
Two Trichothecene Mycotoxins from Myrothecium roridum Induce Apoptosis of HepG-2 Cells via Caspase Activation and Disruption of Mitochondrial Membrane Potential

Abstract
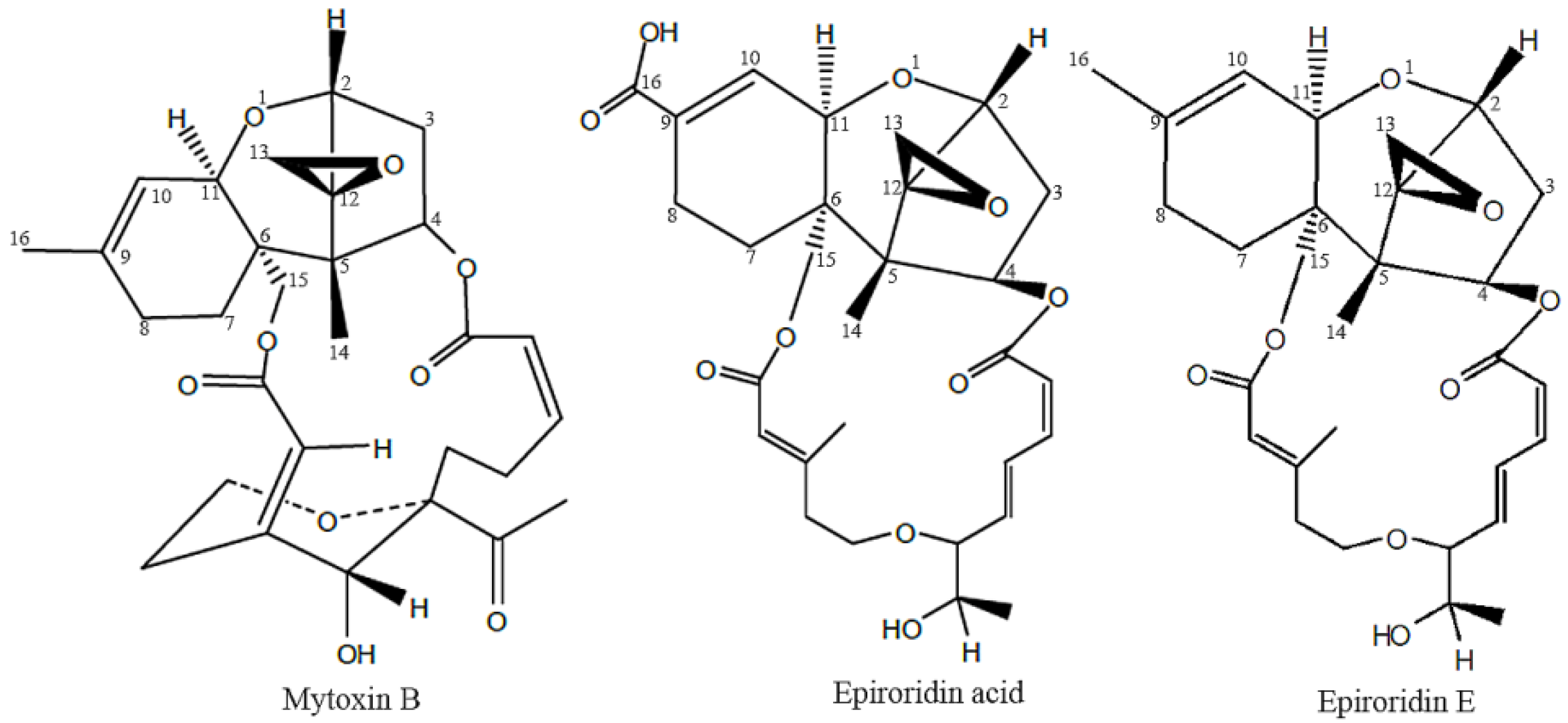
:1. Introduction
2. Results
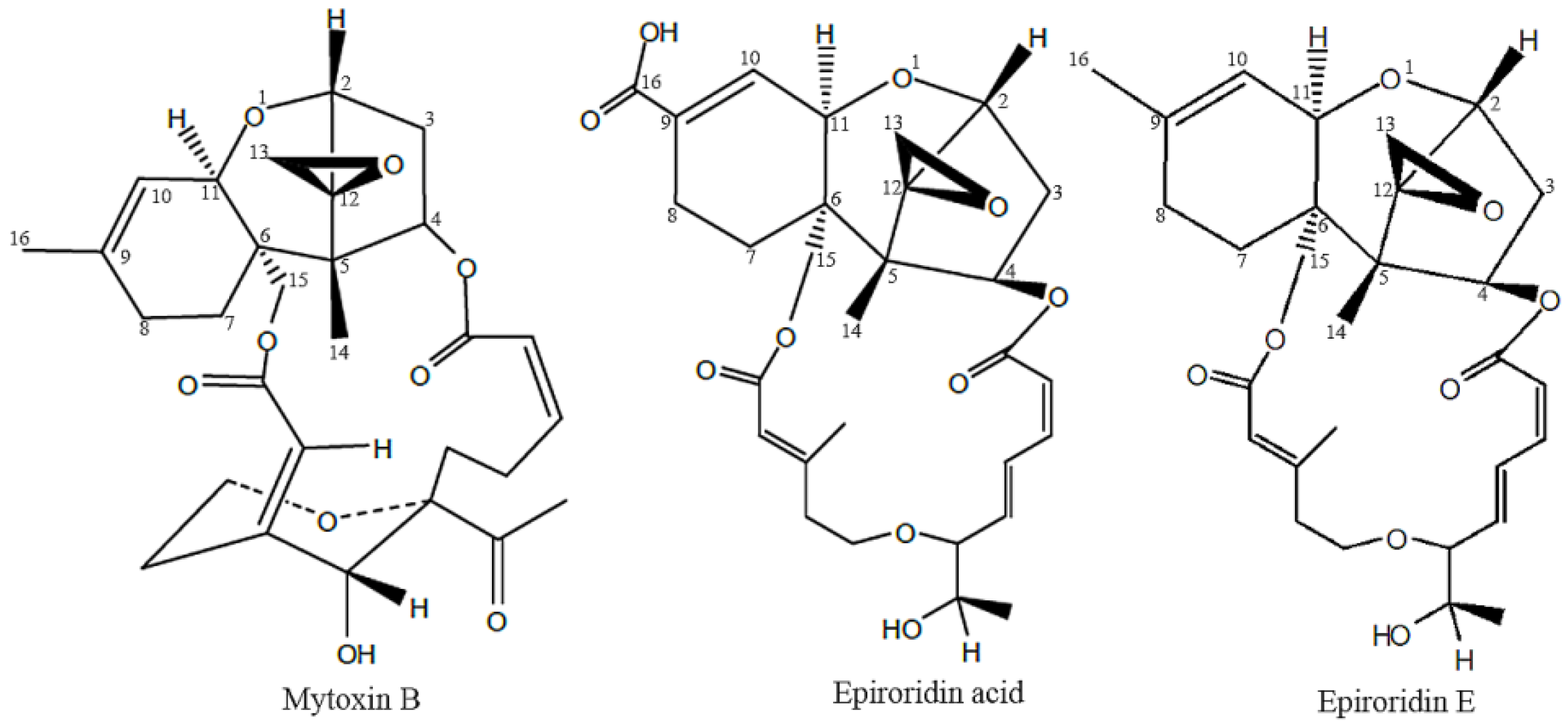
2.1. Cytotoxicity Detection of Epiroridin Acid and Mytoxin B
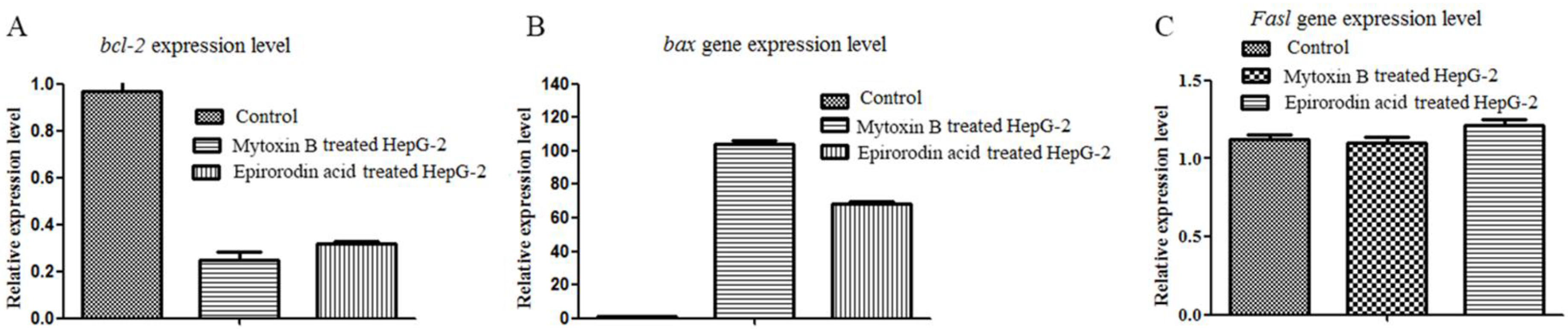
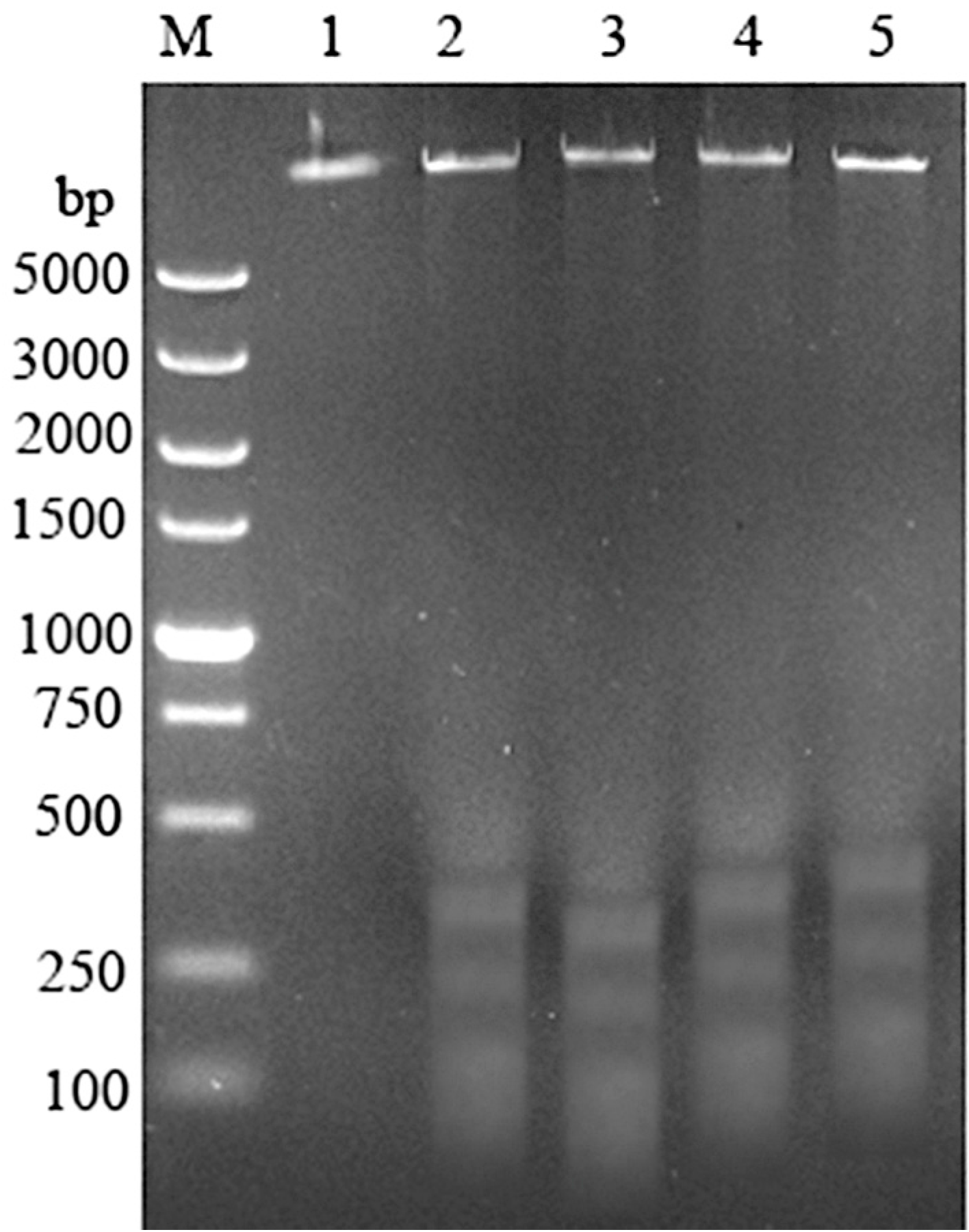
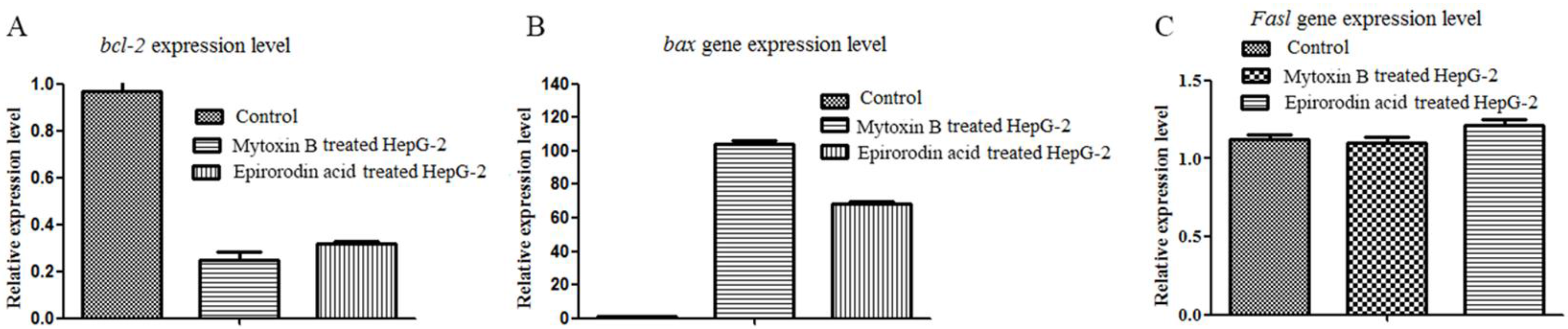
2.2. qRT-PCR Analysis of Mytoxin B- and Epiroridin Acid-treated HepG-2 Cells
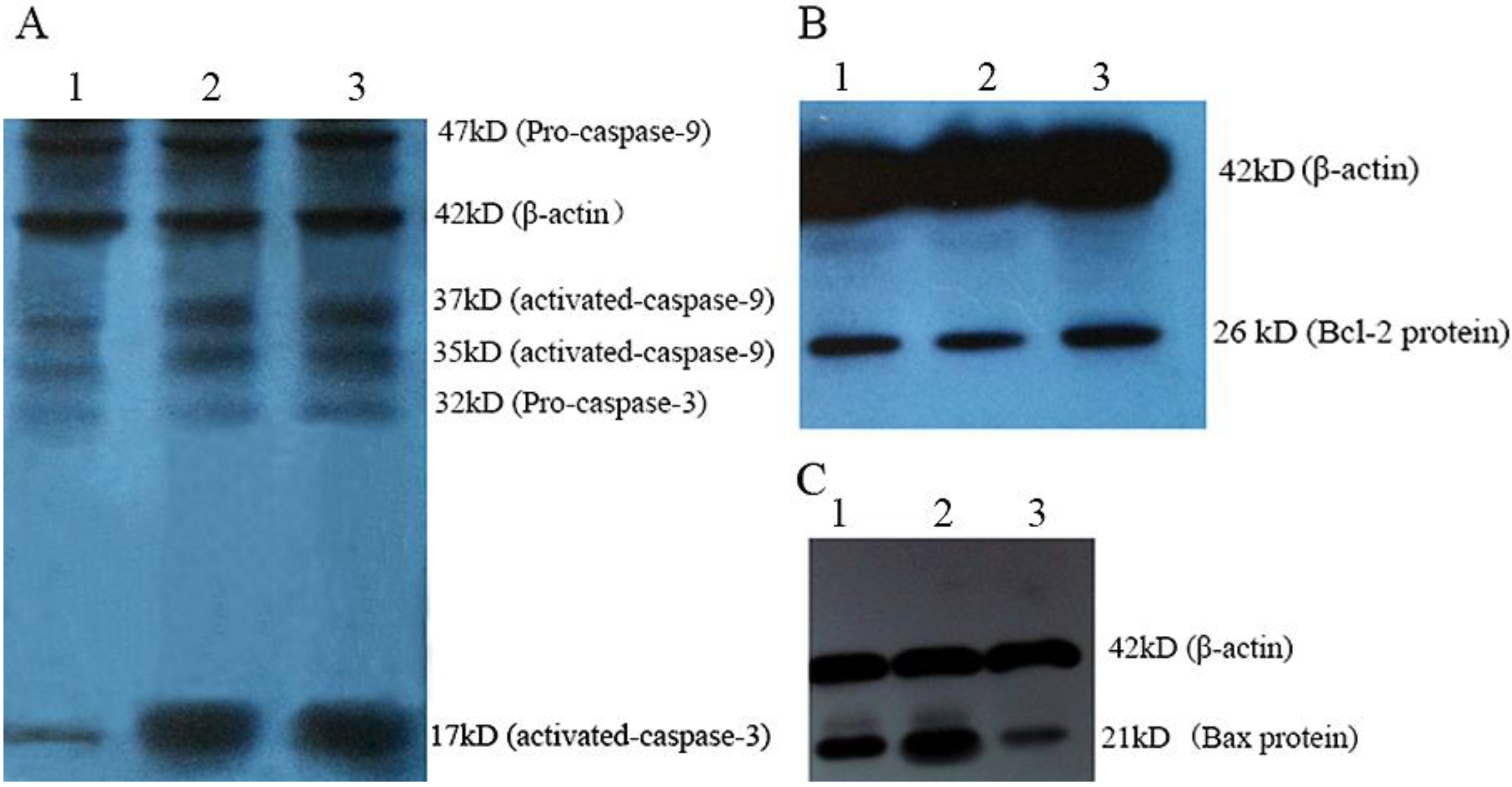
2.3. Western Blot Analysis of Mytoxin B- and Epiroridin Acid-treated HepG-2 Cells
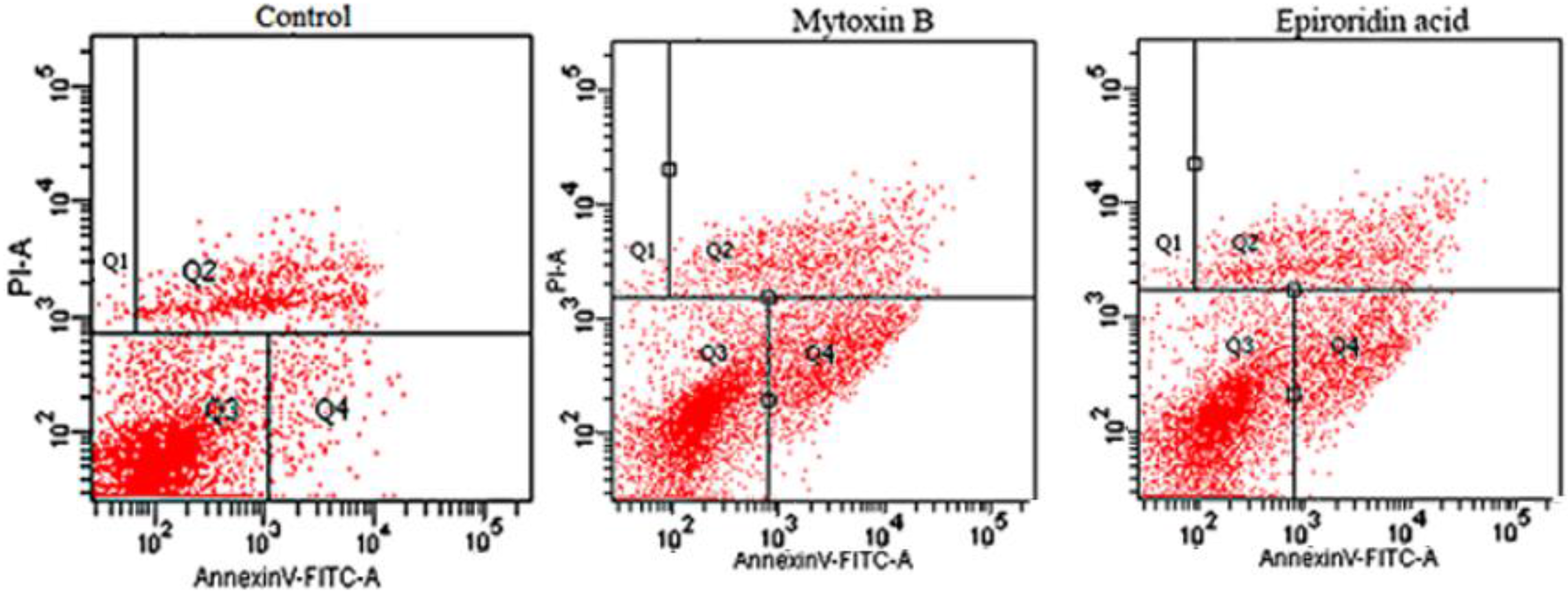
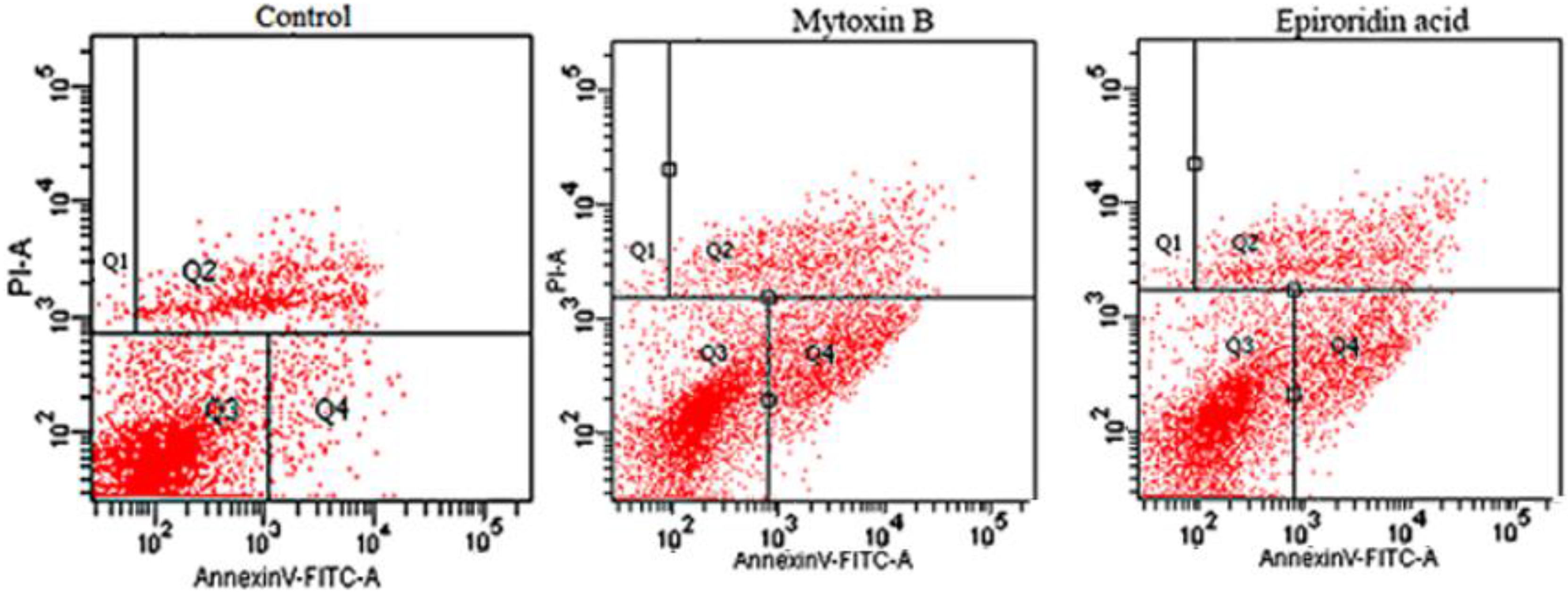
2.4. Flow Cytometry Assay of HepG-2 Cells Treated With Epororidin Acid and Mytoxin B
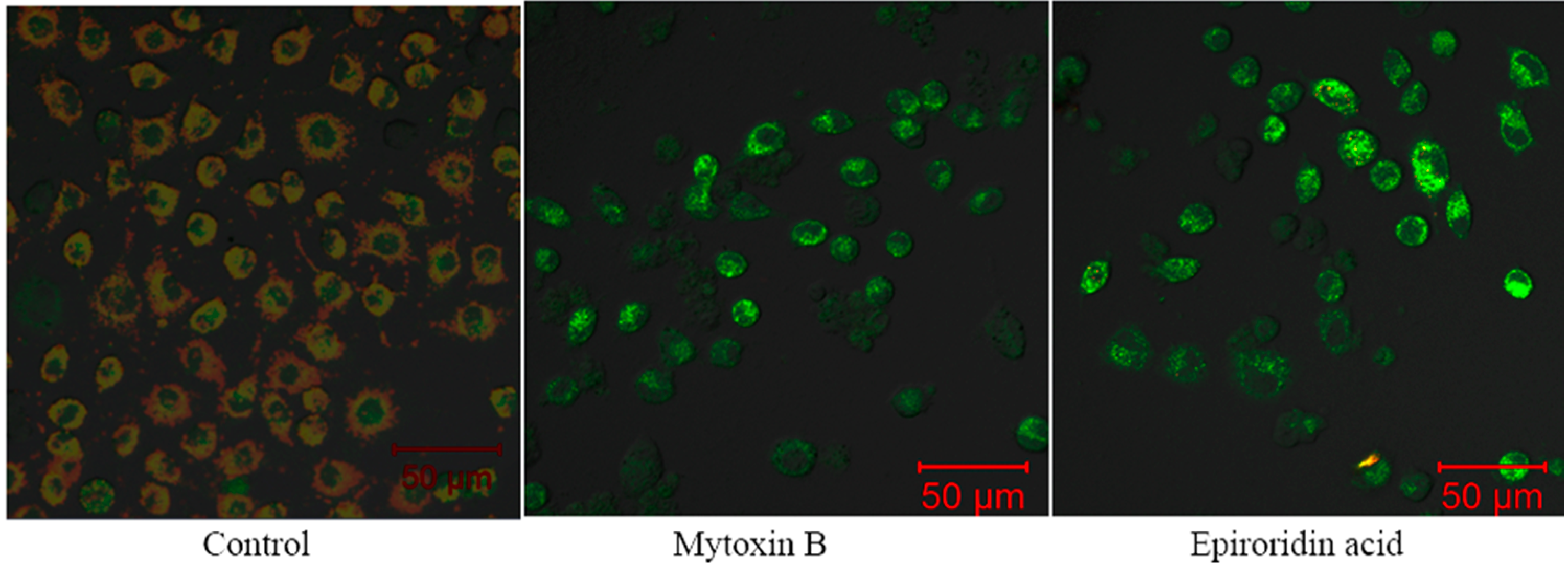
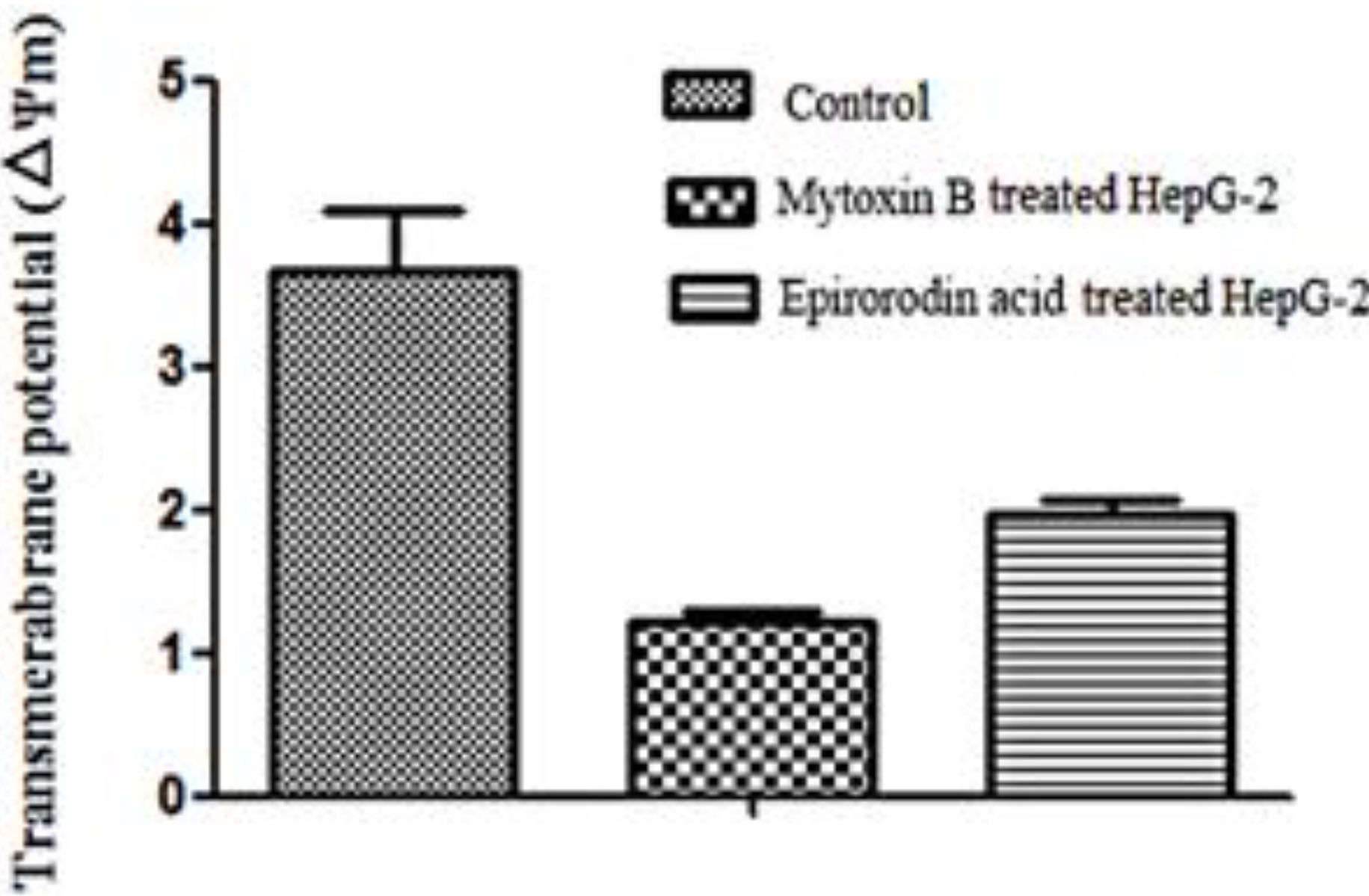
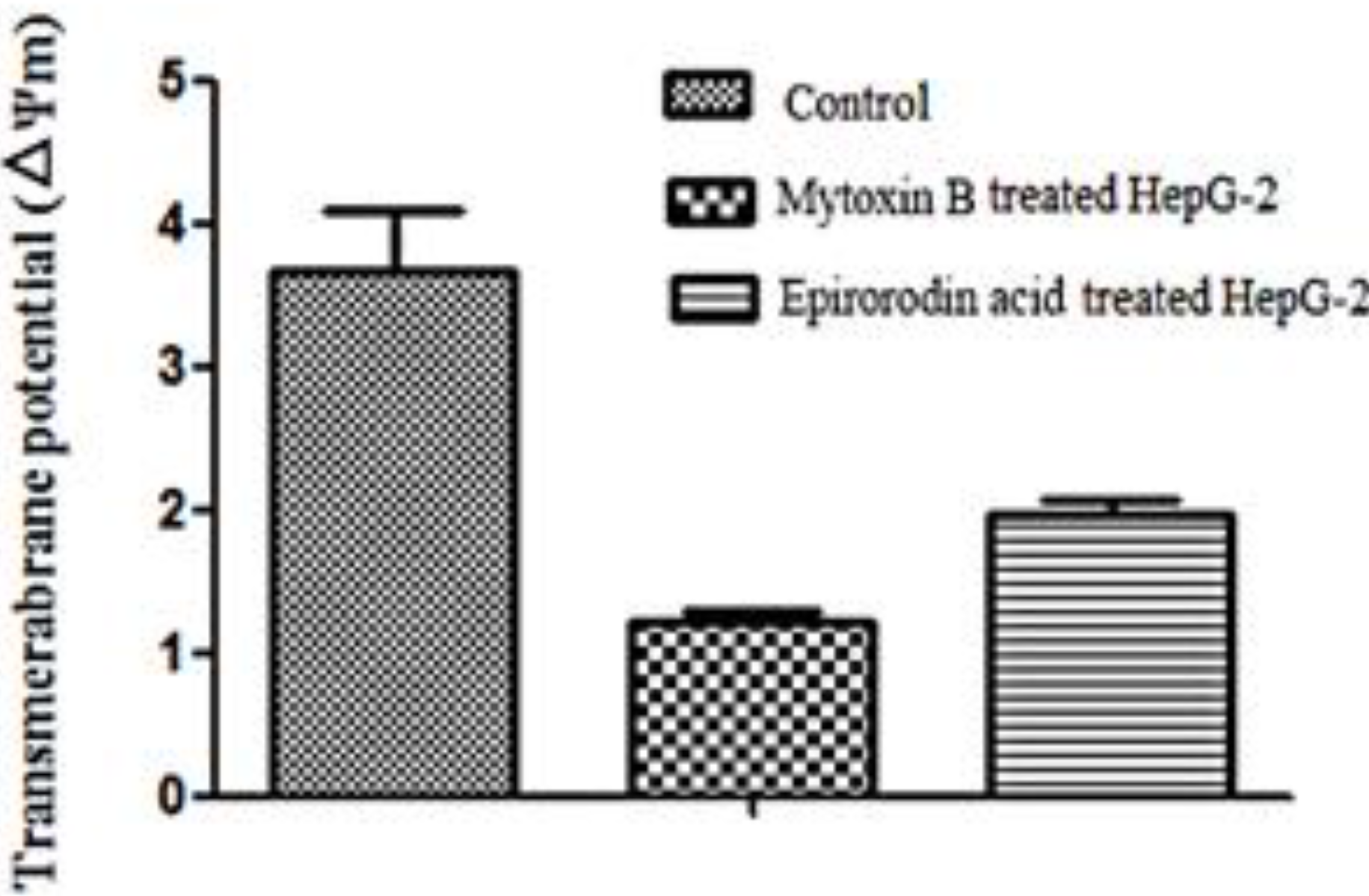
2.5. Detection of Mitochondrial Transmembrane Potential
