffeine in saliva, plasma and urine via liquid chromatography-tandem mass spectrometry: A single analytical protocol applicable to cocoa intervention studies. J. Chromatogr. B 2010, 878, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Huck, C.W.; Guggenbichler, W.; Bonn, G.K. Analysis of caffeine, theobromine and theophylline in coffee by near infrared spectroscopy (NIRS) compared to high-performance liquid chromatography (HPLC) coupled to mass spectrometry. Anal. Chim. Acta 2005, 538, 195–203. [Google Scholar] [CrossRef]
- Bech, B.H.; Bossi, R. Simultaneous determination of methylxanthines and cotinine in human plasma by solid-phase extraction followed by LC-MS-MS. Spectroscopy 2015, 27, 31–34. [Google Scholar]
- Rail, T.W. Central nervous system stimulants, the xanthines. In In The Pharmacologic Basis of Therapeutics, 6th ed.; Gilman, L.G.A., Ed.; Macmillan: Toronto, ON, Canada, 1980; p. 592. [Google Scholar]
- Moffat, A. Clarke’s ‘Isolation and Identification of Drugs’ (2nd edition) Senior consulting editor: A.C. Moffat Reviewed by I A Blair. J. Pharm. Pharmacol. 1986, 38, 942–944. [Google Scholar]
- Tarka, S.M.; Cornish, H.H. The toxicology of cocoa and methylxanthines: A review of the literature. CRC Crit. Rev. Toxicol. 1982, 9, 275–312. [Google Scholar] [CrossRef]
- Testa, B. Purine receptors and heir pharmacological roles. In Advances in Drug Research; Jovanovic, H.B., Ed.; Academic Press: London, UK, 1989; Volume 18, pp. 292–430. [Google Scholar]
- Mueller, C.E.; Shi, D.; Manning, M.; Daly, J.W. Synthesis of paraxanthine analogs (1,7-disubstituted xanthines) and other xanthines unsubstituted at the 3-position: Structure-activity relationships at adenosine receptors. J. Med. Chem. 1993, 36, 3341–3349. [Google Scholar] [CrossRef]
- Miyamoto, K.-I.; Sakai, R.; Yamamoto, Y.; Konno, K.; Sanae, F.; Hasegawa, T.; Takagi, K. Selective Bronchodilators from 1-(5′-Oxohexyl)xanthines. J. Pharm. Pharmacol. 1992, 44, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Hasegawa, T.; Kuzuya, T.; Ogawa, K.; Watanabe, T.; Satake, T.; Miyamoto, K.; Wakusawa, S.; Koshiura, R. Structure-activity relationship in N3-alkyl-xanthine derivatives. Jpn. J. Pharmacol. 1988, 46, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.; Müller, C.E.; Daly, J.W.; Beaven, M.A. Methylxanthines block antigen-induced responses in RBL-2H3 cells independently of adenosine receptors or cyclic AMP: Evidence for inhibition of antigen binding to IgE. J. Pharmacol. Exp. Ther. 1991, 258, 954–962. [Google Scholar] [PubMed]
- Bruns, R.F. Adenosine antagonism by purines, pteridines and benzopteridines in human fibroblasts. Biochem. Pharmacol. 1981, 30, 325–333. [Google Scholar] [CrossRef]
- Bruns, R.F.; Daly, J.W.; Snyder, S.H. Adenosine receptor binding: Structure-activity analysis generates extremely potent xanthine antagonists. Proc. Natl. Acad. Sci. USA 1983, 80, 2077–2080. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.W.; Padgett, W.; Shamim, M.T.; Butts-Lamb, P.; Waters, J. 1,3-Dialkyl-8-(p-sulfophenyl)xanthines: Potent water-soluble antagonists for A1- and A2-adenosine receptors. J. Med. Chem. 1985, 28, 487–492. [Google Scholar] [CrossRef] [PubMed]
- León-Carmona, J.R.; Galano, A. Is caffeine a good scavenger of oxygenated free radicals? J. Phys. Chem. B 2011, 115, 4538–4546. [Google Scholar] [CrossRef] [PubMed]
- Grucka-Mamczar, E.; Zalejska-Fiolka, J.; Chlubek, D.; Kasperczyk, S.; Blaszczyk, U.; Kasperczyk, A.; Swietochowska, E.; Birkner, E. The influence of sodium fluoride and caffeine on the activity of antioxidative enzymes and the concentration of malondialdehyde in rat liver. Fluoride 2009, 42, 105–109. [Google Scholar]
- Ofluoglu, E.; Pasaoglu, H.; Pasaoglu, A. The effects of caffeine on l-arginine metabolism in the brain of rats. Neurochem. Res. 2008, 34, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Azam, S.; Hadi, N.; Khan, N.U.; Hadi, S.M. Antioxidant and prooxidant properties of caffeine, theobromine and xanthine. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2003, 9, BR325–BR330. [Google Scholar]
- Choi, O.H.; Shamim, M.T.; Padgett, W.L.; Daly, J.W. Caffeine and theophylline analogues: Correlation of behavioral effects with activity as adenosine receptor antagonists and as phosphodiesterase inhibitors. Life Sci. 1988, 43, 387–398. [Google Scholar] [CrossRef]
- McPherson, P.S.; Kim, Y.-K.; Valdivia, H.; Knudson, C.M.; Takekura, H.; Franzini-Armstrong, C.; Coronadot, R.; Campbell, K.P. The brain ryanodine receptor: A caffeine-sensitive calcium release channel. Neuron 1991, 7, 17–25. [Google Scholar] [CrossRef]
- Chen, J.-F.; Chern, Y. Impacts of methylxanthines and adenosine receptors on neurodegeneration: Human and experimental studies. In Methylxanthines; Springer: Berlin/Heidelberg, Germany, 2011; pp. 267–310. [Google Scholar]
- Tazzeo, T.; Bates, G.; Roman, H.N.; Lauzon, A.-M.; Khasnis, M.D.; Eto, M.; Janssen, L.J. Caffeine relaxes smooth muscle through actin depolymerization. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L334–L342. [Google Scholar] [CrossRef] [PubMed]
- Dent, G.; Giembycz, M.A.; Rabe, K.F.; Wolf, B.; Barnes, P.J.; Magnussen, H. Theophylline suppresses human alveolar macrophage respiratory burst through phosphodiesterase inhibition. Am. J. Respir. Cell Mol. Biol. 1994, 10, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Meissner, G. Structure-activity relationship of xanthines and skeletal muscle ryanodine receptor/Ca2+ release channel. Pharmacology 1997, 54, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Nawrot, P.; Jordan, S.; Eastwood, J.; Rotstein, J.; Hugenholtz, A.; Feeley, M. Effects of caffeine on human health. Food Addit. Contam. 2003, 20, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; IJzerman, A.P.; Jacobson, K.A.; Klotz, K.-N.; Linden, J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 2001, 53, 527–552. [Google Scholar] [PubMed]
- Olah, M.E.; Stiles, G.L. Adenosine receptor subtypes: Characterization and therapeutic regulation. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 581–606. [Google Scholar] [CrossRef] [PubMed]
- Green, R.D.; Stanberry, L.R. Elevation of cyclic amp in C-1300 murine neuroblastoma by adenosine and related compounds and the antagonism of this response by methylxanthines. Biochem. Pharmacol. 1977, 26, 37–43. [Google Scholar] [CrossRef]
- Boulenger, J.P.; Patel, J.; Marangos, P.J. Effects of caffeine and theophylline on adenosine and benzodiazepine receptors in human brain. Neurosci. Lett. 1982, 30, 161–166. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Irenius, E.; Kull, B.; Schulte, G. Comparison of the potency of adenosine as an agonist at human adenosine receptors expressed in Chinese hamster ovary cells. Biochem. Pharmacol. 2001, 61, 443–448. [Google Scholar] [CrossRef]
- Klotz, K.-N.; Hessling, J.; Hegler, J.; Owman, C.; Kull, B.; Fredholm, B.B.; Lohse, J.M. Comparative pharmacology of human adenosine receptor subtypes—Characterization of stably transfected receptors in CHO cells. Naunyn Schmiedeberg’s Arch. Pharmacol. 1997, 357, 1–9. [Google Scholar] [CrossRef]
- Daly, J.W.; Butts-Lamb, P.; Padgett, W. Subclasses of adenosine receptors in the central nervous system: Interaction with caffeine and related methylxanthines. Cell. Mol. Neurobiol. 1983, 3, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Baraldi, P.G.; Cacciari, B.; Romagnoli, R.; Merighi, S.; Varani, K.; Borea, P.A.; Spalluto, G. A3 adenosine receptor ligands: History and perspectives. Med. Res. Rev. 2000, 20, 103–128. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Persson, C.G.A. Xanthine derivatives as adenosine receptor antagonists. Eur. J. Pharmacol. 1982, 81, 673–676. [Google Scholar] [CrossRef]
- Franco, R.; Oñatibia-Astibia, A.; Martínez-Pinilla, E. Health benefits of methylxanthines in cacao and chocolate. Nutrients 2013, 5, 4159–4173. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Daly, J.W. Chronic effects of xanthines on levels of central receptors in mice. Cell. Mol. Neurobiol. 1999, 19, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Carney, J.M.; Holloway, F.A.; Modrow, H.E. Discriminative stimulus properties of methylxanthines and their metabolites in rats. Life Sci. 1985, 36, 913–920. [Google Scholar] [CrossRef]
- Schwabe, U.; Ukena, D.; Lohse, M.J. Xanthine derivatives as antagonists at A1 and A2 adenosine receptors. Naunyn Schmiedeberg Arch. Pharmacol. 1985, 330, 212–221. [Google Scholar] [CrossRef]
- Moro, S.; Gao, Z.-G.; Jacobson, K.A.; Spalluto, G. Progress in the pursuit of therapeutic adenosine receptor antagonists. Med. Res. Rev. 2006, 26, 131–159. [Google Scholar] [CrossRef] [PubMed]
- Baraldi, P.G.; Fruttarolo, F.; Tabrizi, M.A.; Romagnoli, R.; Preti, D. Novel 8-heterocyclyl xanthine derivatives in drug development—An update. Expert Opin. Drug Discov. 2007, 2, 1161–1183. [Google Scholar] [CrossRef] [PubMed]
- Van der Walt, M.M.; Terre’Blanche, G. 1,3,7-Triethyl-substituted xanthines—Possess nanomolar affinity for the adenosine A1 receptor. Bioorg. Med. Chem. 2015, 23, 6641–6649. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.A.; von Lubitz, D.K.J.E.; Daly, J.W.; Fredholm, B.B. Adenosine receptor ligands: Differences with acute versus chronic treatment. Trends Pharmacol. Sci. 1996, 17, 108–113. [Google Scholar] [CrossRef]
- Varani, K.; Portaluppi, F.; Merighi, S.; Ongini, E.; Belardinelli, L.; Borea, P.A. Caffeine alters A2A adenosine receptors and their function in human platelets. Circulation 1999, 99, 2499–2502. [Google Scholar] [CrossRef] [PubMed]
- Biaggioni, I.; Paul, S.; Puckett, A.; Arzubiaga, C. Caffeine and theophylline as adenosine receptor antagonists in humans. J. Pharmacol. Exp. Ther. 1991, 258, 588–593. [Google Scholar] [PubMed]
- Herman, A.; Herman, A.P. Caffeine’s mechanisms of action and its cosmetic use. Skin Pharmacol. Physiol. 2013, 26, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Beavo, J.A.; Rogers, N.L.; Crofford, O.B.; Hardman, J.G.; Sutherland, E.W.; Newman, E.V. Effects of xanthine derivatives on lipolysis and on adenosine 3′,5′-monophosphate phosphodiesterase activity. Mol. Pharmacol. 1970, 6, 597–603. [Google Scholar] [PubMed]
