
Protocatechualdehyde Induces S-Phase Arrest and Apoptosis by Stimulating the p27KIP1-Cyclin A/D1-CDK2 and Mitochondrial Apoptotic Pathways in HT-29 Cells

Abstract
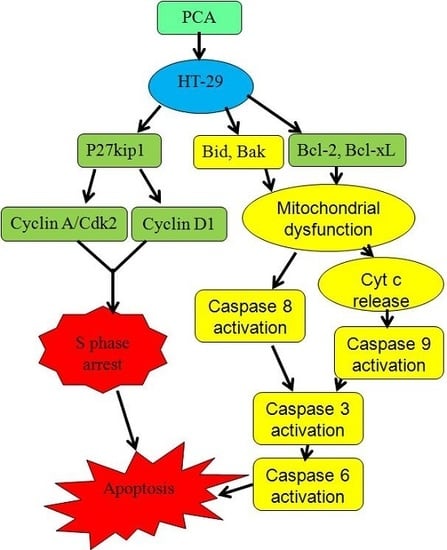
:
1. Introduction
2. Results
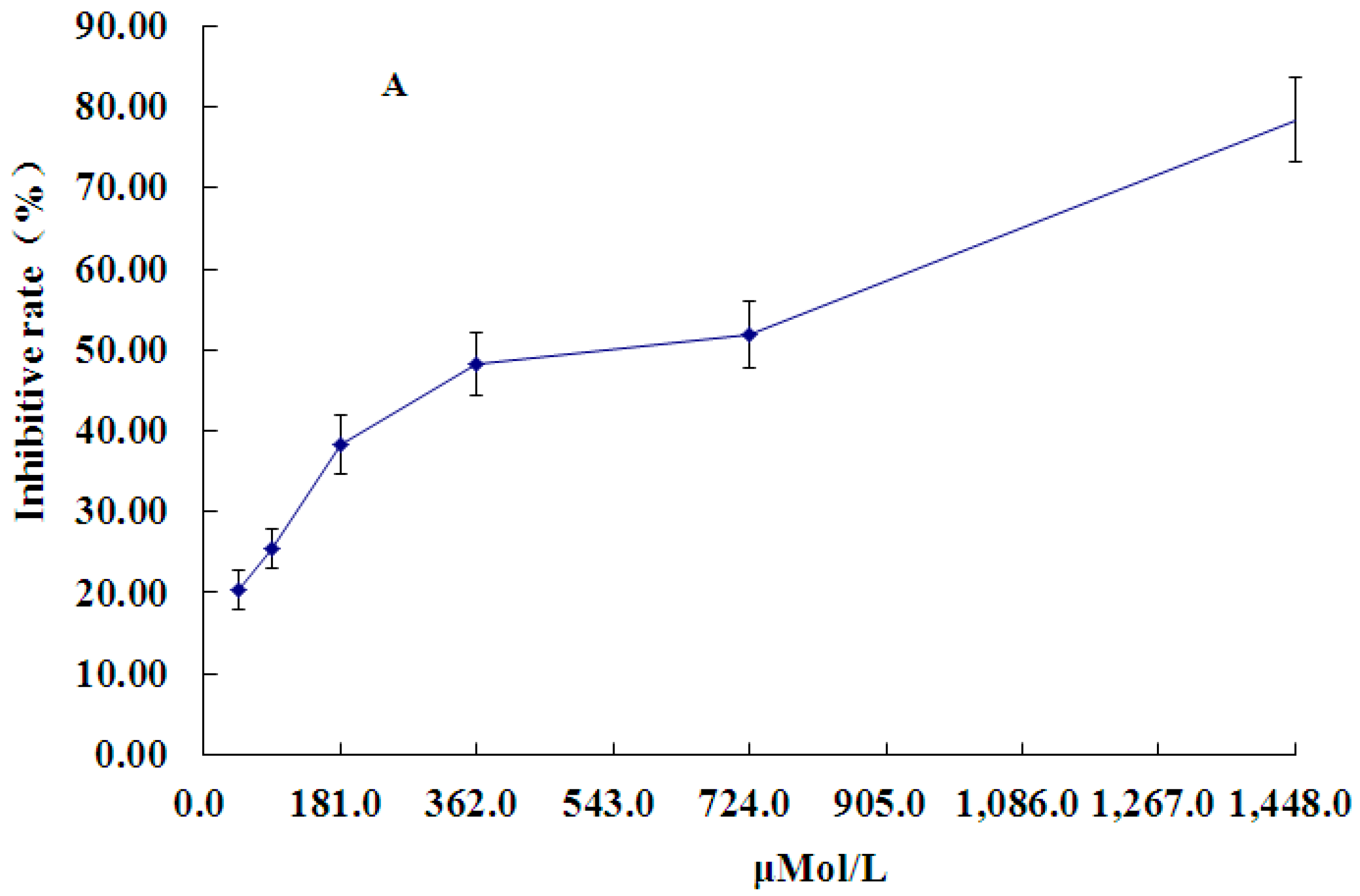
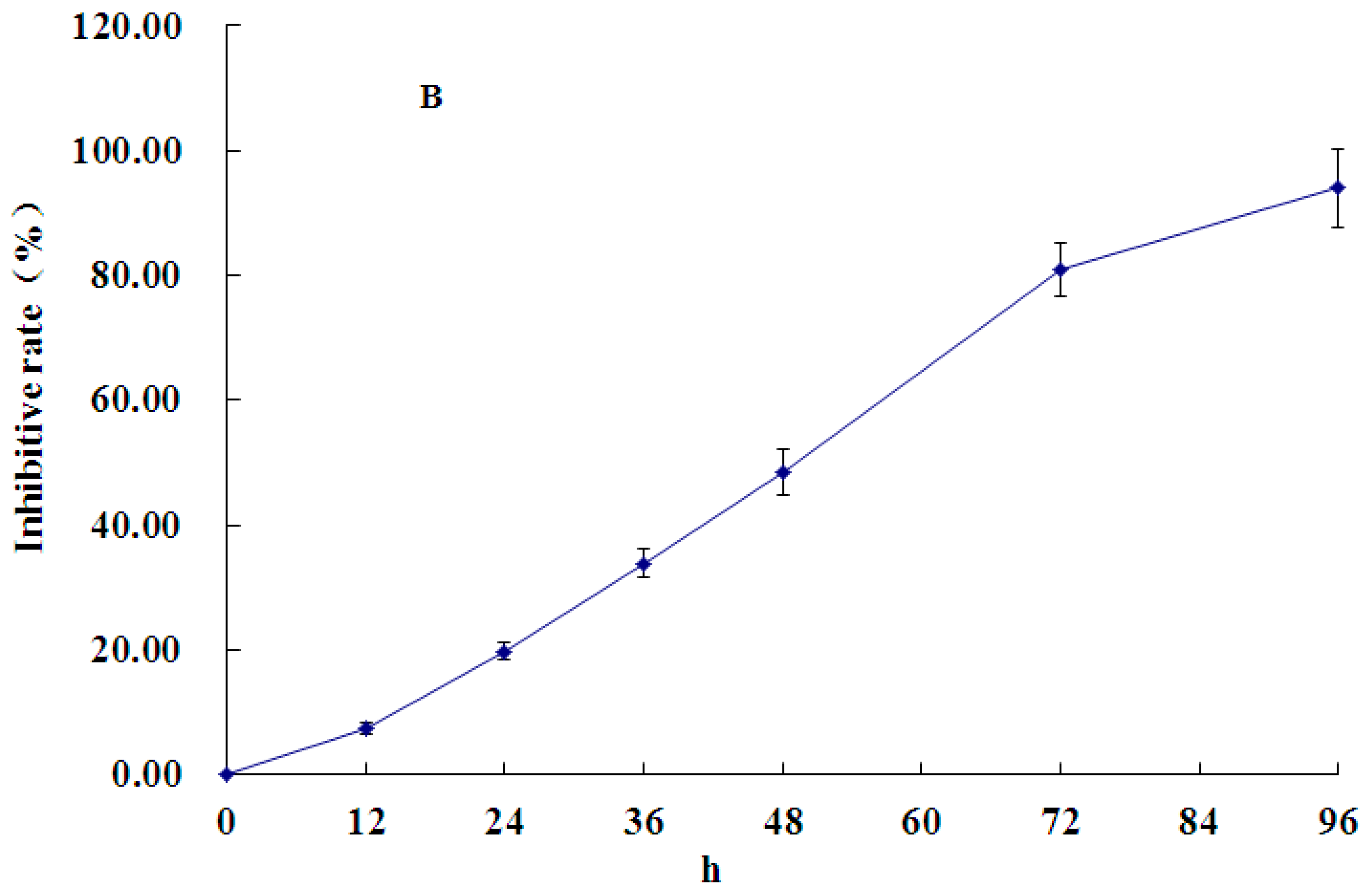
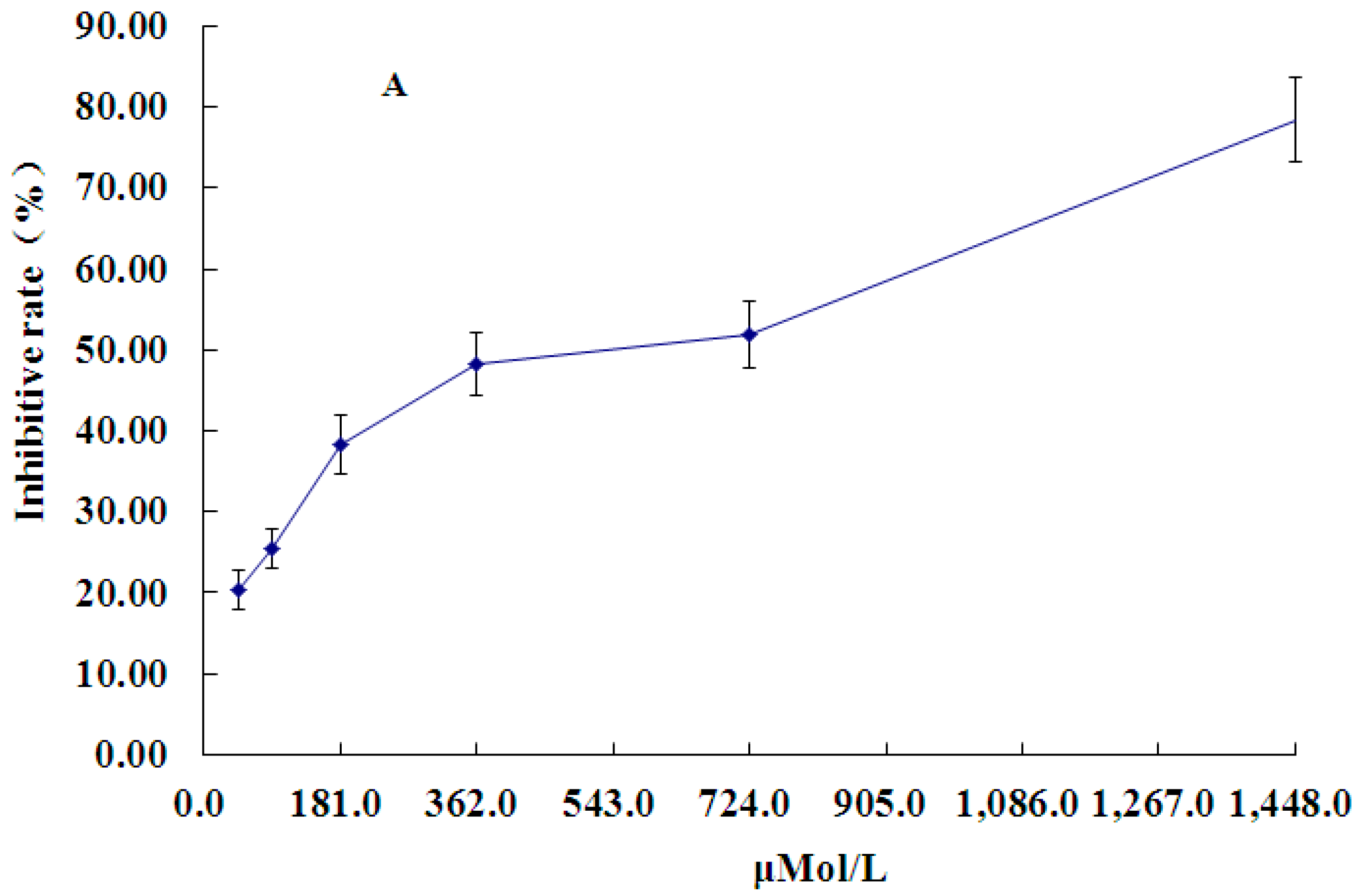
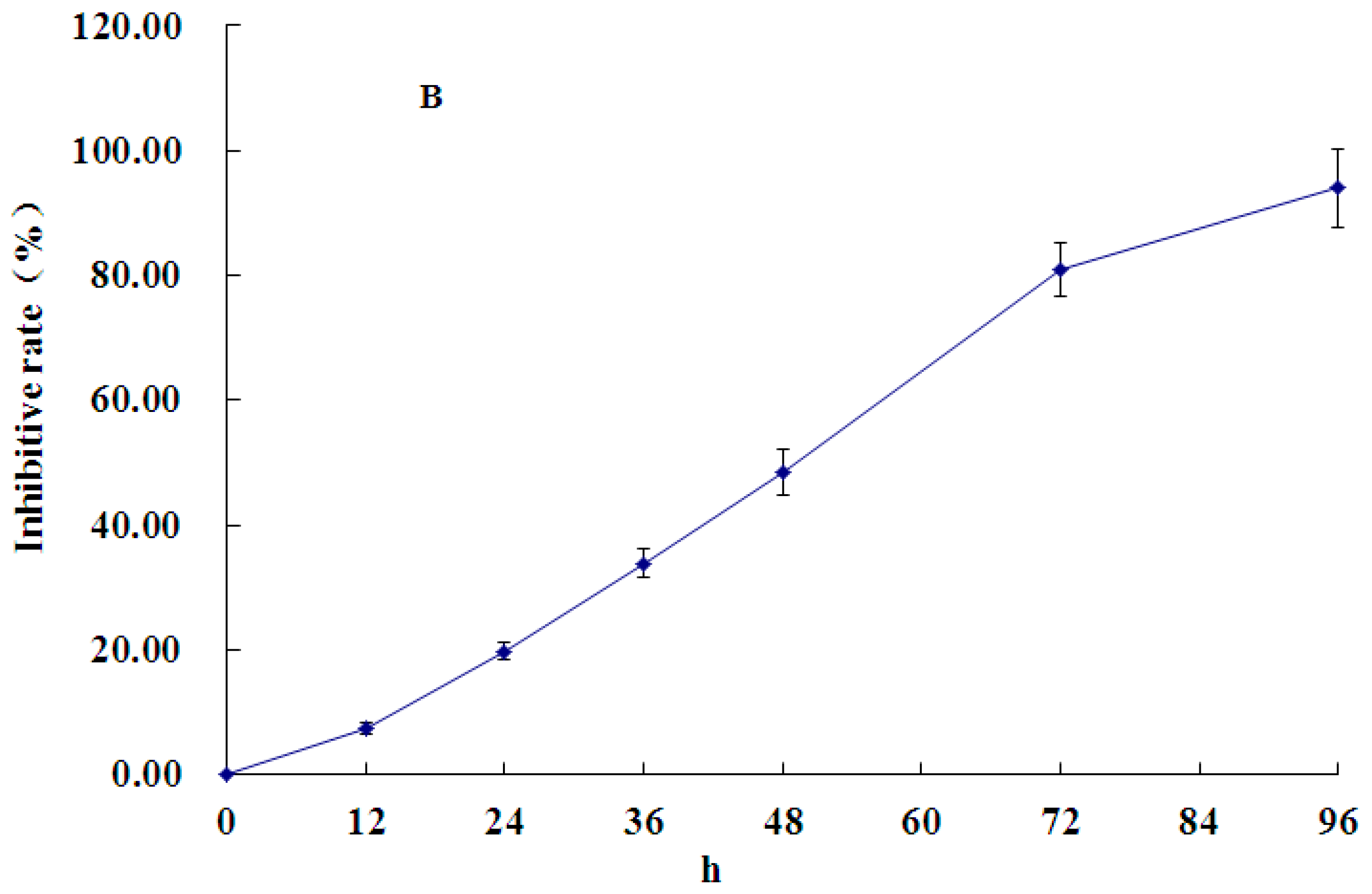
2.1. Effect of PCA on HT-29 Cell Proliferation
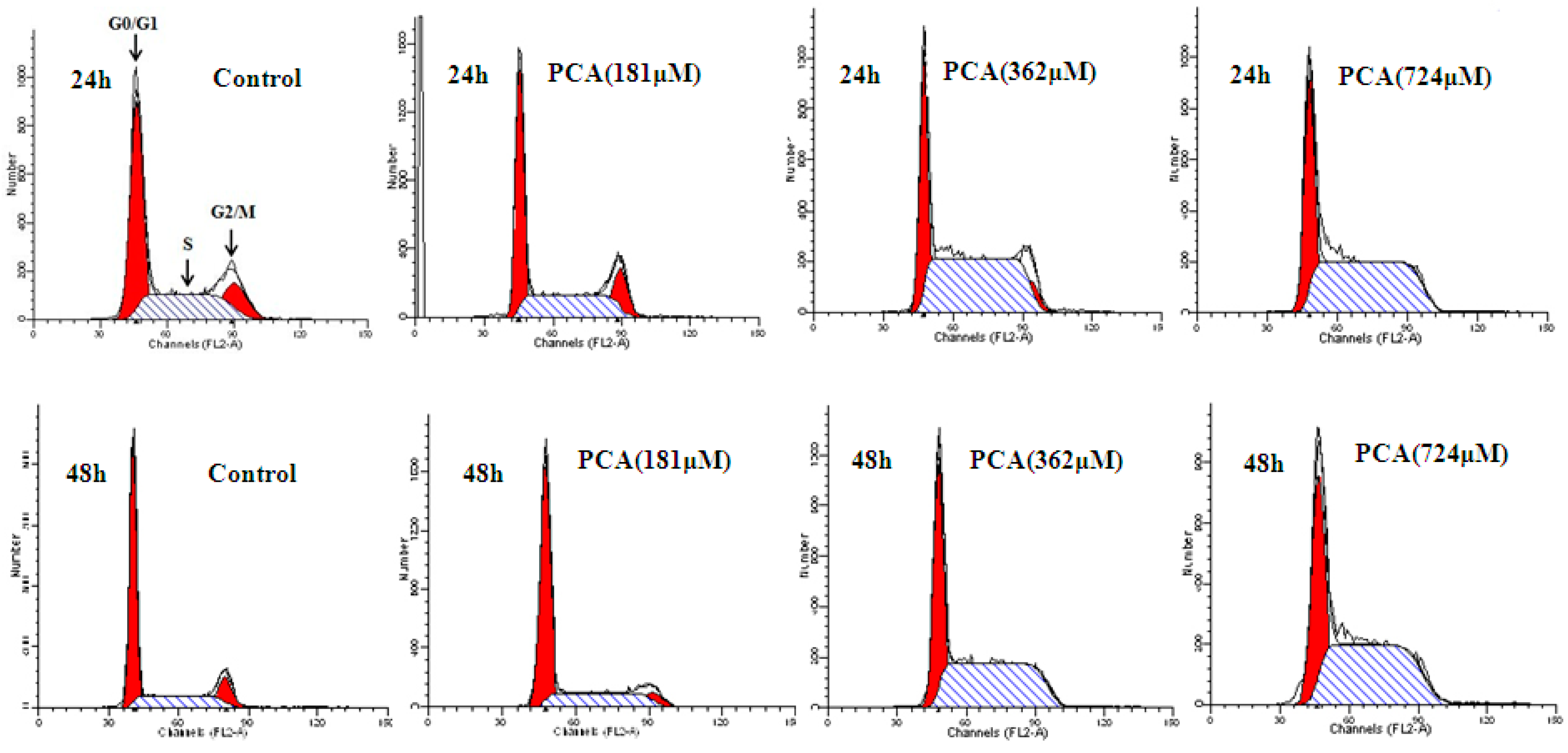
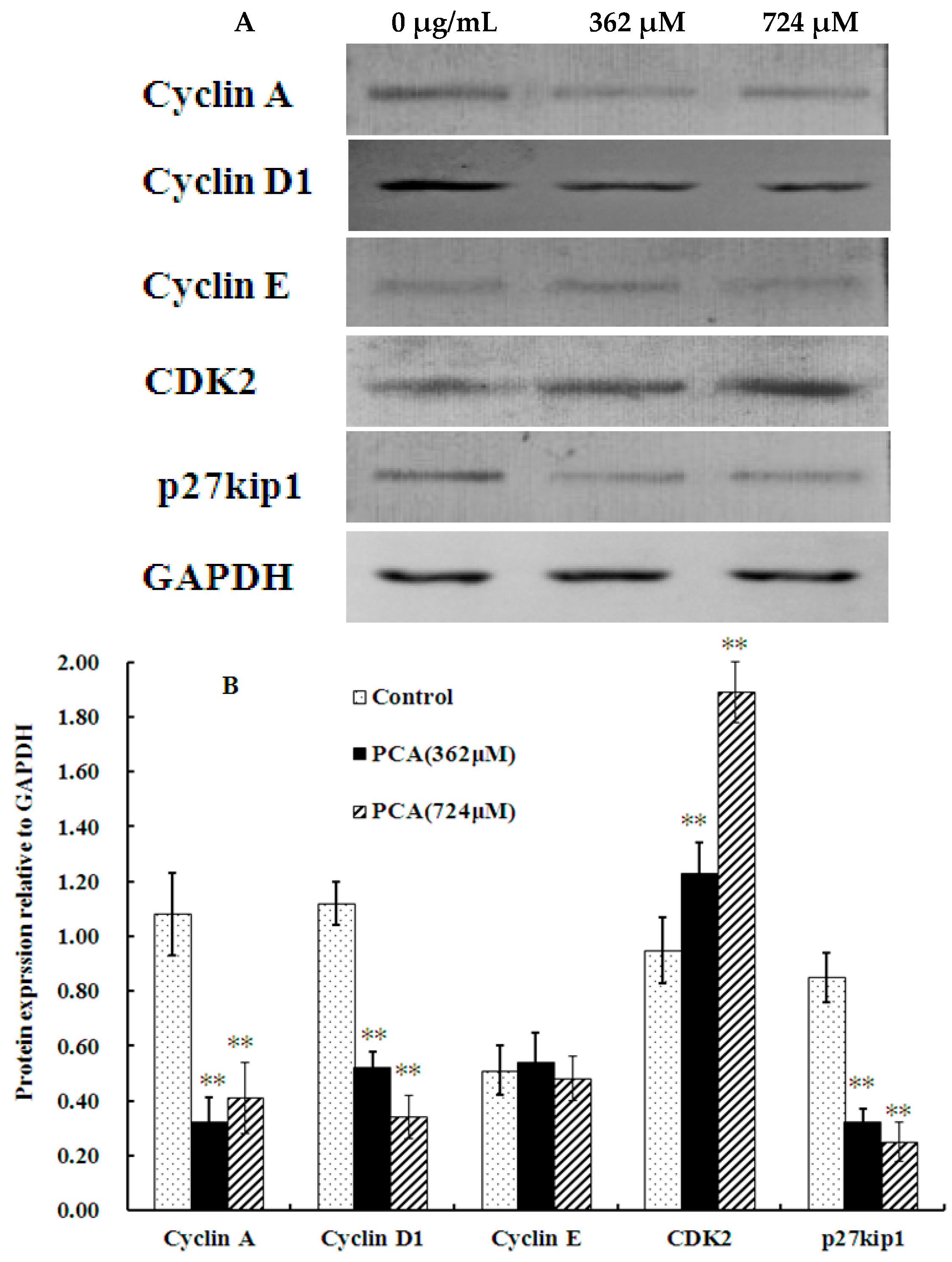
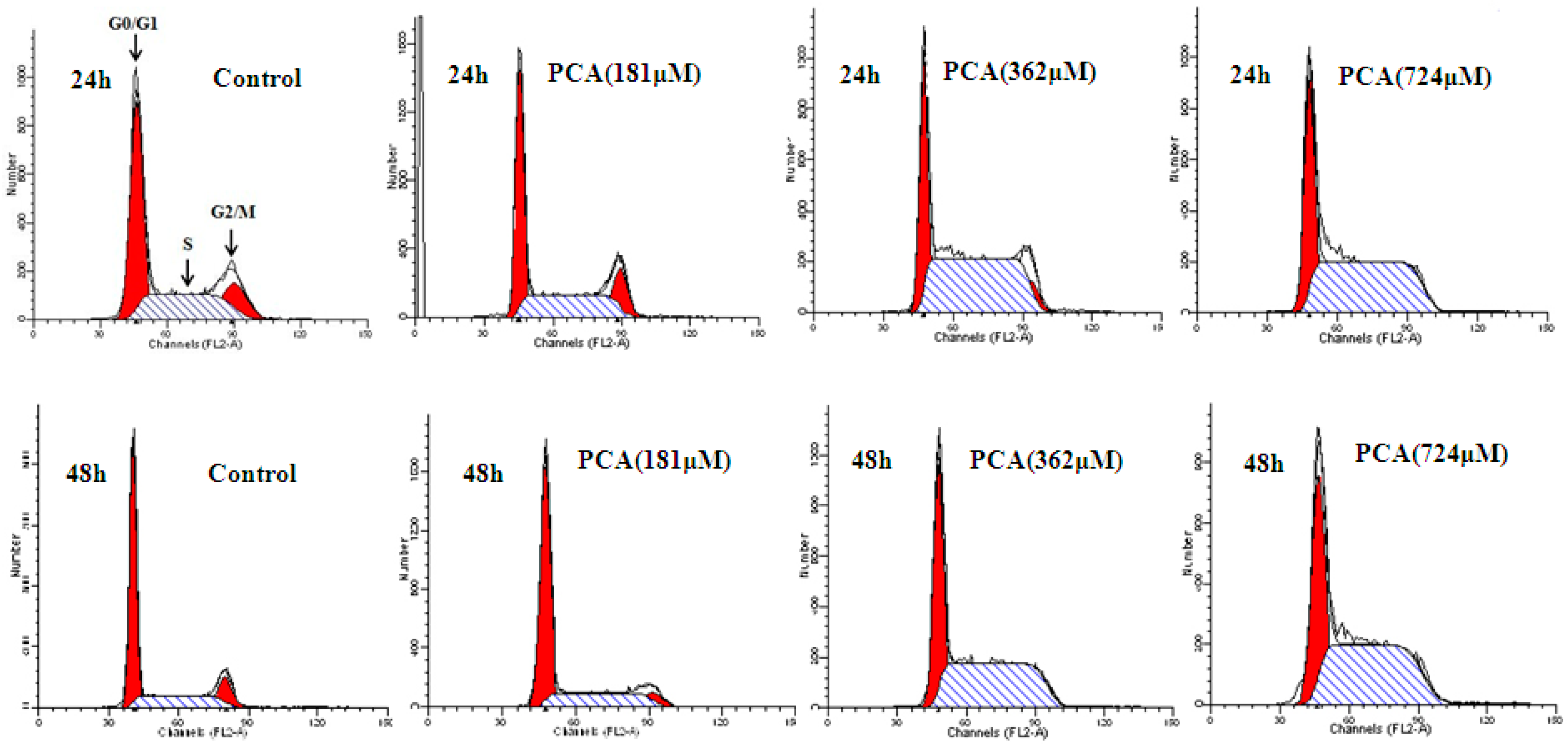
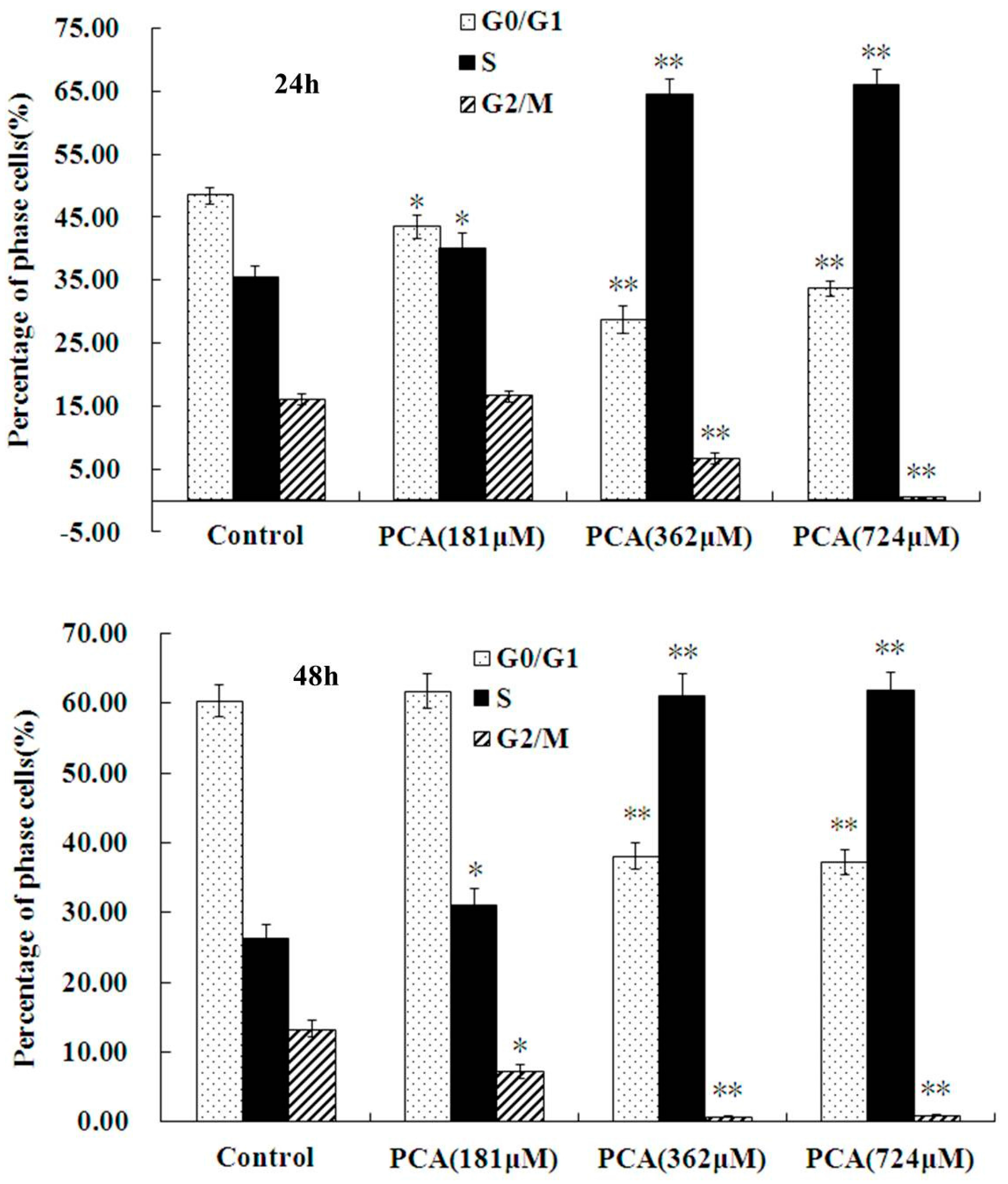
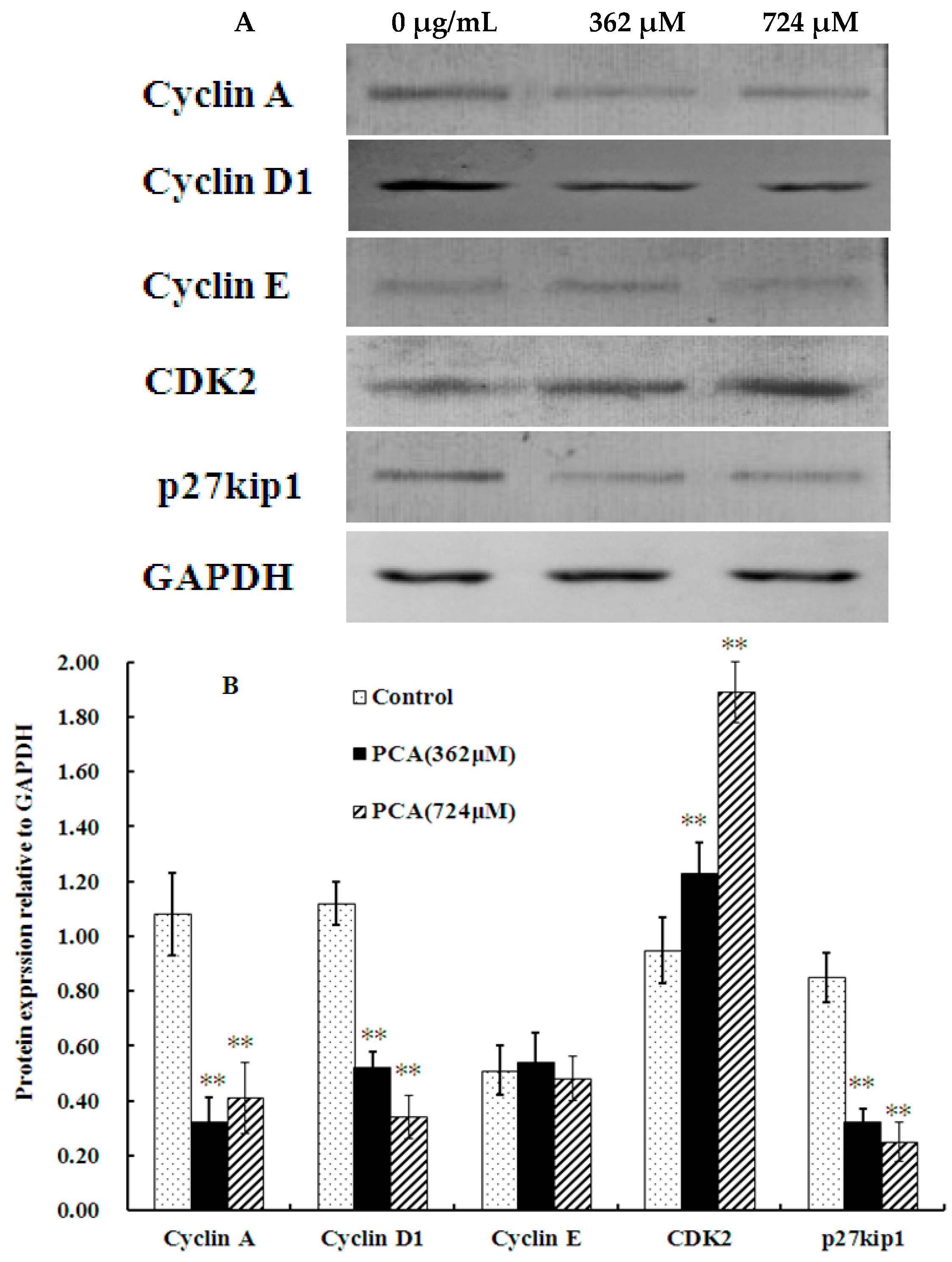
2.2. PCA Induced S-Phase Cell Cycle Arrest
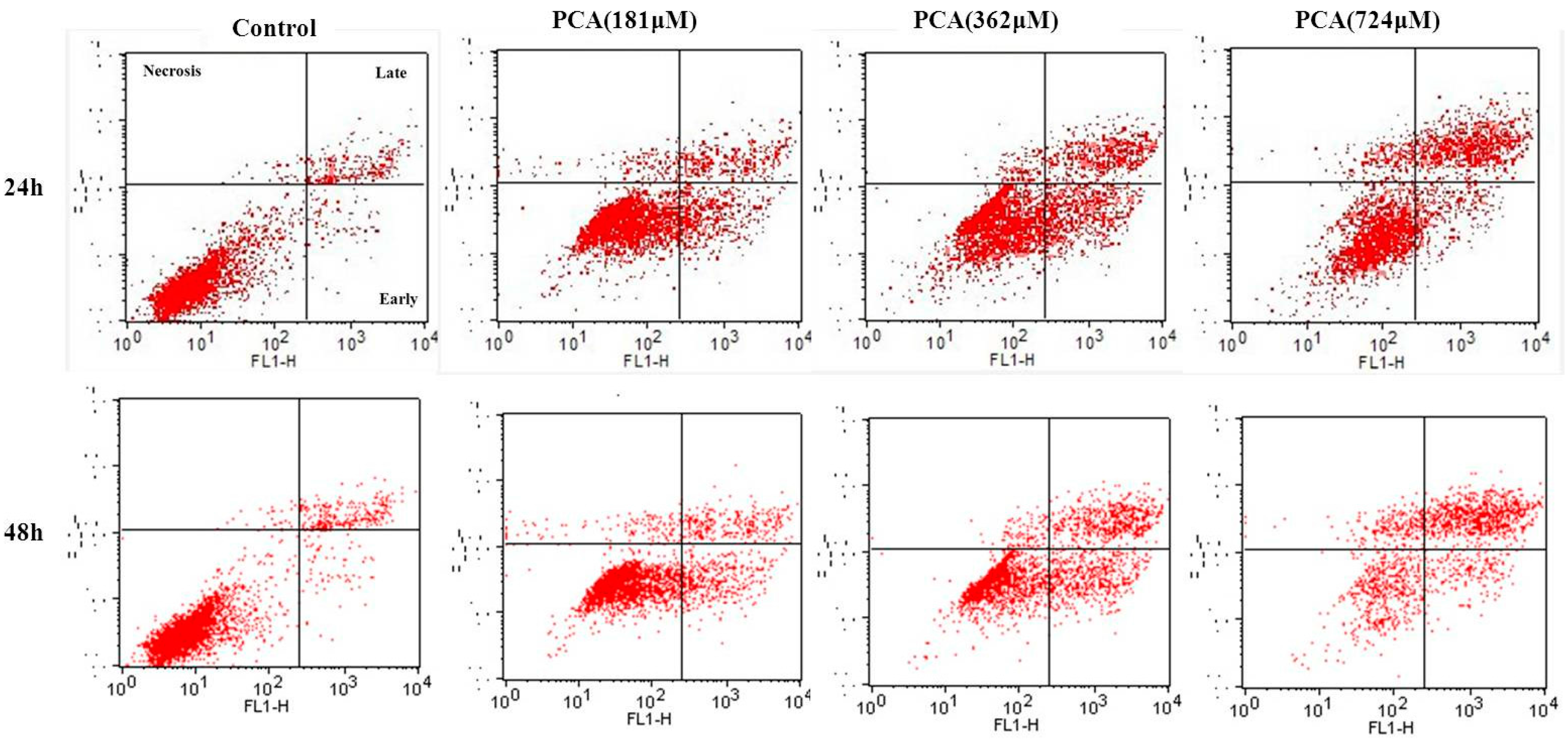
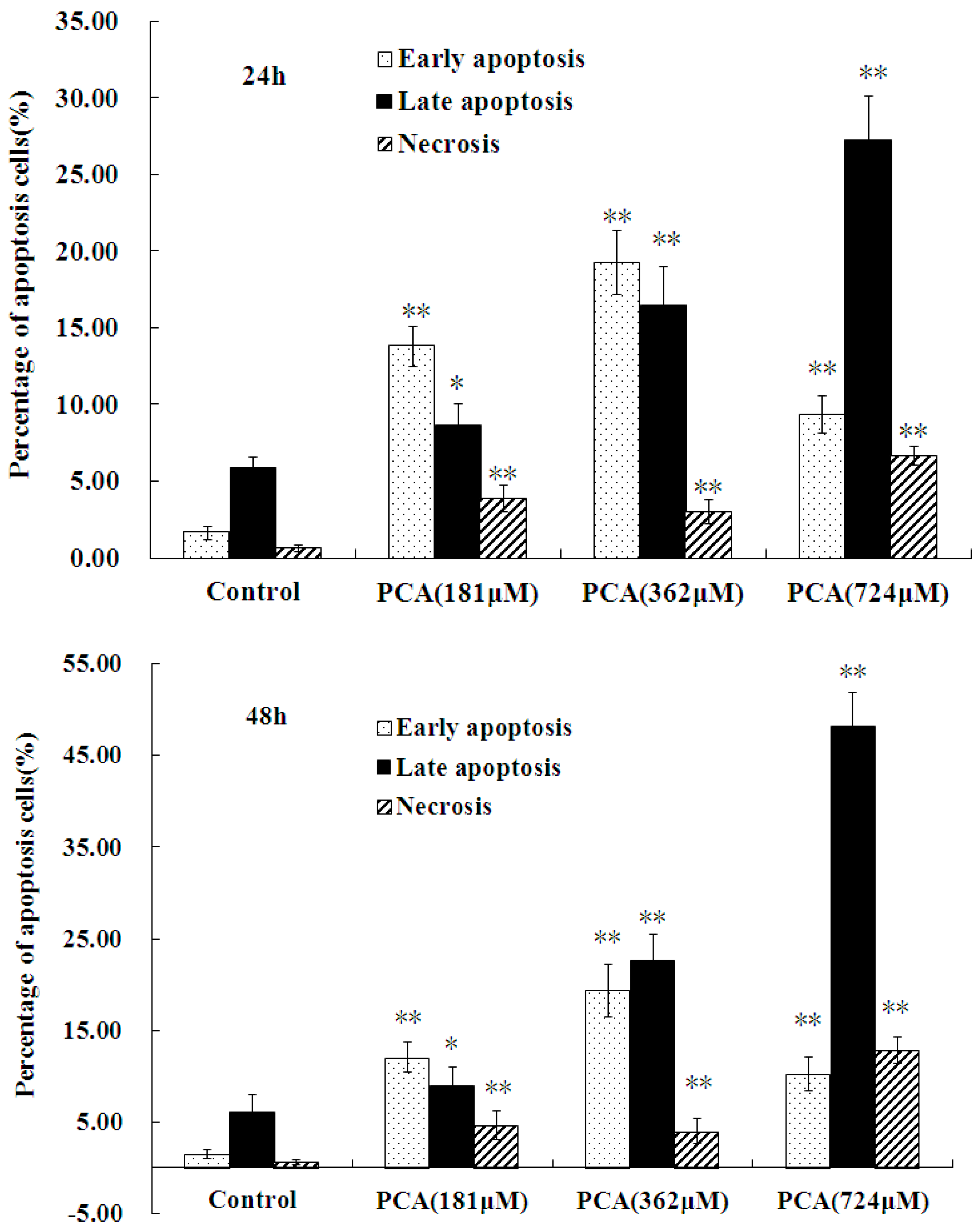
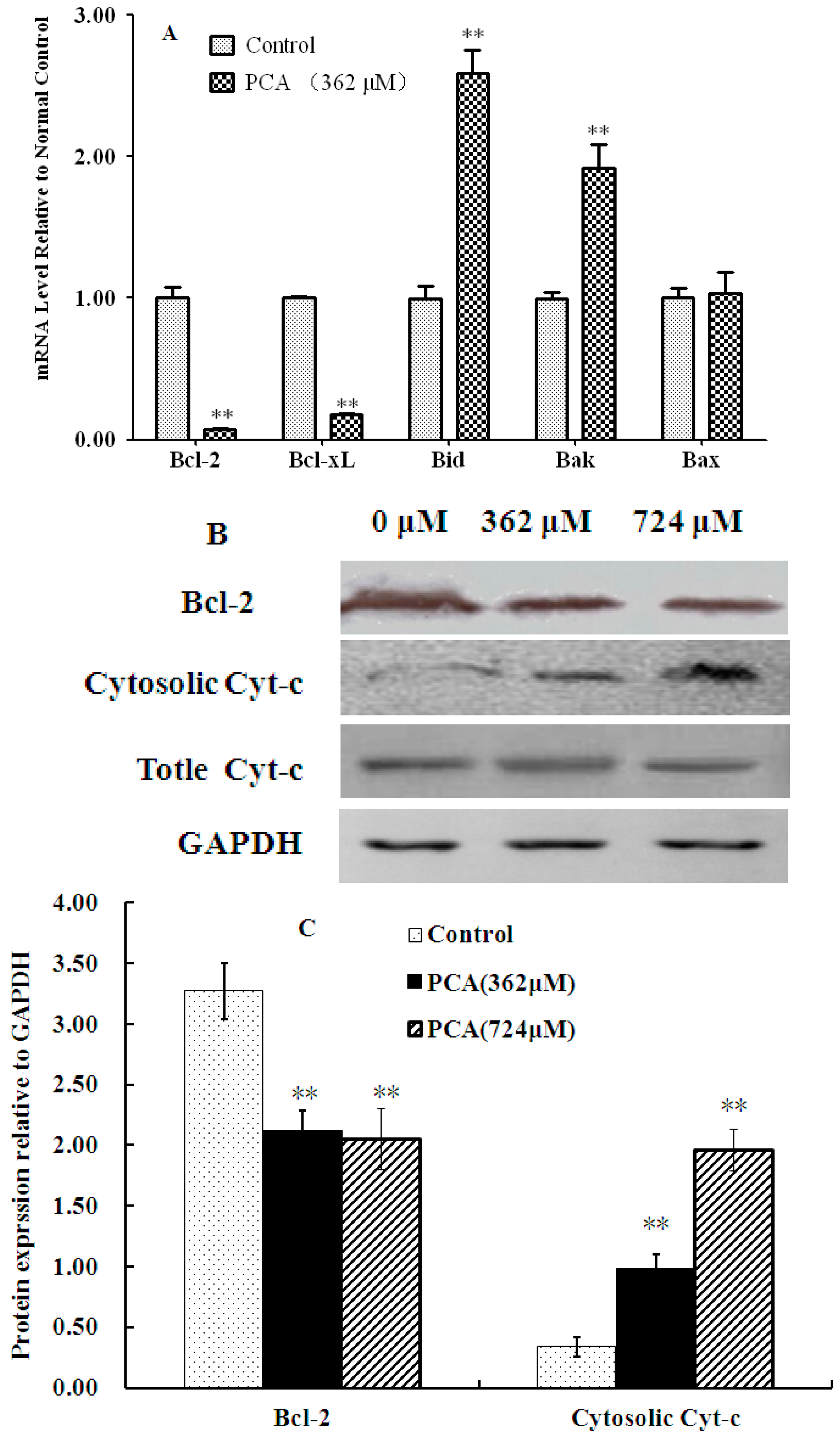
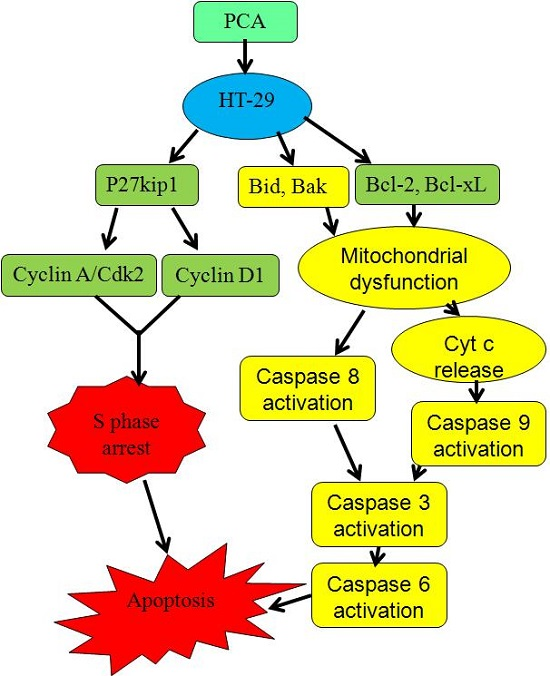
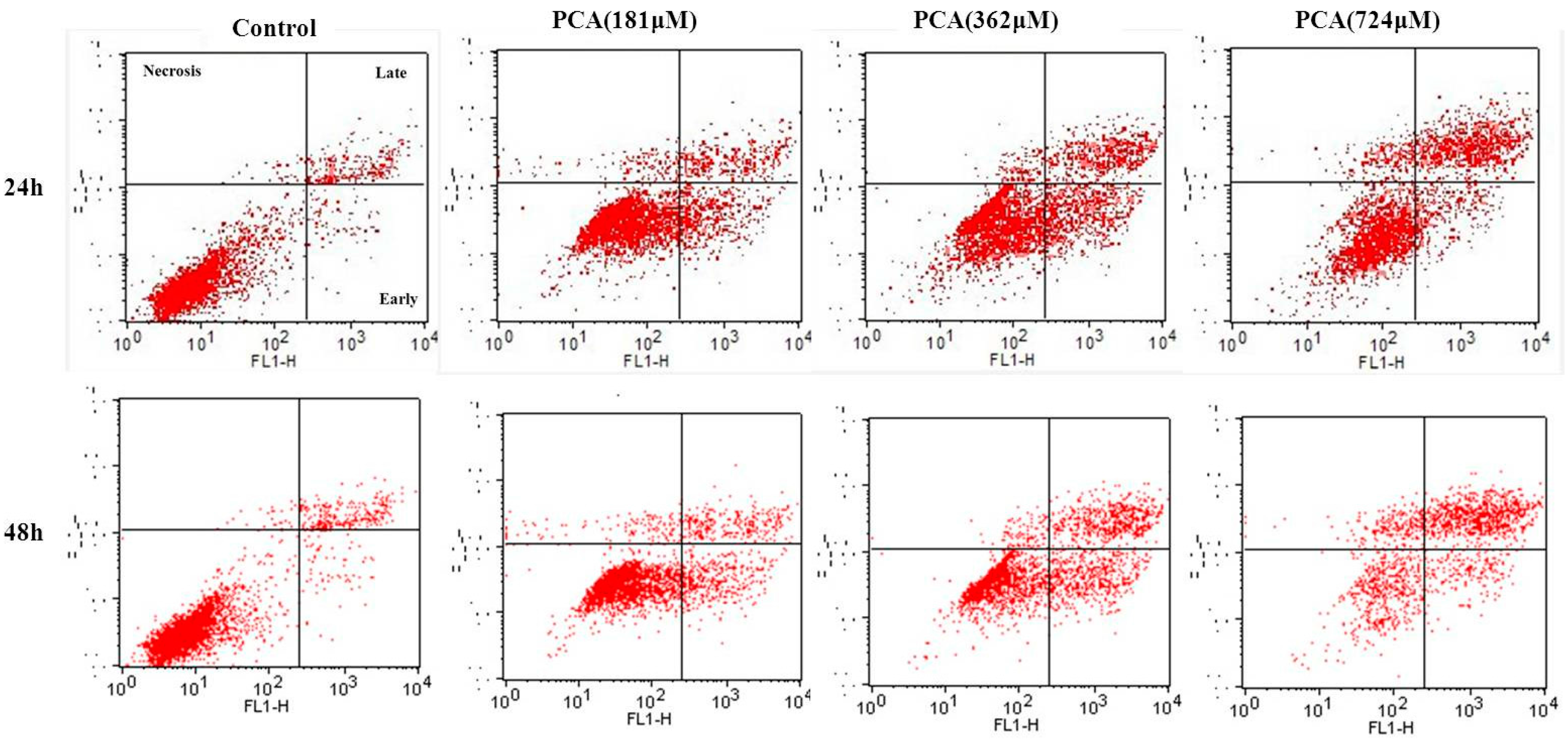
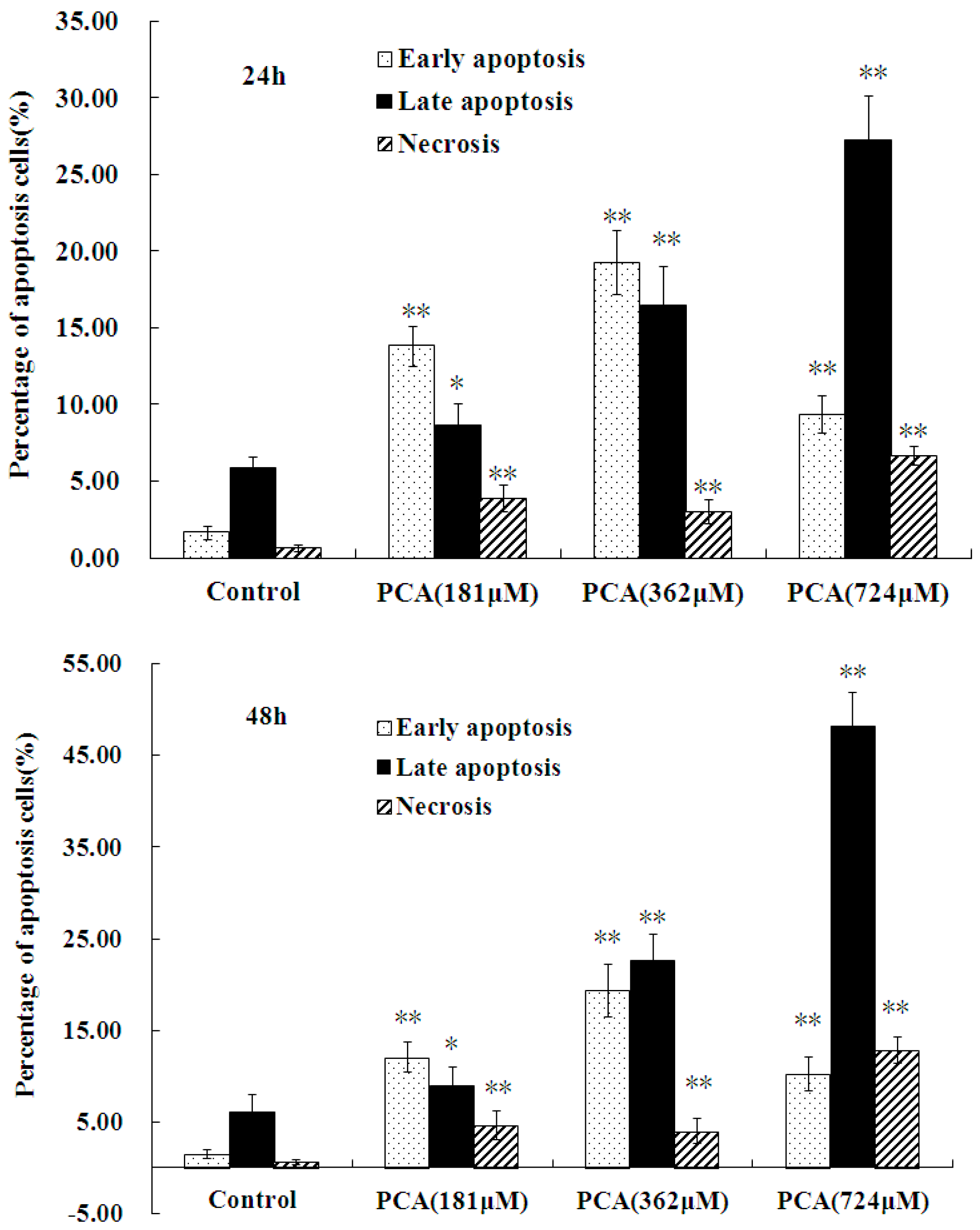
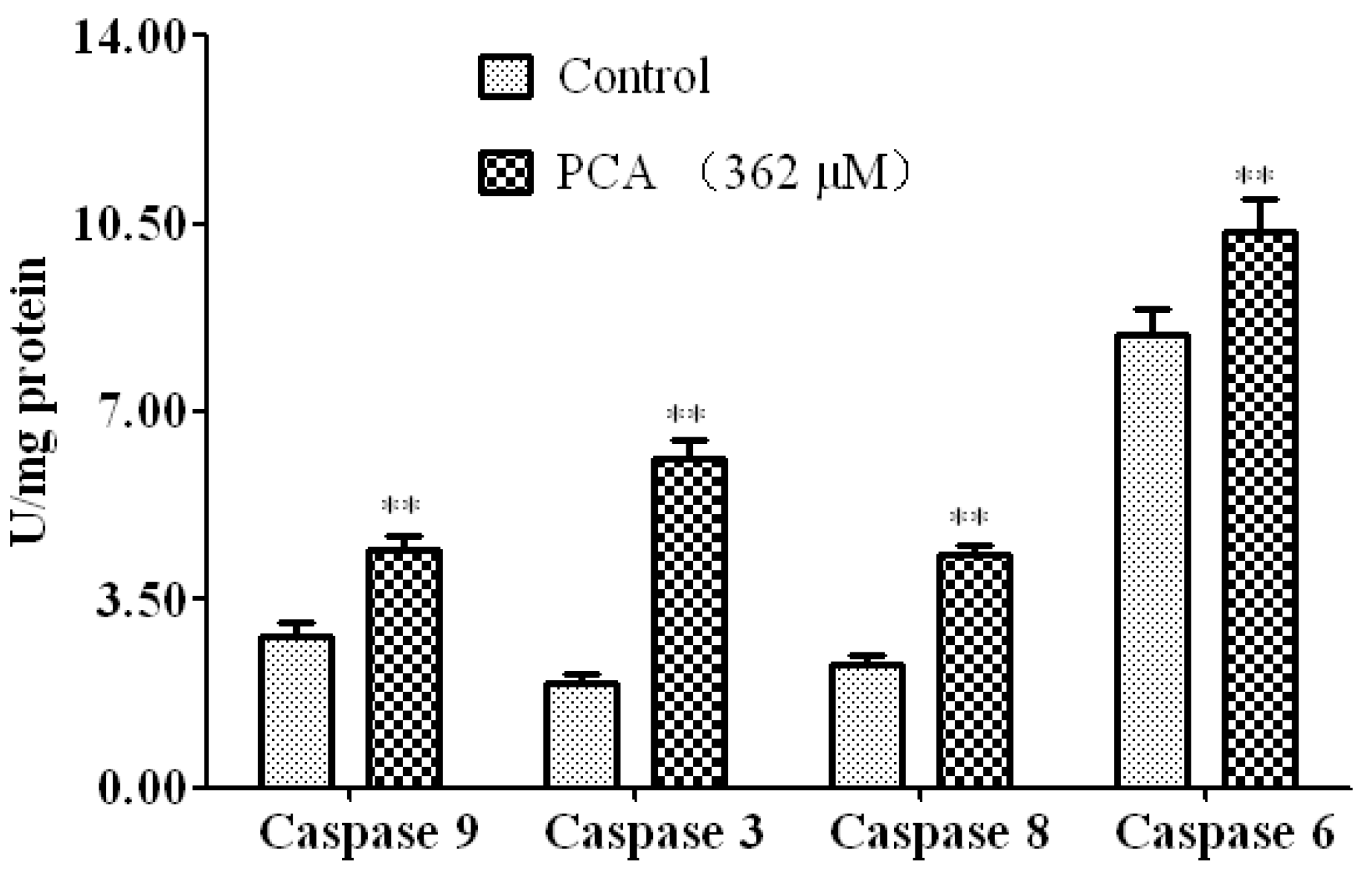
2.3. PCA-Induced Apoptosis in HT-29 Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
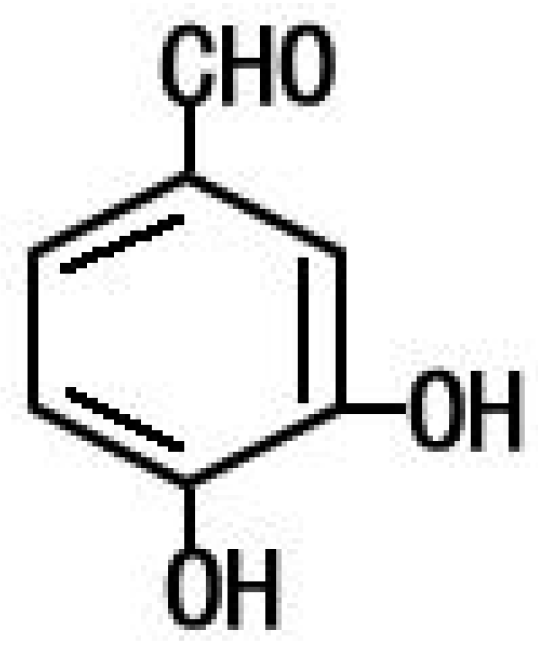
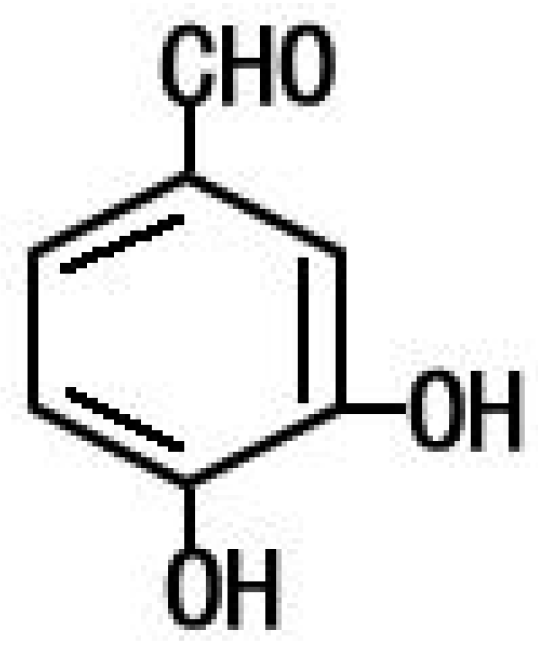
4.2. PCA from Phellinus gilvus
4.3. Cell Proliferation Assay
4.4. Cell Cycle and Apoptosis Assay
4.5. Quantitative Real-Time Reverse Transcription-PCR Analysis
4.6. Protein Expression and Western Blot Analyses
4.7. Caspase Activity Assay