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cytotoxicity Detection of Epiroridin Acid and Mytoxin B
4.3. qRT-PCR Analysis of Genes Related to the Apoptosis
4.4. Western Blot Analysis of Caspase-3, Caspase-9 Protein
4.5. Flow Cytometry Assay of Trichothecene Mycotoxin-treated HepG-2 Cells
4.6. Detection of Mitochondrial Transmembrane Potential
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Veiseh, M.; Gabikian, P.; Bahrami, S.-B.; Veiseh, O.; Zhang, M.; Hackman, R.C.; Ravanpay, A.C.; Stroud, M.R.; Kusuma, Y.; Hansen, S.J. Tumor paint: A chlorotoxin: Cy5.5 bioconjugate for intraoperative visualization of cancer foci. Cancer Res. 2007, 67, 6882–6888. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.-F.; Chiu, H.-C.; Chen, K.-C.; Chang, C.-H.; Chou, E. C.-L. Botulinum toxin A for the Treatment of Overactive Bladder. Toxins 2016, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Mergel, K.; Weng, A.; von Mallinckrodt, B.; Gilabert-Oriol, R.; Dürkop, H.; Melzig, M.F.; Fuchs, H. Targeted tumor therapy by epidermal growth factor appended toxin and purified saponin: An evaluation of toxicity and therapeutic potential in syngeneic tumor bearing mice. Mol. Oncol. 