- Sugimoto, N.; Miwa, S.; Hitomi, Y.; Nakamura, H.; Tsuchiya, H.; Yachie, A. Theobromine, the primary methylxanthine found in Theobroma cacao, prevents malignant glioblastoma proliferation by negatively regulating phosphodiesterase-4, extracellular signal-regulated kinase, Akt/mammalian target of rapamycin kinase, and nuclear factor-κB. Nutr. Cancer 2014, 66, 419–423. [Google Scholar] [PubMed]
- Ruangkittisakul, A.; Ballanyi, K. Methylxanthine reversal of opioid-evoked inspiratory depression via phosphodiesterase-4 blockade. Respir. Physiol. Neurobiol. 2010, 172, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Sassone-Corsi, P. The cyclic AMP pathway. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.H.; Sekhar, K.R.; Ke, H.; Corbin, J.D. Inhibition of cyclic nucleotide phosphodiesterases by methylxanthines and related compounds. In Methylxanthines; Springer: Berlin/Heidelberg, Germany, 2011; pp. 93–133. [Google Scholar]
- Daly, J.W. Caffeine analogs: Biomedical impact. Cell. Mol. Life Sci. 2007, 64, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Rabe, K.; Magnussen, H.; Dent, G. Theophylline and selective PDE inhibitors as bronchodilators and smooth muscle relaxants. Eur. Respir. J. 1995, 8, 637–642. [Google Scholar] [PubMed]
- Pissarnitski, D.A.; Asberom, T.; Boyle, C.D.; Chackalamannil, S.; Chintala, M.; Clader, J.W.; Greenlee, W.J.; Hu, Y.; Kurowski, S.; Myers, J.; et al. SAR development of polycyclic guanine derivatives targeted to the discovery of a selective PDE5 inhibitor for treatment of erectile dysfunction. Bioorg. Med. Chem. Lett. 2004, 14, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Arnold, R.; Beer, D.; Bhalay, G.; Baettig, U.; Collingwood, S.P.; Craig, S.; Devereux, N.; Dunstan, A.; Glen, A.; Gomez, S.; et al. 8-Aryl xanthines potent inhibitors of phosphodiesterase 5. Bioorg. Med. Chem. Lett. 2002, 12, 2587–2590. [Google Scholar] [CrossRef]
- Huai, Q.; Liu, Y.; Francis, S.H.; Corbin, J.D.; Ke, H. Crystal structures of phosphodiesterases 4 and 5 in complex with inhibitor 3-isobutyl-1-methylxanthine suggest a conformation determinant of inhibitor selectivity. J. Biol. Chem. 2004, 279, 13095–13101. [Google Scholar] [CrossRef] [PubMed]
- Scapin, G.; Patel, S.B.; Chung, C.; Varnerin, J.P.; Edmondson, S.D.; Mastracchio, A.; Parmee, E.R.; Singh, S.B.; Becker, J.W.; van der Ploeg, L.H.T.; et al. Crystal structure of human phosphodiesterase 3B: Atomic basis for substrate and inhibitor specificity. Biochemistry 2004, 43, 6091–6100. [Google Scholar] [CrossRef] [PubMed]
- Lopez, F.; Miller, L.G.; Greenblatt, D.J.; Kaplan, G.B.; Shader, R.I. Interaction of caffeine with the GABAA receptor complex: Alterations in receptor function but not ligand binding. Eur. J. Pharmacol. Mol. Pharmacol. 1989, 172, 453–459. [Google Scholar] [CrossRef]
- Sugimoto, T.; Sugimoto, M.; Uchida, I.; Mashimo, T.; Okada, S. Inhibitory effect of theophylline on recombinant GABAA receptor. Neuro Rep. 2001, 12, 489–493. [Google Scholar] [CrossRef]
- Shi, D.; Padgett, W.L.; Daly, J.W. Caffeine analogs: Effects on ryanodine-sensitive calcium-release channels and GABAA receptors. Cell. Mol. Neurobiol. 2003, 23, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Roca, D.J.; Schiller, G.D.; Farb, D.H. Chronic caffeine or theophylline exposure reduces γ-aminobutyric acid/benzodiazepine receptor site interactions. Mol. Pharmacol. 1988, 33, 481–485. [Google Scholar] [PubMed]
- Daly, J.W. Alkylxanthines as research tools. J. Auton. Nerv. Syst. 2000, 81, 44–52. [Google Scholar] [CrossRef]
- Gaburjakova, J.; Gaburjakova, M. Coupled gating modifies the regulation of cardiac ryanodine receptors by luminal Ca2+. Biochim. Biophys. Acta BBA Biomembr. 2014, 1838, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.E.; Daly, J.W. Stimulation of calcium release by caffeine analogs in pheochromocytoma cells. Biochem. Pharmacol. 1993, 46, 1825–1829. [Google Scholar] [CrossRef]
- Rousseau, E.; Ladine, J.; Liu, Q.-Y.; Meissner, G. Activation of the Ca2+ release channel of skeletal muscle sarcoplasmic reticulum by caffeine and related compounds. Arch. Biochem. Biophys. 1988, 267, 75–86. [Google Scholar] [CrossRef]
- Shou, Q.; Pan, S.; Tu, J.; Jiang, J.; Ling, Y.; Cai, Y.; Chen, M.; Wang, D. Modulation effect of Smilax glabra flavonoids on ryanodine receptor mediated intracellular Ca2+ release in cardiomyoblast cells. J. Ethnopharmacol. 2013, 150, 389–392. [Google Scholar] [CrossRef] [PubMed]
- Kuemmerle, J.F.; Murthy, K.S.; Makhlouf, G.M. Agonist-activated, ryanodine-sensitive, IP3-insensitive Ca2+ release channels in longitudinal muscle of intestine. Am. J. Physiol. Cell Physiol. 1994, 266, C1421–C1431. [Google Scholar]
- Cavallaro, R.A.; Filocamo, L.; Galuppi, A.; Galione, A.; Brufani, M.; Genazzani, A.A. Potentiation of cADPR-induced Ca2+-release by methylxanthine analogues. J. Med. Chem. 1999, 42, 2527–2534. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B. On the mechanism of action of theophylline and caffeine. Acta Medica Scand. 1985, 217, 149–153. [Google Scholar] [CrossRef]
- Marangos, P.J.; Paul, S.M.; Parma, A.M.; Goodwin, F.K.; Syapin, P.; Skolnick, P. Purinergic inhibition of diazepam binding to rat brain (in vitro). Life Sci. 1979, 24, 851–857. [Google Scholar] [CrossRef]
- Sattin, A.; Rall, T.W. The effect of adenosine and adenine nucleotides on the cyclic adenosine 3′,5′-phosphate content of guinea pig cerebral cortex slices. Mol. Pharmacol. 1970, 6, 13–23. [Google Scholar] [PubMed]
- Cardinali, D.P. Methylxanthines: Possible mechanisms of action in brain. Trends Pharmacol. Sci. 1980, 1, 405–407. [Google Scholar] [CrossRef]
- Butcher, R.W.; Sutherland, E.W. Adenosine 3′,5′-Phosphate in Biological Materials: I. Purification and properties of cyclic 3′,5′-nucleotide phosphodiesterase and use of this enzyme to characterize adenosine 3′,5′-phosphate in human urine. J. Biol. Chem. 1962, 237, 1244–1250. [Google Scholar] [PubMed]
- Nicholson, C.D.; Jackman, S.A.; Wilke, R. The ability of denbufylline to inhibit cyclic nucleotide phosphodiesterase and its affinity for adenosine receptors and the adenosine re-uptake site. Br. J. Pharmacol. 1989, 97, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Geraets, L.; Moonen, H.J.J.; Wouters, E.F.M.; Bast, A.; Hageman, G.J. Caffeine metabolites are inhibitors of the nuclear enzyme poly(ADP-ribose)polymerase-1 at physiological concentrations. Biochem. Pharmacol. 2006, 72, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Rao, F.V.; Andersen, O.A.; Vora, K.A.; DeMartino, J.A.; van Aalten, D.M.F. Methylxanthine drugs are chitinase inhibitors: Investigation of inhibition and binding modes. Chem. Biol. 2005, 12, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Lim, S.; Caramori, G.; Cosio, B.; Chung, K.F.; Adcock, I.M.; Barnes, P.J. A molecular mechanism of action of theophylline: Induction of histone deacetylase activity to decrease inflammatory gene expression. Proc. Natl. Acad. Sci. USA 2002, 99, 8921–8926. [Google Scholar] [CrossRef] [PubMed]
- Cosio, B.G.; Tsaprouni, L.; Ito, K.; Jazrawi, E.; Adcock, I.M.; Barnes, P.J. Theophylline restores histone deacetylase activity and steroid responses in COPD macrophages. J. Exp. Med. 2004, 200, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M.; Dobes, P. Caffeine inhibits acetylcholinesterase, but not butyrylcholinesterase. Int. J. Mol. Sci. 2013, 14, 9873–9882. [Google Scholar] [CrossRef] [PubMed]
- Miroslav, P. The effects of caffeine on the cholinergic system. Mini Rev. Med. Chem. 2014, 14, 543–549. [Google Scholar]
- Petzer, A.; Pienaar, A.; Petzer, J.P. The interactions of caffeine with monoamine oxidase. Life Sci. 2013, 93, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Manfredi, G.; Sammartano, S. Quantitative study on the non-covalent interactions between ATP and caffeine, theophylline and theobromine in aqueous solution. Fluid Phase Equilibria 2011, 308, 47–54. [Google Scholar] [CrossRef]
- Johnson, I.M.; Prakash, H.; Prathiba, J.; Raghunathan, R.; Malathi, R. Spectral analysis of naturally occurring methylxanthines (theophylline, theobromine and caffeine) binding with DNA. PLoS ONE 2012, 7, e50019. [Google Scholar] [CrossRef] [PubMed]
- Johnson, I.M.; Kumar, S.G.B.; Malathi, R. RNA binding efficacy of theophylline, theobromine and caffeine. J. Biomol. Struct. Dyn. 2003, 20, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pinilla, E.; Oñatibia-Astibia, A.; Franco, R. The relevance of theobromine for the beneficial effects of cocoa consumption. Front. Pharmacol. 2015, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, M.J. Pharmacokinetics and metabolism of natural methylxanthines in animal and man. In Methylxanthines; Springer: Berlin/Heidelberg, Germany, 2011; pp. 33–91. [Google Scholar]
- Lelo, A.; Birkett, D.J.; Robson, R.A.; Miners, J.O. Comparative pharmacokinetics of caffeine and its primary demethylated metabolites paraxanthine, theobromine and theophylline in man. Br. J. Cl. Pharmacol. 1986, 22, 177–182. [Google Scholar] [CrossRef]
- Blanchard, J.; Sawers, S.J.A. The absolute bioavailability of caffeine in man. Eur. J. Clin. Pharmacol. 1983, 24, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Zandvliet, A.S.; Huitema, A.D.R.; de Jonge, M.E.; den Hoed, R.; Sparidans, R.W.; Hendriks, V.M.; van Den Brink, W.; van Ree, J.M.; Beijnen, J.H. Population pharmacokinetics of caffeine and its metabolites theobromine, paraxanthine and theophylline after inhalation in combination with diacetylmorphine. Basic Clin. Pharmacol. Toxicol. 2005, 96, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Teekachunhatean, S.; Tosri, N.; Rojanasthien, N.; Srichairatanakool, S.; Sangdee, C. Pharmacokinetics of caffeine following a single administration of coffee enema versus oral coffee consumption in healthy male subjects. ISRN Pharmacol. 2013, 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Perera, V.; Gross, A.S.; McLachlan, A.J. Caffeine and paraxanthine HPLC assay for CYP1A2 phenotype assessment using saliva and plasma. Biomed. Chromatogr. 2010, 24, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.B.; Simons, K.J.; Gillespie, C.A.; Simons, F.E.R. The bronchodilator effects and pharmacokinetics of caffeine in asthma. N. Engl. J. Med. 1984, 310, 743–746. [Google Scholar] [CrossRef] [PubMed]