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, G.; Kim, D.H.; Kim, T.D.; Park, B.J.; Park, H.D.; Park, J.I.; Na, M.K.; Kim, H.C.; Hong, N.D.; Lim, K.; et al. Protein-bound polysaccharide from Phellinus linteus induces G2/M phase arrest and apoptosis in SW480 human colon cancer cells. Cancer Lett. 2004, 216, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Zhu, J.X.; Chen, S.; Hu, G.Y. Anti-tumor Effects of Proteoglycan from Phellinus linteus by Immunomodulating and Inhibiting Reg IV/EGFR/Akt Signaling Pathway in Colorectal Carcinoma. Int. J. Biol. Macromol. 2011, 48, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Liu, P.G.; Lv, Z.Q.; Zhu, J.X.; Chen, J.E.; Chen, H.P. Polysaccharide from Phellinus linteus induces S-phase arrest in HepG2 cells by decreasing calreticulin expression and activating the p27KIP1-cyclin A/D1/E-CDK2 pathway. J. Ethnopharmacol. 2013, 150, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Liaw, C.C.; Pan, S.Z.; Yang, H.C.; Ng, L.T. Phellinus linteus polysaccharides and their immunomodulatory properties in human monocytic cells. J. Funct. Foods 2013, 5, 679–688. [Google Scholar] [CrossRef]
- Guo, J.; Zhu, T.; Collins, L.; Xiao, Z.X.; Kim, S.H. Modulation of Lung cancer growth arrest and apoptosis by Phellinus linteus. Mol. Carcinog. 2007, 46, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Zhu, T.; Guo, J.; Collins, L.; Kelly, J.; Xiao, Z.J.; Kim, S.H. Phellinus linteus activates different pathways to induce apoptosis in prostate cancer cells. Br. J. Cancer 2007, 96, 583–590. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Song, Y.S.; Kim, S.K.; Kim, B.C.; Lim, C.J.; Park, E.H. Anti-inflammatory and related pharmacological activities of the n-BuOH subfraction of mushroom Phellinus linteus. J. Ethnopharmacol. 2004, 93, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Zhong, S.; Ji, D.F. Comparison of bioactive substances between wild and cultivated fresh-fruiting bodies of Phellinus linteus. Acta Sericol. Sin. 2016, in press. [Google Scholar]
- Kim, K.J.; Kim, M.A.; Jung, J.H. Antitumor and antioxidant activity of protocatechualdehyde produced from Streptomyces lincolnensis M-20. Arch. Pharm. Res. 2008, 31, 1572–1577. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Yoon, S.H.; Kim, Y.S.; Kim, S.K.; Moon, B.J.; Bae, Y.S. Apoptotic cell death through inhibition of protein kinase CKII activity by 3,4-dihydroxybenzaldehyde purified from Xanthium strumarium. Nat. Prod. Res. 2008, 22, 1441–1450. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.B.; Lee, S.H. Protocatechualdehyde possesses anti-cancer activity through downregulating cyclin D1 and HDAC2 in human colorectal cancer cells. Biochem. Biophys. Res. Commun. 2013, 430, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.R.; Lee, M.H.; Eo, H.J.; Park, G.H.; Song, H.M.; Kim, M.K.; Lee, J.W.; Jeong, J.B. The contribution of activating transcription factor 3 to apoptosis of human colorectal cancer cells by protocatechualdehyde, a naturally occurring phenolic compound. Arch. Biochem. Biophys. 