2013, 7, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Vyas, V.K.; Brahmbhatt, K.; Bhatt, H.; Parmar, U. Therapeutic potential of snake venom in cancer therapy: Current perspectives. Asian Pac. J. Trop. biomed. 2013, 3, 156–162. [Google Scholar] [CrossRef]
- Erin, K.W.; Sangeetha, R.; Garrett, L.R.; Brigitte, B.; Jennifer, E.; William, N.; Jack, P.H. A novel targeted engineered toxin body for treatment of HER2 positive breast cancer. Cancer Res. 2015, 75. [Google Scholar] [CrossRef]
- Berstad, M.; Cheung, L.; Berg, K.; Peng, Q.; Fremstedal, A.; Patzke, S.; Rosenblum, M.; Weyergang, A. Design of an EGFR-targeting toxin for photochemical delivery: In vitro and in vivo selectivity and efficacy. Oncogene 2015, 34, 5582–5592. [Google Scholar] [CrossRef] [PubMed]
- Koshikawa, N.; Takenaga, K. HyPoxia-regulated expression of attenuated diphiheria toxinA fused with hypoxia-inducible factor-lα oxygen-dependent degradation domain preferentially induces apoptosis of hypoxic cells in solid tumor. Cancer Res. 2005, 65, 11622–11630. [Google Scholar] [CrossRef] [PubMed]