- French, I.W.; Mildon, C.A. The pharmacokinetics of theophylline. Curr. Med. Res. Opin. 1979, 6 (Suppl. S6), 3–13. [Google Scholar] [CrossRef]
- Resman, B.H.; Blumenthal, H.P.; Jusko, W.J. Breast milk distribution of theobromine from chocolate. J. Pediatr. 1977, 91, 477–480. [Google Scholar] [CrossRef]
- Mumford, K.G.; Benowitz, L.N.; Evans, M.S.; Kaminski, J.B.; Preston, L.K.; Sannerud, A.C.; Silverman, K.; Griffiths, R.R. Absorption rate of methylxanthines following capsules, cola and chocolate. Eur. J. Clin. Pharmacol. 1996, 51, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Laska, E.M.; Sunshine, A.; Mueller, F.; Elvers, W.B.; Siegel, C.; Rubin, A. Caffeine as an analgesic adjuvant. JAMA 1984, 251, 1711–1718. [Google Scholar] [CrossRef] [PubMed]
- Derry, C.J.; Derry, S.; Moore, R.A. Caffeine as an analgesic adjuvant for acute pain in adults. Cochrane Database Syst. Rev. 2014, 12, 1–62. [Google Scholar]
- Lee, I.-A.; Kamba, A.; Low, D.; Mizoguchi, E. Novel methylxanthine derivative-mediated anti-inflammatory effects in inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Batéjat, D.; Coste, O.; Doireau, P.; Chauffard, F.; Enslen, M.; Lagarde, D.; Pierard, C. Recovery after prolonged sleep deprivation: Residual effects of slow-release caffeine on recovery sleep, sleepiness and cognitive functions. Neuropsychobiology 2005, 51, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Lozano, R.P.; García, Y.A.; Tafalla, D.B.; Albaladejo, M.F. Cafeína: Un nutriente, un fármaco, o una droga de abuso. Adicciones 2007, 19, 225–238. [Google Scholar] [CrossRef]
- Smit, H.J.; Gaffan, E.A.; Rogers, P.J. Methylxanthines are the psycho-pharmacologically active constituents of chocolate. Psychopharmacology 2004, 176, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.N.; Pamplona, F.A.; Prediger, R.D. Adenosine receptor antagonists for cognitive dysfunction: A review of animal studies. Front. Biosci. 2008, 13, 2614–2632. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, M.E.M.; Cesário, C.; Hiroi, R.H.; Rosalen, P.L.; Cunha, C.D. Effects of caffeine on learning and memory in rats tested in the Morris water maze. Braz. J. Med. Biol. Res. 2002, 35, 1201–1208. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R. The effects of ginseng, ephedrine, and caffeine on cognitive performance, mood and energy. Nutr. Rev. 2001, 59, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Haskell, C.F.; Kennedy, D.O.; Wesnes, K.A.; Scholey, A.B. Cognitive and mood improvements of caffeine in habitual consumers and habitual non-consumers of caffeine. Psychopharmacology 2005, 179, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Riedel, W.; Hogervorst, E.; Leboux, R.; Verhey, F.; van Praag, H.; Jolles, J. Caffeine attenuates scopolamine-induced memory impairment in humans. Psychopharmacology 1995, 122, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Maia, L.; de Mendonça, A. Does caffeine intake protect from Alzheimer’s disease? Eur. J. Neurol. 2002, 9, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Cao, C. Caffeine and coffee as therapeutics against Alzheimer’s disease. J. Alzheimers Dis. 2010, 20, S117–S126. [Google Scholar] [PubMed]
- Eskelinen, M.H.; Ngandu, T.; Tuomilehto, J.; Soininen, H.; Kivipelto, M. Midlife coffee and tea drinking and the risk of latelife dementia: A population-based CAIDE study. J. Alzheimers Dis. 2009, 16, 85–91. [Google Scholar] [PubMed]
- Chen, J.F.; Xu, K.; Petzer, J.P.; Staal, R.; Xu, Y.H.; Beilstein, M.; Sonsalla, P.K.; Castagnoli, K.; Castagnoli, N.J.; Schwarzschild, M.A. Neuroprotection by caffeine and A2A adenosine receptor inactivation in a model of Parkinson’s disease. J. Neurosci. 2001, 21, RC143. [Google Scholar] [PubMed]
- Costa, J.; Lunet, N.; Santos, C.; Santos, J.; Vaz-Carneiro, A. Caffeine exposure and the risk of Parkinson’s disease: A systematic review and meta-analysis of observational studies. J. Alzheimers Dis. 2010, 20, S221–S238. [Google Scholar] [PubMed]
- Postuma, R.B.; Lang, A.E.; Munhoz, R.P.; Charland, K.; Pelletier, A.; Moscovich, M.; Filla, L.; Zanatta, D.; Rios Romenets, S.; Altman, R.; et al. Caffeine for treatment of Parkinson disease: A randomized controlled trial. Neurology 2012, 79, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Doré, A.S.; Robertson, N.; Errey, J.C.; Ng, I.; Hollenstein, K.; Tehan, B.; Hurrell, E.; Bennett, K.; Congreve, M.; Magnani, F.; et al. Structure of the adenosine A2A receptor in complex with ZM241385 and the xanthines XAC and caffeine. Structure 2011, 19, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gawryluk, J.W.; Wagener, J.F.; Ghribi, O.; Geiger, J.D. Caffeine blocks disruption of blood brain barrier in a rabbit model of Alzheimer’s disease. J. Neuroinflammation 2008, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lan, X.; Roche, I.; Liu, R.; Geiger, J.D. Caffeine protects against MPTP-induced blood-brain barrier dysfunction in mouse striatum. J. Neurochem. 2008, 107, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Schleif, W.; Rezai-Zadeh, K.; Jackson, E.K.; Zacharia, L.C.; Cracchiolo, J.R.; Shippy, D.; Tan, J. Caffeine protects Alzheimer’s mice against cognitive impairment and reduces brain β-amyloid production. Neuroscience 2006, 142, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Cirrito, J.R.; Lin, X.; Wang, L.; Verges, D.K.; Dickson, A.; Mamcarz, M.; Zhang, C.; Mori, T.; Arendash, G.W.; et al. Caffeine suppresses β-amyloid levels in plasma and brain of Alzheimer’s transgenic mice. J. Alzheimer’s Dis. JAD 2009, 17, 681–697. [Google Scholar] [PubMed]
- Chu, Y.-F.; Chang, W.-H.; Black, R.M.; Liu, J.-R.; Sompol, P.; Chen, Y.; Wei, H.; Zhao, Q.; Cheng, I.H. Crude caffeine reduces memory impairment and amyloid β1-42 levels in an Alzheimer’s mouse model. Food Chem. 2012, 135, 2095–2102. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Mori, T.; Cao, C.; Mamcarz, M.; Runfeldt, M.; Dickson, A.; Rezai-Zadeh, K.; Tane, J.; Citron, B.A.; et al. Caffeine reverses cognitive impairment and decreases brain amyloid-beta levels in aged Alzheimer’s disease mice. J. Alzheimers Dis. 2009, 17, 661–680. [Google Scholar] [PubMed]
- Lindsay, J.; Laurin, D.; Verreault, R.; Hébert, R.; Helliwell, B.; Hill, G.B.; McDowell, I. Risk factors for Alzheimer’s disease: A prospective analysis from the Canadian Study of Health and Aging. Am. J. Epidemiol. 2002, 156, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, K.; Carrière, I.; de Mendonça, A.; Portet, F.; Dartigues, J.F.; Rouaud, O.; Barberger-Gateau, P.; Ancelin, M.L. The neuroprotective effects of caffeine: A prospective population study (the Three City Study). Neurology 2007, 69, 536–545. [Google Scholar] [CrossRef] [PubMed]
- Johnson-Kozlow, M.; Kritz-Silverstein, D.; Barrett-Connor, E.; Morton, D. Coffee consumption and cognitive function among older adults. Am. J. Epidemiol. 2002, 156, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Van Gelder, B.M.; Buijsse, B.; Tijhuis, M.; Kalmijn, S.; Giampaoli, S.; Nissinen, A.; Kromhout, D. Coffee consumption is inversely associated with cognitive decline in elderly European men: The FINE Study. Eur. J. Clin. Nutr. 2006, 61, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Dai, S.; An, J.; Li, P.; Chen, X.; Xiong, R.; Liu, P.; Wang, H.; Zhao, Y.; Zhu, M.; et al. Chronic but not acute treatment with caffeine attenuates traumatic brain injury in the mouse cortical impact model. Neuroscience 2008, 151, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Sonsalla, P.K.; Wong, L.-Y.; Harris, S.L.; Richardson, J.R.; Khobahy, I.; Li, W.; Gadad, B.S.; German, D.C. Delayed caffeine treatment prevents nigral dopamine neuron loss in a progressive rat model of Parkinson’s disease. Exp. Neurol. 2012, 234, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, M.D.; Bower, J.H.; Maraganore, D.M.; McDonnell, S.K.; Peterson, B.J.; Ahlskog, J.E.; Schaid, D.J.; Rocca, W.A. Smoking, alcohol, and coffee consumption preceding Parkinson’s disease: A case-control study. Neurology 2000, 55, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Ross, G.; Abbott, R.D.; Petrovitch, H.; Morens, D.M.; Grandinetti, A.; Tung, K.H.; Tanner, C.M.; Masaki, K.H.; Blanchette, P.L.; Curb, J.D.; et al. Association of coffee and caffeine intake with the risk of parkinson disease. JAMA 2000, 283, 2674–2679. [Google Scholar] [CrossRef] [PubMed]
- Saaksjarvi, K.; Knekt, P.; Rissanen, H.; Laaksonen, M.A.; Reunanen, A.; Mannisto, S. Prospective study of coffee consumption and risk of Parkinson’s disease. Eur. J. Clin. Nutr. 2007, 62, 908–915. [Google Scholar] [CrossRef] [PubMed]
- Ascherio, A.; Zhang, S.M.; Hernán, M.A.; Kawachi, I.; Colditz, G.A.; Speizer, F.E.; Willett, W.C. Prospective study of caffeine consumption and risk of Parkinson’s disease in men and women. Ann. Neurol. 2001, 50, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Palacios, N.; Gao, X.; McCullough, M.L.; Schwarzschild, M.A.; Shah, R.; Gapstur, S.; Ascherio, A. Caffeine and risk of Parkinson disease in a large cohort of men and women. Mov. Disord. 2012, 27, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Li, S. Dose-response meta-analysis on coffee, tea and caffeine consumption with risk of Parkinson’s disease. Geriatr. Gerontol. Int. 2014, 14, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, N.; Simões, A.T.; Cunha, R.A.; de Almeida, L.P. Caffeine and adenosine A2A receptor inactivation decrease striatal neuropathology in a lentiviral-based model of Machado-Joseph disease. Ann. Neurol. 2013, 73, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Simonin, C.; Duru, C.; Salleron, J.; Hincker, P.; Charles, P.; Delval, A.; Youssov, K.; Burnouf, S.; Azulay, J.-P.; Verny, C.; et al. Association between caffeine intake and age at onset in Huntington’s disease. Neurobiol. Dis. 2013, 58, 179–182. [Google Scholar] [CrossRef] [PubMed]