2014, 564, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Takai, N.; Ueda, T.; Nasu, K.; Yamashita, S.; Toyofuku, M. Targeting calcium/calmodulin-dependence kinase I and II as a potential anti-proliferation remedy for endometrial carcinomas. Cancer Lett. 2009, 277, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Murray, A.W. Recycling the cell cycle: Cyclins revisited. Cell 2004, 116, 221–234. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, A.G.; Xia, T.; He, P. Effects of fluoride on DNA damage, S-phase cell-cycle arrest and the expression of NF-κB in primary cultured rat hippocampal neurons. Toxicol. Lett. 2008, 179. [Google Scholar] [CrossRef] [PubMed]
- Gamet-Payrastre, L.; Li, P.; Lumeau, S.; Cassar, G.; Dupont, M.-A.; Chevolleau, S.; Gasc, N.; Tulliez, J.; Tercé, F. Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 2000, 60, 1426–1433. [Google Scholar] [PubMed]
- Supriatno, K.H.; Mohammad, O.H.; Takashi, B.; Hideo, Y.; Mitsunobu, S. Overexpression of p27KIP1 induces growth arrest and apoptosis in an oral cancer cell line. Oral Oncol. 2002, 38, 730–736. [Google Scholar] [CrossRef]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Renault, T.T.; Chipuk, J.E. Getting away with murder: How does the BCL-2 family of proteins kill with immunity? Ann. N. Y. Acad. Sci. 2013, 1285, 59–79. [Google Scholar] [CrossRef] [PubMed]