- Steinmetz, W.E.; Robustelli, P.; Edens, E.; Heineman, D. Structure and conformational dynamics of trichothecene mycotoxins. J. Nat. Prod. 2008, 71, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.J.; Bielawski, J.P.; Kistler, H.C.; Sullivan, E.; O′Donnell, K. Ancestral polymorphism and adaptive evolution in the trichothecene mycotoxin gene cluster of phytopathogenic Fusarium. Proc. Nat. Acad. Sci. USA 2002, 99, 9278–9283. [Google Scholar] [CrossRef] [PubMed]
- Ueno, Y.; Umemori, K.; Niimi, E.C.; Tanuma, S.I.; Nagata, S.; Sugamata, M.; Ihara, T.; Sekijima, M.; Kawai, K.I.; Ueno, I. Induction of apoptosis by T-2 toxin and other natural toxins in HL-60 human promyelotic leukemia cells. Nat. Toxins 1995, 3, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Frankič, T.; Pajk, T.; Rezar, V.; Levart, A.; Salobir, J. The role of dietary nucleotides in reduction of DNA damage induced by T-2 toxin and deoxynivalenol in chicken leukocytes. Food Chem. Toxicol. 2006, 44, 1838–1844. [Google Scholar] [CrossRef] [PubMed]
- Cetin, Y.; Bullerman, L.B. Cytotoxicity of Fusarium mycotoxins to mammalian cell cultures as determined by the MTT bioassay. Food Chem. Toxicol. 2005, 43, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Liu, W.Z.; Chen, Y.C.; Sun, Z.H.; Tan, Y.Z.; Li, H.H.; Zhang, W.M. Cytotoxic trichothecene macrolides from the endophyte fungus Myrothecium roridum. J. Asian Nat. Prod. Res. 2016, 1–6. [Google Scholar] [CrossRef]
- Anderson, D.; Black, R.; Lee, C.; Pottage, C.; Rickard, R.; Sandford, M.; Webber, T.; Williams, N. Structure-activity studies of trichothecenes: Cytotoxicity of analogs and reaction products derived from T-2 toxin and neosolaniol. J. Med. Chem. 1989, 32, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Sun, M.; Zhu, W.; Wang, Z.; Li, Y.; Ma, J. The anti-cancer potency and mechanism of a novel tumor-activated fused toxin, DLM. Toxins 2015, 7, 423–438. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Choi, D.