- Henderson-Smart, D.J.; De Paoli, A.G. Methylxanthine treatment for apnoea in preterm infants. Cochrane Database Syst. Rev. 2010, 8. [Google Scholar] [CrossRef]
- Comer, A.M.; Perry, C.M.; Figgitt, D.P. Caffeine citrate: A review of its use in apnoea of prematurity. Paediatric Drugs 2001, 3, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, J. Current options in the management of apnea of prematurity. Clin. Pediatr. 2000, 39, 327–336. [Google Scholar] [CrossRef]
- Aranda, J.V.; Beharry, K.; Valencia, G.B.; Natarajan, G.; Davis, J. Caffeine impact on neonatal morbidities. J. Matern. Fetal Neonatal Med. 2010, 23 (Suppl. S3), 20–23. [Google Scholar] [CrossRef] [PubMed]
- Schoen, K.; Yu, T.; Stockmann, C.; Spigarelli, M.G.; Sherwin, C.M.T. Use of methylxanthine therapies for the treatment and prevention of apnea of prematurity. Pediatr. Drugs 2014, 16, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Lagercrantz, H.; Yamamoto, Y.; Fredholm, B.B.; Prabhakar, N.R.; von Euler, C. Adenosine analogues depress ventilation in rabbit neonates. Theophylline stimulation of respiration via adenosine receptors? Pediatr. Res. 1984, 18, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Kawai, A.; Okada, Y.; Mückenhoff, K.; Scheid, P. Theophylline and hypoxic ventilatory response in the rat isolated brainstem-spinal cord. Respir. Physiol. 1995, 100, 25–32. [Google Scholar] [CrossRef]
- Wennergren, G.; Wennergren, M. Neonatal breathing control mediated via the central chemoreceptors. Acta Physiol. Scand. 1983, 119, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Herlenius, E.; Lagercrantz, H. Adenosinergic modulation of respiratory neurones in the neonatal rat brainstem in vitro. J. Physiol. 1999, 518, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Eldridge, F.L.; Millhorn, D.E.; Kiley, J.P. Antagonism by theophylline of respiratory inhibition induced by adenosine. J. Appl. Physiol. 1985, 59, 1428–1433. [Google Scholar] [PubMed]
- Hedner, T.; Hedner, J.; Bergman, B.; Mueller, R.A.; Jonason, J. Characterization of adenosine-induced respiratory depression in the preterm rabbit. Biol. Neonate 1985, 47, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Mosca, E.V.; Ciechanski, P.; Roy, A.; Scheibli, E.C.; Ballanyi, K.; Wilson, R.J.A. Methylxanthine reversal of opioid-induced respiratory depression in the neonatal rat: Mechanism and location of action. Respir. Physiol. Neurobiol. 2014, 200, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Theophylline in chronic obstructive pulmonary disease: New horizons. Proc. Am. Thorac. Soc. 2005, 2, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Fozard, J.R.; McCarthy, C. Adenosine receptor ligands as potential therapeutics in asthma. Curr. Opin. Investig. Drugs 2002, 3, 69–77. [Google Scholar] [PubMed]
- Russo, C.; Arcidiacono, G.; Polosa, R. Adenosine receptors: Promising targets for the development of novel therapeutics and diagnostics for asthma. Fundam. Clin. Pharmacol. 2006, 20, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Theophylline for COPD. Thorax 2006, 61, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Usmani, O.S.; Belvisi, M.G.; Patel, H.J.; Crispino, N.; Birrell, M.A.; Korbonits, M.; Korbonits, D.; Barnes, P.J. Theobromine inhibits sensory nerve activation and cough. FASEB J. 2004. [Google Scholar] [CrossRef] [PubMed]
- Smit, H.J. Theobromine and the pharmacology of cocoa. In Methylxanthines; Springer: Berlin, Germany; Heidelberg, Germany, 2011; pp. 201–234. [Google Scholar]
- Riksen, N.P.; Smits, P.; Rongen, G.A. The cardiovascular effects of methylxanthines. In Methylxanthines; Springer: Berlin, Germany; Heidelberg, Germany, 2011; pp. 413–437. [Google Scholar]
- Cameron, O.G.; Modell, J.G.; Hariharan, M. Caffeine and human cerebral blood flow: A positron emission tomography study. Life Sci. 1990, 47, 1141–1146. [Google Scholar] [CrossRef]
- Okuno, T.; Sugiyama, T.; Tominaga, M.; Kojima, S.; Ikeda, T. Effects of caffeine on microcirculation of the human ocular fundus. Jpn. J. Ophthalmol. 2002, 46, 170–176. [Google Scholar] [CrossRef]
- Batterman, R.G.; Grossman, A.J.; Dubinsky, J.; Mouratoff, G. Reevaluation of the usefullness of theobromine calcium gluconate for the management of congestive heart failure and anginal syndrome. Int. Rec. Med. Gen. Pract. Clin. 1959, 172, 318–323. [Google Scholar] [PubMed]
- Larsson, S.C.; Orsini, N. Coffee consumption and risk of stroke: A dose-response meta-analysis of prospective studies. Am. J. Epidemiol. 2011, 174, 993–1001. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Bhupathiraju, S.N.; Satija, A.; van Dam, R.M.; Hu, F.B. Long-term coffee consumption and risk of cardiovascular disease: A systematic review and a dose-response meta-analysis of prospective cohort studies. Circulation 2014, 129, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Bohn, S.K.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. Effects of tea and coffee on cardiovascular disease risk. Food Funct. 2012, 3, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Monagas, M.; Andres-Lacueva, C.; Casas, R.; Urpí-Sardà, M.; Lamuela-Raventós, R.M.; Estruch, R. Regular consumption of cocoa powder with milk increases HDL cholesterol and reduces oxidized LDL levels in subjects at high-risk of cardiovascular disease. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Kris-Etherton, P.M.; Derr, J.A.; Mustad, V.A.; Seligson, F.H.; Pearson, T.A. Effects of a milk chocolate bar per day substituted for a high-carbohydrate snack in young men on an NCEP/AHA Step 1 Diet. Am. J. Clin. Nutr. 1994, 60, 1037S–1042S. [Google Scholar]
- Mursu, J.; Voutilainen, S.; Nurmi, T.; Rissanen, T.H.; Virtanen, J.K.; Kaikkonen, J.; Nyyssönen, K.; Salonen, J.T. Dark chocolate consumption increases HDL cholesterol concentration and chocolate fatty acids may inhibit lipid peroxidation in healthy humans. Free Radic. Biol. Med. 2004, 37, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Baba, S.; Osakabe, N.; Kato, Y.; Natsume, M.; Yasuda, A.; Kido, T.; Fukuda, K.; Muto, Y.; Kondo, K. Continuous intake of polyphenolic compounds containing cocoa powder reduces LDL oxidative susceptibility and has beneficial effects on plasma HDL-cholesterol concentrations in humans. Am. J. Clin. Nutr. 2007, 85, 709–717. [Google Scholar] [PubMed]
- Mellor, D.D.; Sathyapalan, T.; Kilpatrick, E.S.; Beckett, S.; Atkin, S.L. High-cocoa polyphenol-rich chocolate improves HDL cholesterol in Type 2 diabetes patients. Diabet. Med. 2010, 27, 1318–1321. [Google Scholar] [CrossRef] [PubMed]
- Neufingerl, N.; Zebregs, Y.E.; Schuring, E.A.; Trautwein, E.A. Effect of cocoa and theobromine consumption on serum HDL-cholesterol concentrations: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Kobayashi-Hattori, K.; Horiuchi, Y.; Oishi, Y.; Arai, S.; Takita, T. Regulation of the body fat percentage in developmental-stage rats by methylxanthine derivatives in a high-fat diet. Biosci. Biotechnol. Biochem. 2006, 70, 1134–1139. [Google Scholar] [CrossRef] [PubMed]
- Heppel, L.A.; Porterfield, V.T.; Peake, E.G. The lipotropic activity of caffeine, theobromine and theophylline. Arch. Biochem. Biophys. 1947, 15, 439–443. [Google Scholar]
- Bray, G.A.; Mothon, S.; Cohen, A.S. Mobilization of fatty acids in genetically obese rats. J. Lipid Res. 1970, 11, 517–521. [Google Scholar] [PubMed]
- Kuo, J.F.; de Renzo, E.C. A comparison of the effects of lipolytic and antilipolytic agents on adenosine 3′,5′-monophosphate levels in adipose cells as determined by prior labeling with adenine-8-14C. J. Biol. Chem. 1969, 244, 2252–2260. [Google Scholar] [PubMed]
- Acheson, K.J.; Gremaud, G.; Meirim, I.; Montigon, F.; Krebs, Y.; Fay, L.B.; Gay, L.-J.; Schneiter, P.; Schindler, C.; Tappy, L. Metabolic effects of caffeine in humans: Lipid oxidation or futile cycling? Am. J. Clin. Nutr. 2004, 79, 40–46. [Google Scholar] [PubMed]
- Kim, T.-W.; Shin, Y.-O.; Lee, J.-B.; Min, Y.-K.; Yang, H.-M. Effect of caffeine on the metabolic responses of lipolysis and activated sweat gland density in human during physical activity. Food Sci. Biotechnol. 2010, 19, 1077–1081. [Google Scholar] [CrossRef]
- Lopez-Garcia, E.; van Dam, R.M.; Rajpathak, S.; Willett, W.C.; Manson, J.E.; Hu, F.B. Changes in caffeine intake and long-term weight change in men and women. Am. J. Clin. Nutr. 2006, 83, 674–680. [Google Scholar] [PubMed]
- Westerterp-Plantenga, M.S.; Lejeune, M.P.G.M.; Kovacs, E.M.R. Body weight loss and weight maintenance in relation to habitual caffeine intake and green tea supplementation. Obes. Res. 2005, 13, 1195–1204. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.O.; Shock, N.W. The effect of theophylline ethylene diamine on renal function in control subjects and in patients with congestive heart failure. J. Clin. Investig. 1949, 28 Pt. 2, 1459–1468. [Google Scholar] [CrossRef] [PubMed]
- Osswald, H.; Schnermann, J. Methylxanthines and the kidney. Handb. Exp. Pharmacol. 2011, 200, 391–412. [Google Scholar] [PubMed]
- Wilcox, C.S.; Welch, W.J.; Schreiner, G.F.; Belardinelli, L. Natriuretic and diuretic actions of a highly selective adenosine A1 receptor antagonist. J. Am. Soc. Nephrol. 1999, 10, 714–720. [Google Scholar] [PubMed]
- Rieg, T.; Steigele, H.; Schnermann, J.; Richter, K.; Osswald, H.; Vallon, V. Requirement of intact adenosine A1 receptors for the diuretic and natriuretic action of the methylxanthines theophylline and caffeine. J. Pharmacol. Exp. Ther. 2005, 313, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Hedqvist, P.; Vernet, L. Effect of theophylline and other drugs on rabbit renal cyclic nucleotide phosphodiesterase, 5′-nucleotidase and adenosine deaminase. Biochem. Pharmacol. 1978, 27, 2845–2850. [Google Scholar] [CrossRef]