- Etoh, H.; Murakami, K.; Yogoh, T.; Ishikawa, H.; Fukuyama, Y.; Tanaka, H. Antioxidative compounds in barley tea. Biosci. Biotechnol. Biochem. 2004, 68, 2616–2618. [Google Scholar] [CrossRef] [PubMed]
- Mulvena, D.; Webb, E.C.; Zerner, B. 3,4-Dihydroxybenzaldehyde a fungistatic substance from green Cavendish bananas. Phytochemistry 1969, 8, 393–395. [Google Scholar] [CrossRef]
- Weber, B.; Hoesch, L.; Rast, D.M. Protocatechualdehyde and other phenols as cell-wall components of grapevine leaves. Phytochemistry 1995, 40, 433–437. [Google Scholar] [CrossRef]
- OuYang, F.U.; Wang, G.B.; Guo, W.; Zhang, Y.Y.; Xiang, W.H.; Zhao, M. AKT signalling and mitochondrial pathways are involved in mushroom polysaccharide-induced apoptosis and G1 or S phase arrest in human hepatoma cells. Food Chem. 2013, 138, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Draetta, G.F. Cell cycle: Will the real Cdk-activating kinase please stand up. Curr. Biol. 1997, 7, 50–52. [Google Scholar] [CrossRef]
- Moon, J.; Yu, S.J.; Kim, H.S.; Sohn, J. Induction of G1 Cell Cycle Arrest and p27KIP1 Increase by Panaxydol Isolated from Panax ginseng. Biochem. Pharmacol. 2000, 59, 1109–1116. [Google Scholar] [CrossRef]