-K.; Jung, H.J. Neuroprotective effects of vanillyl alcohol in Gastrodia elata Blume through suppression of oxidative stress and anti-apoptotic activity in toxin-induced dopaminergic MN9D cells. Molecules 2011, 16, 5349–5361. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Narita, M.; Tsujimoto, Y. Bcl-2 family proteins regulate the release of apoptogenic cytochrome c by the mitochondrial channel VDAC. Nature 1999, 399, 483–487. [Google Scholar] [PubMed]
- Yang, J.; Liu, X.; Bhalla, K.; Kim, C.N.; Ibrado, A.M.; Cai, J.; Peng, T.-I.; Jones, D.P.; Wang, X. Prevention of apoptosis by Bcl-2: Release of cytochrome c from mitochondria blocked. Science 1997, 275, 1129–1132. [Google Scholar] [CrossRef] [PubMed]
- Pattingre, S.; Tassa, A.; Qu, X.; Garuti, R.; Liang, X.; Mizushima, N.; Packer, M.; Schneider, M.; Levine, B. Bcl-2 antiapoptotic proteins inhibit Beclin 1-dependent autophagy. Cell 2005, 122, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, L.; Hong, Z.; Zheng, H.; Li, N.; Gao, H.; Chen, B.; Zhao, Y. Methanolic