- Coulson, R.; Scheinman, S.J. Xanthine effects on renal proximal tubular function and cyclic AMP metabolism. J. Pharmacol. Exp. Ther. 1989, 248, 589–595. [Google Scholar] [PubMed]
- Bhupathiraju, S.N.; Pan, A.; Malik, V.S.; Manson, J.E.; Willett, W.C.; van Dam, R.M.; Hu, F.B. Caffeinated and caffeine-free beverages and risk of type 2 diabetes. Am. J. Clin. Nutr. 2013, 97, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Biessels, G.J. Caffeine, diabetes, cognition, and dementia. J. Alzheimers Dis. 2010, 20 (Suppl. S1), S143–S150. [Google Scholar] [PubMed]
- Bidel, S.; Hu, G.; Qiao, Q.; Jousilahti, P.; Antikainen, R.; Tuomilehto, J. Coffee consumption and risk of total and cardiovascular mortality among patients with type 2 diabetes. Diabetologia 2006, 49, 2618–2626. [Google Scholar] [CrossRef] [PubMed]
- Muley, A.; Muley, P.; Shah, M. Coffee to reduce risk of type 2 diabetes? A systematic review. Curr. Diabetes Rev. 2012, 8, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Feskens, E.J.M. Coffee consumption and risk of type 2 diabetes mellitus. Lancet 2002, 360, 1477–1478. [Google Scholar] [CrossRef]
- Van Dam, R.M.; Hu, F.B. Coffee consumption and risk of type 2 diabetes: A systematic review. JAMA 2005, 294, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Paynter, N.P.; Yeh, H.-C.; Voutilainen, S.; Schmidt, M.I.; Heiss, G.; Folsom, A.R.; Brancati, F.L.; Kao, W.H.L. Coffee and sweetened beverage consumption and the risk of type 2 diabetes mellitus: The atherosclerosis risk in communities study. Am. J. Epidemiol. 2006, 164, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, D.S.; Fagherazzi, G.; Balkau, B.; Touillaud, M.S.; Boutron-Ruault, M.-C.; de Lauzon-Guillain, B.; Clavel-Chapelon, F. Differential effects of coffee on the risk of type 2 diabetes according to meal consumption in a French cohort of women: The E3N/EPIC cohort study. Am. J. Clin. Nutr. 2010, 91, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Bhupathiraju, S.N.; Chen, M.; van Dam, R.M.; Hu, F.B. Caffeinated and decaffeinated coffee consumption and risk of type 2 diabetes: A systematic review and a dose-response meta-analysis. Diabetes Care 2014, 37, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Dieren, S.; Uiterwaal, C.S.P.M.; Schouw, Y.T.; van der A, D.L.; Boer, J.M.A.; Spijkerman, A.; Grobbee, D.E.; Beulens, J.W.J. Coffee and tea consumption and risk of type 2 diabetes. Diabetologia 2009, 52, 2561–2569. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.-S.; Wang, W.-Y.; Fan, W.-Y.; Deng, Q.; Wang, X. Tea consumption and risk of type 2 diabetes: A dose–response meta-analysis of cohort studies. Br. J. Nutr. 2014, 111, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.A.; Parker, E.D.; Folsom, A.R. Coffee consumption and risk of type 2 diabetes mellitus: An 11-year prospective study of 28,812 postmenopausal women. Arch. Intern. Med. 2006, 166, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Willett, W.C.; Manson, J.E.; Hu, F.B. Coffee, caffeine, and risk of type 2 diabetes: A prospective cohort study in younger and middle-aged U.S. women. Diabetes Care 2006, 29, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Sarriá, B.; Martínez-López, S.; Sierra-Cinos, J.L.; Garcia-Diz, L.; Goya, L.; Mateos, R.; Bravo, L. Effects of bioactive constituents in functional cocoa products on cardiovascular health in humans. Food Chem. 2015, 174, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Seino, S.; Takahashi, H.; Fujimoto, W.; Shibasaki, T. Roles of cAMP signalling in insulin granule exocytosis. Diabetes Obes. Metab. 2009, 11, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Vilela, V.R.; de Oliveira, A.L.; Comar, J.F.; Peralta, R.M.; Bracht, A. Tadalafil inhibits the cAMP stimulated glucose output in the rat liver. Chem. Biol. Interact. 2014, 220, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.-T.; Xie, J.-G.; Wang, Z.Y.; Ho, C.-T.; Lou, Y.-R.; Wang, C.-X.; Hard, G.C.; Conney, A.H. Effects of tea, decaffeinated tea, and caffeine on UVB light-induced complete carcinogenesis in SKH-1 mice: Demonstration of caffeine as a biologically important constitutent of tea. Cancer Res. 1997, 57, 2623–2629. [Google Scholar] [PubMed]
- Lu, Y.-P.; Lou, Y.-R.; Xie, J.-G.; Peng, Q.-Y.; Liao, J.; Yang, C.S.; Huang, M.-T.; Conney, A.H. Topical applications of caffeine or (−)-epigallocatechin gallate (EGCG) inhibit carcinogenesis and selectively increase apoptosis in UVB-induced skin tumors in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 12455–12460. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.-P.; Lou, Y.-R.; Xie, J.-G.; Peng, Q.-Y.; Zhou, S.; Lin, Y.; Shih, W.J.; Conney, A.H. Caffeine and caffeine sodium benzoate have a sunscreen effect, enhance UVB-induced apoptosis, and inhibit UVB-induced skin carcinogenesis in SKH-1 mice. Carcinogenesis 2006, 28, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Ryzhov, S.; McCaleb, J.L.; Goldstein, A.E.; Biaggioni, I.; Feoktistov, I. Role of Adenosine Receptors in the Regulation of Angiogenic Factors and Neovascularization in Hypoxia. J. Pharmacol. Exp. Ther. 2007, 320, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Chung, F.-L.; Wang, M.; Rivenson, A.; Iatropoulos, M.J.; Reinhardt, J.C.; Pittman, B.; Ho, C.-T.; Amin, S.G. Inhibition of lung carcinogenesis by black tea in fischer rats treated with a tobacco-specific carcinogen: Caffeine as an important constituent. Cancer Res. 1998, 58, 4096–4101. [Google Scholar] [PubMed]
- Lu, G.; Liao, J.; Yang, G.; Reuhl, K.R.; Hao, X.; Yang, C.S. Inhibition of adenoma progression to adenocarcinoma in a 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone—Induced lung tumorigenesis model in A/J mice by tea polyphenols and caffeine. Cancer Res. 2006, 66, 11494–11501. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Rouse, J.; Lukes, L.; Lancaster, M.; Veenstra, T.; Zhou, M.; Shi, Y.; Park, Y.-G.; Hunter, K. Caffeine suppresses metastasis in a transgenic mouse model: A prototype molecule for prophylaxis of metastasis. Clin. Exp. Metastasis 2005, 21, 719–735. [Google Scholar] [CrossRef]
- Gude, R.P.; Menon, L.G.; Rao, S.G. Effect of Caffeine, a xanthine derivative, in the inhibition of experimental lung metastasis induced by B16F10 melanoma cells. J. Exp. Clin. Cancer Res. 2001, 20, 287–292. [Google Scholar] [PubMed]
- Merighi, S.; Benini, A.; Mirandola, P.; Gessi, S.; Varani, K.; Simioni, C.; Leung, E.; Maclennan, S.; Baraldi, P.G.; Borea, P.A. Caffeine inhibits adenosine-induced accumulation of hypoxia-inducible factor-1α, vascular endothelial growth factor, and interleukin-8 expression in hypoxic human colon cancer cells. Mol. Pharmacol. 2007, 72, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Shukla, V.; Gude, R.P. Potentiation of lipid peroxidation in B16F10 and B16F1 melanoma cells by caffeine, a methylxanthine derivative: Relationship to intracellular glutathione. Chemotherapy 2003, 49, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, T.; He, Z.; Ma, W.-Y.; Schmid, P.C.; Bode, A.M.; Yang, C.S.; Dong, Z. Caffeine inhibits cell proliferation by G0/G1 phase arrest in JB6 cells. Cancer Res. 2004, 64, 3344–3349. [Google Scholar] [CrossRef] [PubMed]
- Makower, D.; Malik, U.; Novik, Y.; Wiernik, P.H. Therapeutic efficacy of theophylline in chronic lymphocytic leukemia. Med. Oncol. 1999, 16, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Mitra Basu, R. Theophylline as a therapy for chronic lymphocytic leukemia: A case report and review of literature. Haematologia 2000, 30, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Hirsh, L.; Dantes, A.; Suh, B.-S.; Yoshida, Y.; Hosokawa, K.; Tajima, K.; Kotsuji, F.; Merimsky, O.; Amsterdam, A. Phosphodiesterase inhibitors as anti-cancer drugs. Biochem. Pharmacol. 2004, 68, 981–988. [Google Scholar] [CrossRef] [PubMed]
- Barcz, E.; Sommer, E.; Sokolnicka, I.; Gawrychowski, K.; Roszkowska-Purska, K.; Janik, P.; Skopinska-Rózewska, E. The influence of theobromine on angiogenic activity and proangiogenic cytokines production of human ovarian cancer cells. Oncol. Rep. 1998, 5, 517–537. [Google Scholar] [CrossRef] [PubMed]
- Barcz, E.; Sommer, E.; Janik, P.; Marianowski, L.; Skopinska-Rózewska, E. Adenosine receptor antagonism causes inhibition of angiogenic activity of human ovarian cancer cells. Oncol. Rep. 2000, 7, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Gil, M.; Skopińska-Rózewska, E.; Radomska, D.; Demkow, U.; Skurzak, H.; Rochowska, M.; Beuth, J.; Roszkowski, K. Effect of purinergic receptor antagonists suramin and theobromine on tumor-induced angiogenesis in BALB/c mice. Folia Biol. 1993, 39, 63–68. [Google Scholar]
- Takahashi, M.; Yanoma, S.; Yamamoto, Y.; Rino, Y.; Amano, T.; Imada, T. Combined effect of CDDP and caffeine against human gastric cell line in vivo. Anticancer Res. 1998, 18, 4399–4401. [Google Scholar] [PubMed]
- Fingert, H.J.; Pu, A.T.; Chen, Z.Y.; Perdee, A.B. In vivo and in vitro enhanced antitumour effect by pentoxifylline in human cancer cells treated with Thiothepa. Cancer Res. 1988, 48, 4375–4381. [Google Scholar] [PubMed]
- Fingert, H.J.; Chang, J.D.; Pardee, A.B. Cytotoxic, cell cycle, and chromosomal effects of methylxanthines in human tumor cells treated with alkylating agents. Cancer Res. 1986, 46, 2463–2467. [Google Scholar] [PubMed]
- Kawahara, M.; Takashi, Y.; Takazawa, K.; Tsuchiya, H.; Tomita, K.; Yokogawa, K.; Miyamoto, K.-I. Caffeine Dose-dependently Potentiates the Antitumor Effect of Cisplatin on Osteosarcomas. Anticancer Res. 2008, 28, 1681–1685. [Google Scholar] [PubMed]
- Miwa, S.; Kitamura, S.; Shirai, T.; Hayashi, K.; Nishida, H.; Takeuchi, A.; Nojima, T.; Tsuchiya, H. Desmoplastic small round cell tumour successfully treated with caffeine-assisted chemotherapy: A case report and review of the literature. Anticancer Res. 2010, 30, 3769–3774. [Google Scholar] [PubMed]
- Hayashi, M.; Tsuchiya, H.; Yamamoto, N.; Karita, M.; Shirai, T.; Nishida, H.; Takeuchi, A.; Tomita, K. Caffeine-potentiated chemotherapy for metastatic carcinoma and lymphoma of bone and soft tissue. Anticancer Res. 2005, 25, 2399–2405. [Google Scholar] [PubMed]