- Tu, Y.S.; Kang, X.L.; Zhou, J.G.; Lv, X.F.; Tang, Y.B.; Guan, Y.Y. Involvement of Chk1-Cdc25A-cyclin A/CDk2 pathway in simvastatin induced S-phase cell cycle arrest and apoptosis in multiple myeloma cells. Eur. J. Pharmacol. 2011, 670, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.M.; Cory, S. The Bcl-2 apoptotic switch in cancer development and therapy. Oncogene 2007, 26, 1324–1337. [Google Scholar] [CrossRef] [PubMed]
- Polyak, K.; Lee, M.H.; Erdjument-Bromage, H.; Koff, A.; Roberts, J.M.; Tempst, P.; Massagué, J. Cloning of p27KIP1, a cyclin-dependent kinase inhibitor and a potential mediator of extracellular antimitogenic signals. Cell 1994, 78, 59–66. [Google Scholar] [CrossRef]
- Krude, T.; Jackman, M.; Pinesm, J.; Laskey, R.A. Cyclin/Cdk-dependent initiation of DNA replication in a human cell-free system. Cell 1997, 188, 109–119. [Google Scholar] [CrossRef]
- Ding, H.; Han, C.; Guo, D.; Wang, D.; Chen, C.S.; Ambrosio, S.M.D. OSU03012 activates Erk1/2 and Cdks leading to the accumulation of cells in the S-phase and apoptosis. Int. J. Cancer 2008, 123, 2923–2930. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.Y.; Chang, G.C.; Chen, K.C.; Hung, H.W.; Hsu, K.H.; Sheu, G.T.; Hsu, S.L. Sustained activation of ERK and Cdk2/cyclin-A signaling pathway by pemetrexed leading to S-phase arrest and apoptosis in human non-small cell lung cancer A549 cells. Eur. J. Pharmacol. 