extract of Pien Tze Huang induces apoptosis signaling in human osteosarcoma MG63 cells via multiple pathways. Molecules 2016, 21, 283. [Google Scholar] [CrossRef] [PubMed]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Li, G.; Mo, Z.; Xiao, P.; Li, J.; Huang, J. De Novo assembly of the Japanese Flounder (Paralichthys olivaceus) spleen transcriptome to identify putative genes involved in immunity. PLoS ONE 2015, 10, e0117642. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5’-3’) |
|---|---|
| bcl-2 F | TTGGCCCCCGTTGCTT |
| bcl-2 R | CGGTTATCGTACCCCGTTCTC |
| bax F | TCCCCCCCGAGAGGTCTTTT |
| bax R | CGGCCCCAGTTGAAGTTG |
| Fasl F | ATCCCTCTGGAATGGGAAGA |
| Fasl R | CCATATCTGTCCAGTACTGC |
| β-actin F | GGCATCGTAGTGAGCTCCG |
| β-actin R | GCTGGAAGTGAACGCAG |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, W.; Chen, Y.; Li, H.; Zhang, W.; Liu, H.; Sun, Z.; Liu, T.; Li, S. Two Trichothecene Mycotoxins from Myrothecium roridum Induce Apoptosis of HepG-2 Cells via Caspase Activation and Disruption of Mitochondrial Membrane Potential. Molecules 2016, 21, 781. https://doi.org/10.3390/molecules21060781
Ye W, Chen Y, Li H, Zhang W, Liu H, Sun Z, Liu T, Li S. Two Trichothecene Mycotoxins from Myrothecium roridum Induce Apoptosis of HepG-2 Cells via Caspase Activation and Disruption of Mitochondrial Membrane Potential. Molecules. 2016; 21(6):781. https://doi.org/10.3390/molecules21060781
Chicago/Turabian StyleYe, Wei, Yuchan Chen, Haohua Li, Weimin Zhang, Hongxin Liu, Zhanghua Sun, Taomei Liu, and Saini Li. 2016. "Two Trichothecene Mycotoxins from Myrothecium roridum Induce Apoptosis of HepG-2 Cells via Caspase Activation and Disruption of Mitochondrial Membrane Potential" Molecules 21, no. 6: 781. https://doi.org/10.3390/molecules21060781
APA StyleYe, W., Chen, Y., Li, H., Zhang, W., Liu, H., Sun, Z., Liu, T., & Li, S. (2016). Two Trichothecene Mycotoxins from Myrothecium roridum Induce Apoptosis of HepG-2 Cells via Caspase Activation and Disruption of Mitochondrial Membrane Potential. Molecules, 21(6), 781. https://doi.org/10.3390/molecules21060781