- Kakuyama, A.; Sadzuka, Y. Effect of methylxanthine derivatives on doxorubicin transport and antitumor activity. Curr. Drug Metab. 2001, 2, 379–395. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.; Yasutake, H.; Yokogawa, A.; Baba, H.; Ueda, Y.; Tomita, K. Effect of chemotherapy combined with caffeine for osteosarcoma. J. Cancer Res. Clin. Oncol. 1992, 118, 567–569. [Google Scholar] [CrossRef] [PubMed]
- Busse, P.M.; Bose, S.K.; Jones, R.W.; Tolmach, L.J. The action of caffeine on X-Irradiated HeLa cells: III. Enhancement of X-ray-induced killing during G2 arrest. Radiat. Res. 1978, 76, 292–307. [Google Scholar] [CrossRef] [PubMed]
- Malki, A.M.; Gentry, J.; Evans, S.C. Differential effect of selected methylxanthine derivatives on radiosensitization of lung carcinoma cells. Exp. Oncol. 2006, 28, 16–24. [Google Scholar] [PubMed]
- Youn, H.; Hee Kook, Y.; Oh, E.-T.; Jeong, S.-Y.; Kim, C.; Kyung Choi, E.; Uk Lim, B.; Park, H.J. 1-Methylxanthine enhances the radiosensitivity of tumor cells. Int. J. Radiat. Biol. 2009, 85, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.M.; Dong, Z. The enigmatic effects of caffeine in cell cycle and cancer. Cancer Lett. 2007, 247, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Sabisz, M.; Skladanowski, A. Modulation of cellular response to anticancer treatment by caffeine: Inhibition of cell cycle checkpoints, DNA repair and more. Curr. Pharm. Biotechnol. 2008, 9, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.-B.S.; Chaturvedi, P.; Spring, K.; Scott, S.P.; Johanson, R.A.; Mishra, R.; Mattern, M.R.; Winkler, J.D.; Khanna, K.K. Caffeine abolishes the mammalian G2/M DNA damage checkpoint by inhibiting ataxia-telangiectasia-mutated kinase activity. J. Biol. Chem. 2000, 275, 10342–10348. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, T. G2 checkpoint abrogators as anticancer drugs. Mol. Cancer Ther. 2004, 3, 513–519. [Google Scholar] [PubMed]
- Jha, M.N.; Bamburg, J.R.; Bernstein, B.W.; Bedford, J.S. Caffeine eliminates γ-ray-induced G2-phase delay in human tumor cells but not in normal cells. Radiat. Res. 2002, 157, 26–31. [Google Scholar] [CrossRef]
- Dias, T.R.; Alves, M.G.; Bernardino, R.L.; Martins, A.D.; Moreira, A.C.; Silva, J.; Barros, A.; Sousa, M.; Silva, B.M.; Oliveira, P.F. Dose-dependent effects of caffeine in human Sertoli cells metabolism and oxidative profile: Relevance for male fertility. Toxicology 2015, 328, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Kopf, G.S.; Lewis, C.A.; Vacquier, V.D. Characterization of basal and methylxanthine-stimulated Ca2+ transport in abalone spermatozoa. J. Biol. Chem. 1984, 259, 5514–20. [Google Scholar] [PubMed]
- Tash, J.S.; Means, A.R. Regulation of protein phosphorylation and motility of sperm by cyclic adenosine monophosphate and calcium. Biol. Reprod. 1982, 26, 745–763. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, C.J. Regulation of sperm flagellar motility by calcium and cAMP-dependent phosphorylation. J. Cell. Biochem. 1987, 35, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Dias, T.R.; Alves, M.G.; Tomás, G.D.; Socorro, S.; Silva, B.M.; Oliveira, P.F. White tea as a promising antioxidant medium additive for sperm storage at room temperature: A comparative study with green tea. J. Agric. Food Chem. 2014, 62, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Funahashi, H.; Murakami, T. Improved fertility in gilts and sows after artificial insemination of frozen-thawed boar semen by supplementation of semen extender with caffeine and CaCl2. J. Reprod. Dev. 2009, 55, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Klemmer, I.; Yagi, S.; Gressner, O.A. Oral application of 1,7-dimethylxanthine (paraxanthine) attenuates the formation of experimental cholestatic liver fibrosis. Hepatol. Res. 2011, 41, 1094–1109. [Google Scholar] [CrossRef] [PubMed]
- Modi, A.A.; Feld, J.J.; Park, Y.; Kleiner, D.E.; Everhart, J.E.; Liang, T.J.; Hoofnagle, J.H. Increased caffeine consumption is associated with reduced hepatic fibrosis. Hepatology 2010, 51, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Grases, F.; Rodriguez, A.; Costa-Bauza, A. Theobromine inhibits uric acid crystallization. A potential application in the treatment of uric acid nephrolithiasis. PLoS ONE 2014, 9, e111184. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Trier, K.; Zeng, J.; Wu, K.; Yu, M.; Hu, J.; Chen, X.; Ge, J. Effects of 7-methylxanthine on the sclera in form deprivation myopia in guinea pigs. Acta Ophthalmol. 2011, 89, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Trier, K.; Munk Ribel-Madsen, S.; Cui, D.; Brøgger Christensen, S. Systemic 7-methylxanthine in retarding axial eye growth and myopia progression: A 36-month pilot study. J. Ocul. Biol. Dis. Inform. 2008, 1, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Pollard, I.; Williamson, S.; Magre, S. Influence of caffeine administered during pregnancy on the early differentiation of fetal rat ovaries and testes. J. Dev. Physiol. 1990, 13, 59–65. [Google Scholar] [PubMed]
- Chorostowska-Wynimko, J.; Skopińska-Rózewska, E.; Sommer, E.; Rogala, E.; Skopiński, P.; Wojtasik, E. Multiple effects of theobromine on fetus development and postnatal status of the immune system. Int. J. Tissue React. 2004, 26, 53–60. [Google Scholar] [PubMed]
- Skopiński, P.; Woronkowicz, M.; Skopińska-Różewska, E.; Siwicki, A. The effects of prenatal exposure to methylxanthines. Pol. J. Vet. Sci. 2011, 14, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.L.; Bell, E.M.; Druschel, C.M.; Gensburg, L.J.; Mitchell, A.A.; Lin, A.E.; Romitti, P.A.; Correa, A. Maternal caffeine consumption and risk of cardiovascular malformations. Birth Defects Res. Part A Clin. Mol. Teratol. 2007, 79, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Jahanfar, S.; Jaafar, S.H. Effects of restricted caffeine intake by mother on fetal, neonatal and pregnancy outcome. Cochrane Database Syst. Rev. 2013, 2, CD006965. [Google Scholar] [CrossRef] [PubMed]
- Barr, H.M.; Streissguth, A.P. Caffeine use during pregnancy and child outcome: A 7-year prospective study. Neurotoxicol. Teratol. 1991, 13, 441–448. [Google Scholar] [CrossRef]
- Bech, B.H.; Obel, C.; Henriksen, T.B.; Olsen, J. Effect of reducing caffeine intake on birth weight and length of gestation: Randomised controlled trial. BMJ Br. Med. J. 2007, 334, 409–409. [Google Scholar] [CrossRef] [PubMed]
- Browne, M.L.; Hoyt, A.T.; Feldkamp, M.L.; Rasmussen, S.A.; Marshall, E.G.; Druschel, C.M.; Romitti, P.A. Maternal caffeine intake and risk of selected birth defects in the national birth defects prevention study. Birth Defects Res. Part A Clin. Mol. Teratol. 2011, 91, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Group, C.S. Maternal caffeine intake during pregnancy and risk of fetal growth restriction: A large prospective observational study. BMJ Br. Med. J. 2008, 337, a2332. [Google Scholar] [CrossRef]
- Weng, X.; Odouli, R.; Li, D.-K. Maternal caffeine consumption during pregnancy and the risk of miscarriage: A prospective cohort study. Am. J. Obstet. Gynecol. 2008, 198, 279.e1–279.e8. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Romitti, P.A.; Burns, T.L.; Browne, M.L.; Druschel, C.M.; Olney, R.S. Maternal caffeine consumption and risk of neural tube defects. Birth Defects Res. Part A Clin. Mol. Teratol. 2009, 85, 879–889. [Google Scholar] [CrossRef] [PubMed]
- Hoyt, A.T.; Browne, M.; Richardson, S.; Romitti, P.; Druschel, C. Maternal caffeine consumption and small for gestational age births: Results from a population-based case-control study. Matern. Child Health J. 2013, 18, 1540–1551. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Bell, E.M.; Browne, M.L.; Druschel, C.M.; Romitti, P.A.; Schmidt, R.J.; Burns, T.L.; Moslehi, R.; Olney, R.S.; National Birth Defects Prevention Study. Maternal caffeine consumption and risk of congenital limb deficiencies. Birth Defects Res. Part A Clin. Mol. Teratol. 2012, 94, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Kuczkowski, K.M. Caffeine in pregnancy. Arch. Gynecol. Obstet. 2009, 280, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Friedman, L.; Weinberger, M.A.; Farber, T.M.; Moreland, F.M.; Peters, E.L.; Gilmore, C.E.; Khan, M.A. Testicular atrophy and impaired spermatogenesis in rats fed high levels of the methylxanthines caffeine, theobromine, or theophylline. J. Environ. Pathol. Toxicol. 1979, 2, 687–706. [Google Scholar] [PubMed]
- Gans, J.H. Comparative toxicities of dietary caffeine and theobromine in the rat. Food Chem. Toxicol. 1984, 22, 365–369. [Google Scholar] [CrossRef]
- Cappelletti, S.; Daria, P.; Sani, G.; Aromatario, M. Caffeine: Cognitive and physical performance enhancer or psychoactive drug? Curr. Neuropharmacol. 2015, 13, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Hart, A.B.; de Wit, H.; Palmer, A.A. Genetic factors modulating the response to stimulant drugs in humans. Curr. Top. Behav. Neurosci. 2012, 12, 537–577. [Google Scholar] [PubMed]
- Rogers, P.J.; Hohoff, C.; Heatherley, S.V.; Mullings, E.L.; Maxfield, P.J.; Evershed, R.P.; Deckert, J.; Nutt, D.J. Association of the anxiogenic and alerting effects of caffeine with ADORA2A and ADORA1 polymorphisms and habitual level of caffeine consumption. Neuropsychopharmacology 2010, 35, 1973–1983. [Google Scholar] [CrossRef] [PubMed]
- Alsene, K.; Deckert, J.; Sand, P.; de Wit, H. Association between A2A receptor gene polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2003, 28, 1694–1702. [Google Scholar] [CrossRef] [PubMed]
- Planning Committee for a Workshop on Potential Health Hazards Associated with Consumption of Caffeine in Food and Dietary Supplements; Food and Nutrition Board; Board on Health Sciences Policy; Institute of Medicine. In Caffeine in Food and Dietary Supplements: Examining Safety: Workshop Summary; National Academies Press (US): Washington DC, USA, 2014.