2011, 663, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Ito, H.; Tamamori-Adachi, M.; Ono, Y.; Nozato, T.; Abe, S. Cyclin A/cdk2 activation is involved in hypoxia-induced apoptosis in cardiomyocytes. Circ. Res. 2001, 88, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, E.J. Selective inhibitors of Bcl-2 and Bcl-xL: Balancing antitumor activity with on-target toxicity, Bioorg. Med. Chem. Lett. 2016, 9, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Woodhouse, E.C.; Chuaqui, R.F.; Liotta, L.A. General mechanisms of metastasis. Cancer 1997, 80, 1529–1537. [Google Scholar] [CrossRef]
- Lamkanfi, M.; Festjens, N.; Declercq, W.; Vanden Berghe, T.; Vandenabeele, P. Caspases in cell survival, proliferation and differentiation. Cell Death Differ. 2007, 1, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Hajiaghaalipour, F.; Kanthimathi, M.S.; Sanusi, J.; Rajarajeswaran, J. White tea (Camellia sinensis) inhibits proliferation of the colon cancer cell line, HT-29, activates caspases and protects DNA of normal cells against oxidative damage. Food Chem. 2015, 169, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Ji, D.F.; Li, Y.G.; Lin, T.B.; Lv, Z.Q.; Chen, H.P. Activation of p27KIP1-cyclin D1/E-CDK2 pathway by polysaccharide from Phellinus linteus leads to S-phase arrest in HT-29 cells. Chem. Biol. Interact. 2013, 206, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.G.; Ji, D.F.; Zhong, S.; Lin, T.B.; Lv, Z.Q.; Hu, G.Y.; Wang, X. 1-deoxynojirimycin inhibits glucose absorption and accelerates glucose metabolism in streptozotocin-induced diabetic mice. Sci. Rep. 2013, 3. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5’-3’ | PCR Product Size (bp) |