- Glade, M.J. Caffeine—Not just a stimulant. Nutrition 2010, 26, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Clauson, K.A.; Shields, K.M.; McQueen, C.E.; Persad, N. Safety issues associated with commercially available energy drinks. J. Am. Pharm. Assoc. 2003, 48, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Ali, Z.; Levine, B.; Fowler, D.R. Fatal caffeine intoxication: A series of eight cases from 1999 to 2009. J. Forensic Sci. 2014, 59, 865–868. [Google Scholar] [CrossRef] [PubMed]
- Stavric, B. Methylxanthines: Toxicity to humans. 2. Caffeine. Food Chem. Toxicol. 1988, 26, 645–662. [Google Scholar] [CrossRef]
- Tajima, Y. Coffee-induced hypokalaemia. Clin. Med. Insights Case Rep. 2010, 3, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A. Dependence upon coffee and caffeine: An update. In Coffee, Tea, Chocolate, and the Brain; Nehlig, A., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 133–146. [Google Scholar]
- Nehlig, A.; Armspach, J.-P.; Namer, I.J. SPECT assessment of brain activation induced by caffeine: No effect on areas involved in dependence. Dialogues Clin. Neurosci. 2010, 12, 255–263. [Google Scholar] [PubMed]
- Baggott, M.J.; Childs, E.; Hart, A.B.; de Bruin, E.; Palmer, A.A.; Wilkinson, J.E.; de Wit, H. Psychopharmacology of theobromine in healthy volunteers. Psychopharmacology 2013, 228, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Eteng, M.U.; Eyong, E.U.; Akpanyung, E.O.; Agiang, M.A.; Aremu, C.Y. Recent advances in caffeine and theobromine toxicities: A review. Plant Foods Hum. Nutr. 1997, 51, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Czok, G. Concerning the question of the biological effectiveness of methylxanthines in cocoa products. Z. Ernährungswissenschaft 1974, 4, 165–170. [Google Scholar] [CrossRef]
- Knight, R.J.; Bowmer, C.J.; Yates, M.S. The diuretic action of 8-cyclopentyl-1,3-dipropylxanthine, a selective A1 adenosine receptor antagonist. Br. J. Pharmacol. 1993, 109, 271–277. [Google Scholar] [CrossRef] [PubMed]




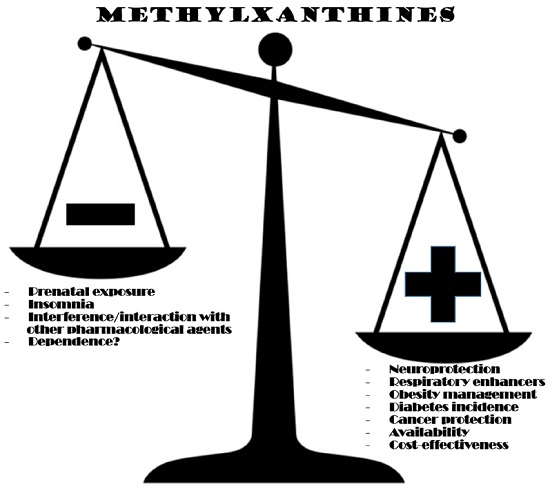{kind=link}
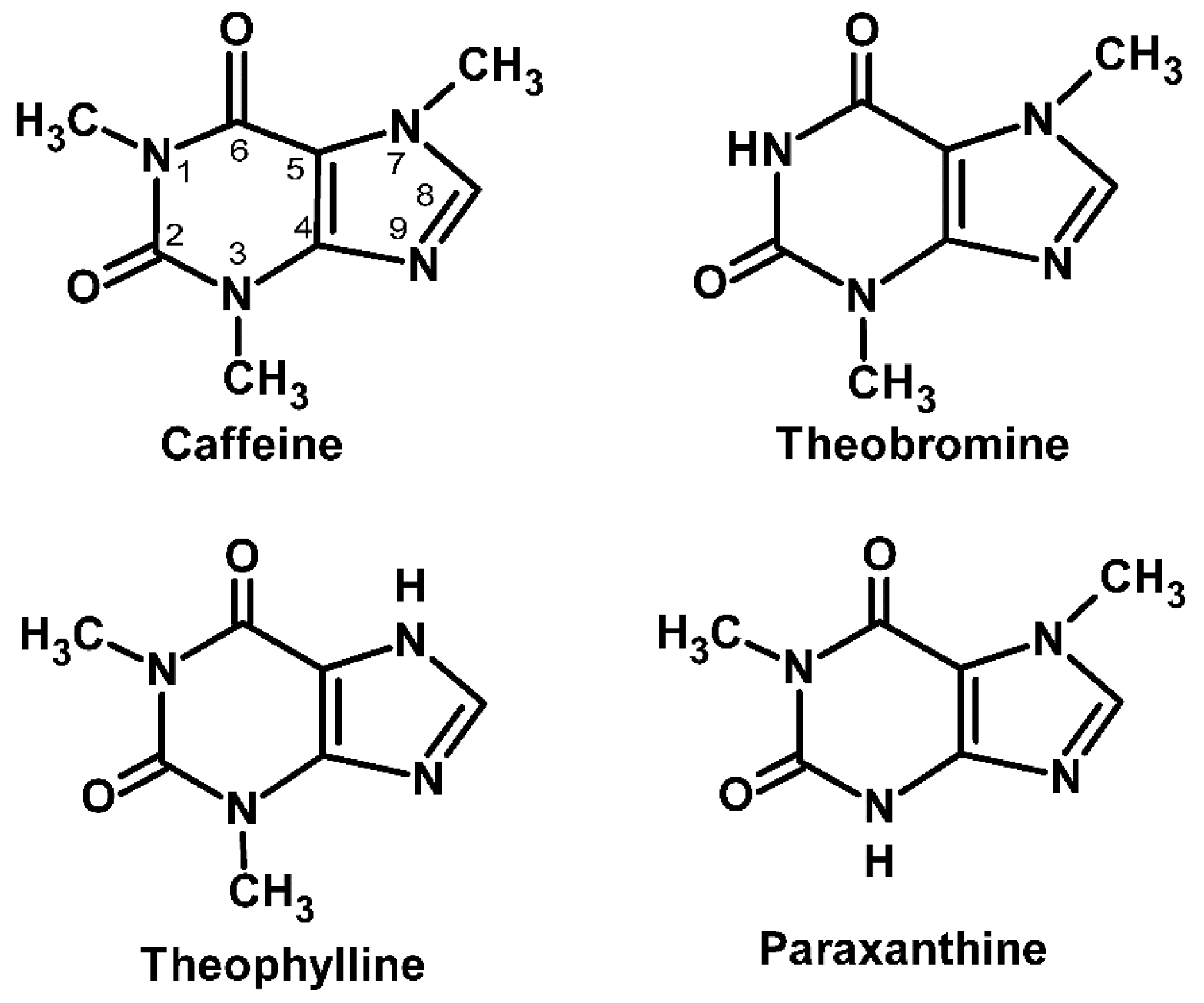{kind=link}
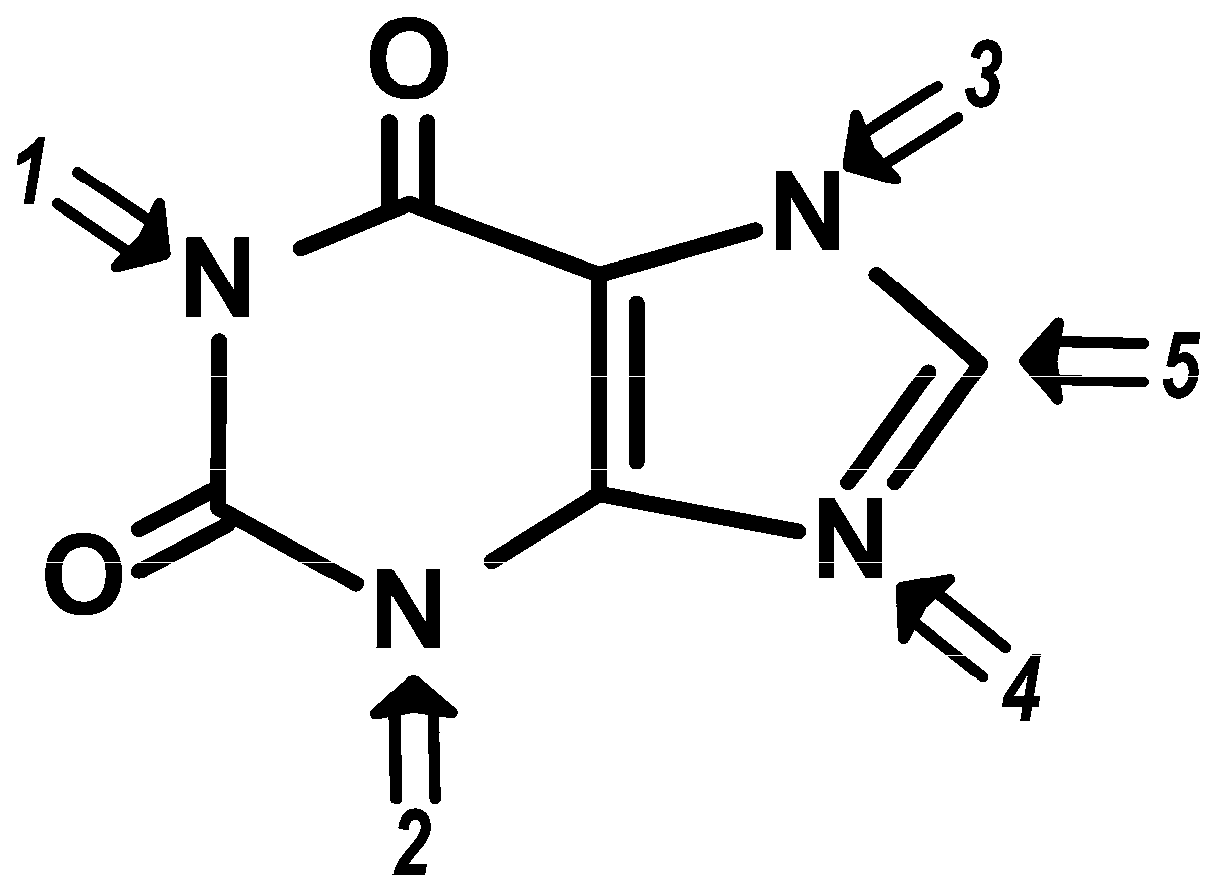{kind=link}
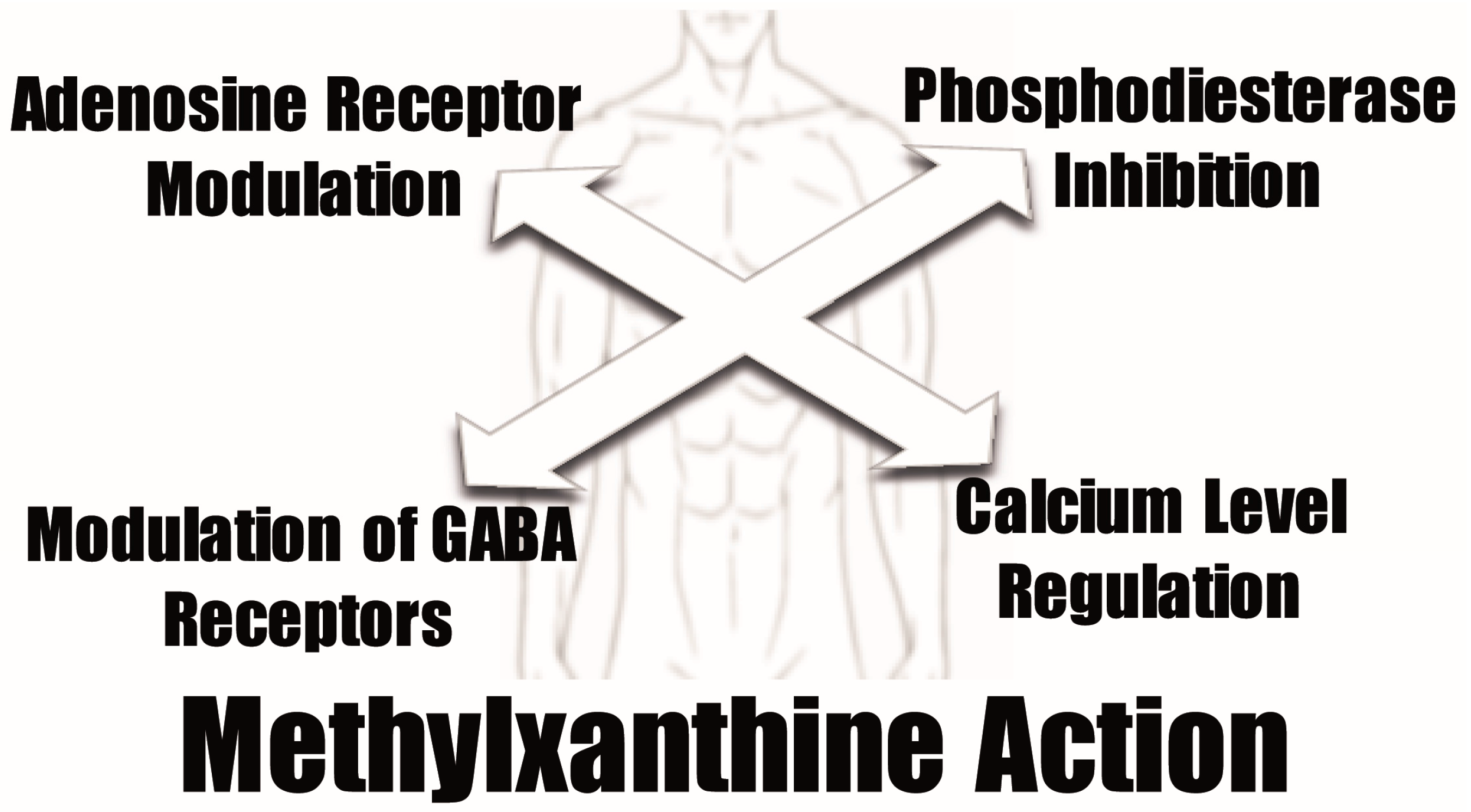{kind=link}
| Dietary Source | Medium Content (mg) | |||
|---|---|---|---|---|
| Caffeine | Theobromine | Theophylline | ||
| Coffee | Expresso (23–70 mL) | 140 (51–532 range) [46] | -------- * | -------- |
| Decaffeinated (30 mL) | 3.0–15.8 range [47] | -------- | -------- | |
| Tea | Black (200–235 mL) | 55.7 (41.6–71.2 range) [48] | 1.8–3.6 [49] | <1 [50] |
| Green (200–235 mL) | 31.0 (20.2–42.8 range) [48] | 0.2–0.7 [51] | -------- | |
| Chocolate | Dark (100 g) | 0.22–0.31 range [52] | 0.26 (0.26–1.16 range) [53] | 0–9 [54] |
| Milk (100 g) | 0.05–0.17 range [52] | 0.17 (0.09–0.28 range) [53] | 5 [54] | |
| Soft-Drink | Cola (330 mL) | 32 | -------- | -------- |
| Diet Cola (330 mL) | 42 | -------- | -------- | |
| Energy Drinks | Red Bull (250 mL) | 80 | -------- | -------- |