|---|---|---|
| Bcl-2-F | GCCTTCTTTGAGTTCGGTG | 91 |
| Bcl-2-R | AGTCATCCACAGGGCGAT | |
| Bcl-xL-F | CCAGAAGGGACTGAATCGGA | 117 |
| Bcl-xL-R | GCATCCAAACTGCTGCTGT | |
| Bid-F | ATCCGTGCTGTCTCCTTTG | 91 |
| Bid-R | AAACTTCCGATGGGACCAA | |
| Bak-F | AACCGACGCTATGACTCAGA | 111 |
| Bak-R | GCCACTCTCAAACAGGCTG | |
| Bax-F | ATGGGCTGGACATTGGACTT | 124 |
| Bax-R | GCCACAAAGATGGTCACGGT | |
| GAPDH-F | GGTGGTCTCCTCTGACTTCAACA | 135 |
| GAPDH-R | GTTGCTGTAGCCAAATTCGTTGT |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, S.; Li, Y.-G.; Ji, D.-F.; Lin, T.-B.; Lv, Z.-Q. Protocatechualdehyde Induces S-Phase Arrest and Apoptosis by Stimulating the p27KIP1-Cyclin A/D1-CDK2 and Mitochondrial Apoptotic Pathways in HT-29 Cells. Molecules 2016, 21, 934. https://doi.org/10.3390/molecules21070934
Zhong S, Li Y-G, Ji D-F, Lin T-B, Lv Z-Q. Protocatechualdehyde Induces S-Phase Arrest and Apoptosis by Stimulating the p27KIP1-Cyclin A/D1-CDK2 and Mitochondrial Apoptotic Pathways in HT-29 Cells. Molecules. 2016; 21(7):934. https://doi.org/10.3390/molecules21070934
Chicago/Turabian StyleZhong, Shi, You-Gui Li, Dong-Feng Ji, Tian-Bao Lin, and Zhi-Qiang Lv. 2016. "Protocatechualdehyde Induces S-Phase Arrest and Apoptosis by Stimulating the p27KIP1-Cyclin A/D1-CDK2 and Mitochondrial Apoptotic Pathways in HT-29 Cells" Molecules 21, no. 7: 934. https://doi.org/10.3390/molecules21070934