| Methylxanthine | [M + H]+ | MS Data (MS/MS Fragmentation Pattern) |
|---|---|---|
| Caffeine | 195.2 | 181.2, 151.2, 138.0 |
| Theophylline | 181.2 | 167.2, 153.2, 123.5 |
| Theobromine | 181.2 | 167.2, 153.2, 107.5 |
| Systematic Effect | Caffeine | Theobromine | Theophylline |
|---|---|---|---|
| CNS Stimulation | +++ | + | ++ |
| Respiratory Stimulation | +++ | + | ++ |
| Diuresis | ++ | + | +++ |
| Coronary Dilatation | + | ++ | +++ |
| Cardiac Stimulation | + | ++ | +++ |
| Skeletal Muscle Stimulation | +++ | + | ++ |
| Smooth Muscle Relaxation (Bronchodilation) | + | + | +++ |
| LD50 (oral, mg/mL) | |||||
|---|---|---|---|---|---|
| Man | Rat | Mouse | Dog | Cat | |
| Caffeine | 150–200 a | 200 | 127 | 145 b | 125 b |
| Theobromine | 1000 | 950 | 135 b | 300 | 200 |
| Theophylline | (no data available) | 206 | 332 | 300 | 700 |
| Molecular Target | Effective Concentrations * | Mediated Therapeutics | Legitimacy of Evidence |
|---|---|---|---|
| Adenosine receptors | 50–55 μM for caffeine, 14 μM for theophylline [122] | asthma [254,255], apnea of prematurity [246,247,248,249,250,251], cough [257,258], diuretic [282,283,377], analgesic [204] | from in vitro studies to clinical trials (therapeutic effects) |
| Phosphodiesterase | 55 μM for theophylline [256] | asthma [19,253], chronic obstructive pulmonary disease [256], apnea of prematurity [252], cough [257,258], diuretic [284,285], chronic lymphocytic leucemia [313] | from in vitro studies to clinical trials (therapeutic effects) |
| GABA receptors | 500 μM for caffeine [167] | ---------------- ** | in vitro studies |
| Ryanodine-sensitive calcium channels | >1 mM for cafeine [170] | ---------------- | in vitro studies |
| Poly(ADPribose) polymerase-1 | 160 μM for theobromine, 195 μM for theophylline and 200 μM for caffeine [182] | ---------------- | in vitro studies |
| Chitinases | 469 μM for caffeine and 1.5 mM theophylline [183] | ---------------- | in vitro studies |
| Histone deacetylases | 24 μM for theophylline [184] | asthma, anti-inflammatory [184] | double-blind crossover controlled study |
| Acetylcholinesterase | 175 μM for cafeine [186] | ---------------- | in vitro studies |
| Monoamine oxidase B | 700 μM for cafeine [188] | ---------------- | in vitro studies |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, J.P.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks. Molecules 2016, 21, 974. https://doi.org/10.3390/molecules21080974
Monteiro JP, Alves MG, Oliveira PF, Silva BM. Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks. Molecules. 2016; 21(8):974. https://doi.org/10.3390/molecules21080974
Chicago/Turabian StyleMonteiro, João P., Marco G. Alves, Pedro F. Oliveira, and Branca M. Silva. 2016. "Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks" Molecules 21, no. 8: 974. https://doi.org/10.3390/molecules21080974
APA StyleMonteiro, J. P., Alves, M. G., Oliveira, P. F., & Silva, B. M. (2016). Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks. Molecules, 21(8), 974. https://doi.org/10.3390/molecules21080974