
The Protective Effects of Alisol A 24-Acetate from Alisma canaliculatum on Ovariectomy Induced Bone Loss in Vivo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Alisol A 24-Acetate from Alisma canaliculatum
2.2. Animals and Experimental Treatments
2.3. Measurements of Serum Ca, IP, and TCHO
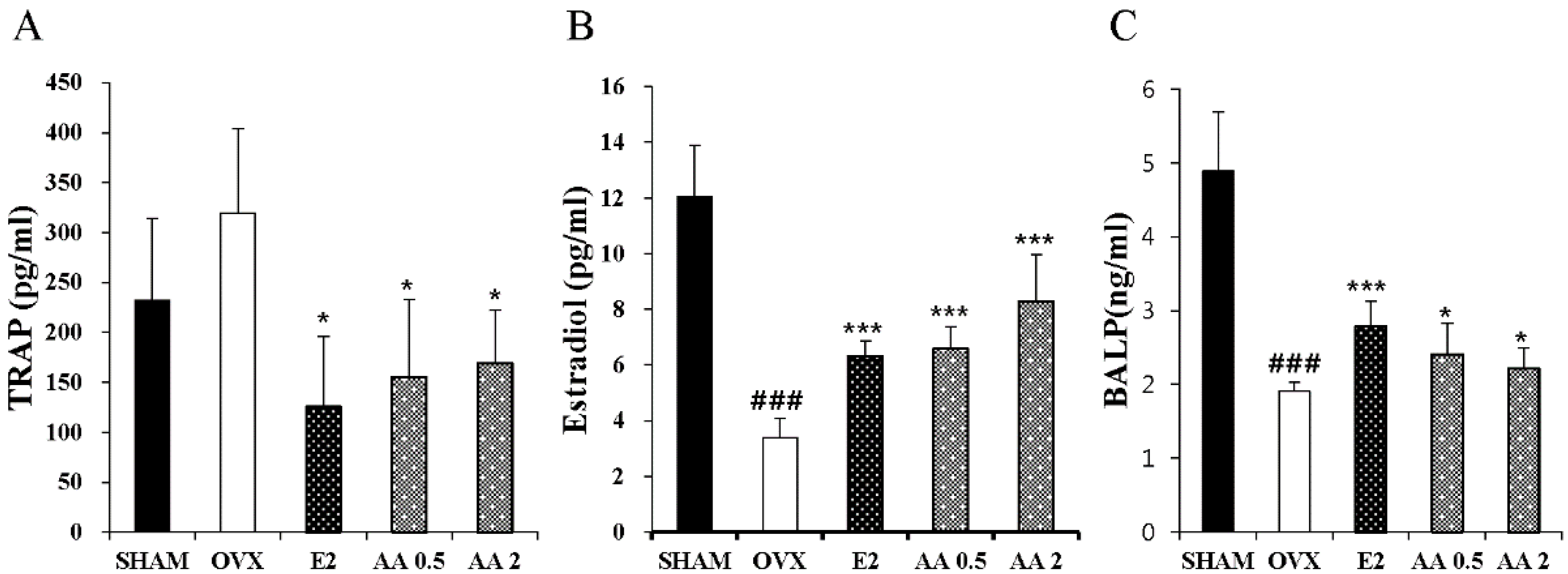
2.4. Measurements of TRAP, E2 and BALP in Serum by ELISA
2.5. Flow Cytometry
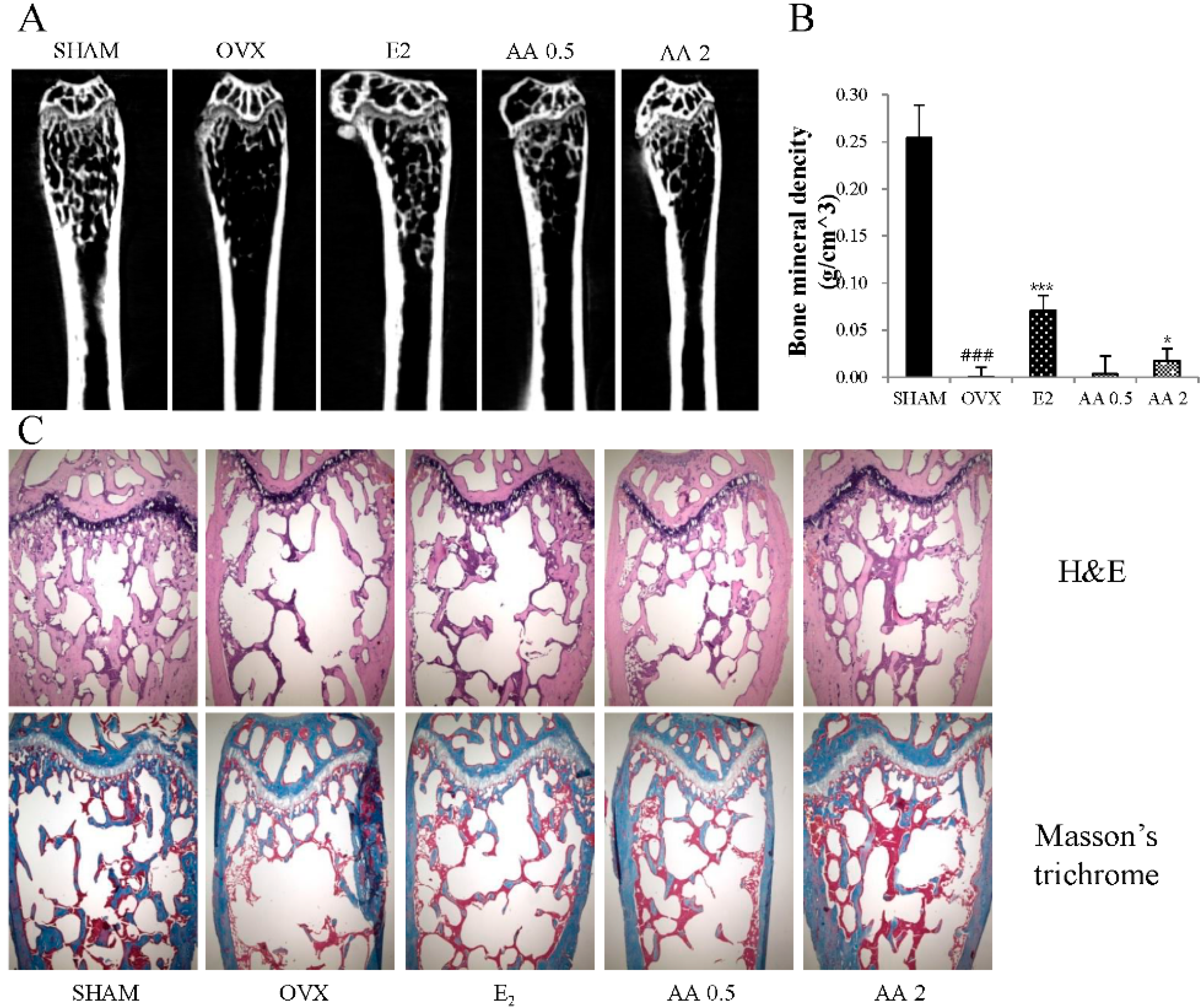
2.6. Bone Structure Analysis
2.7. Histological Analysis
2.8. Statistical Analysis
3. Results and Discussion
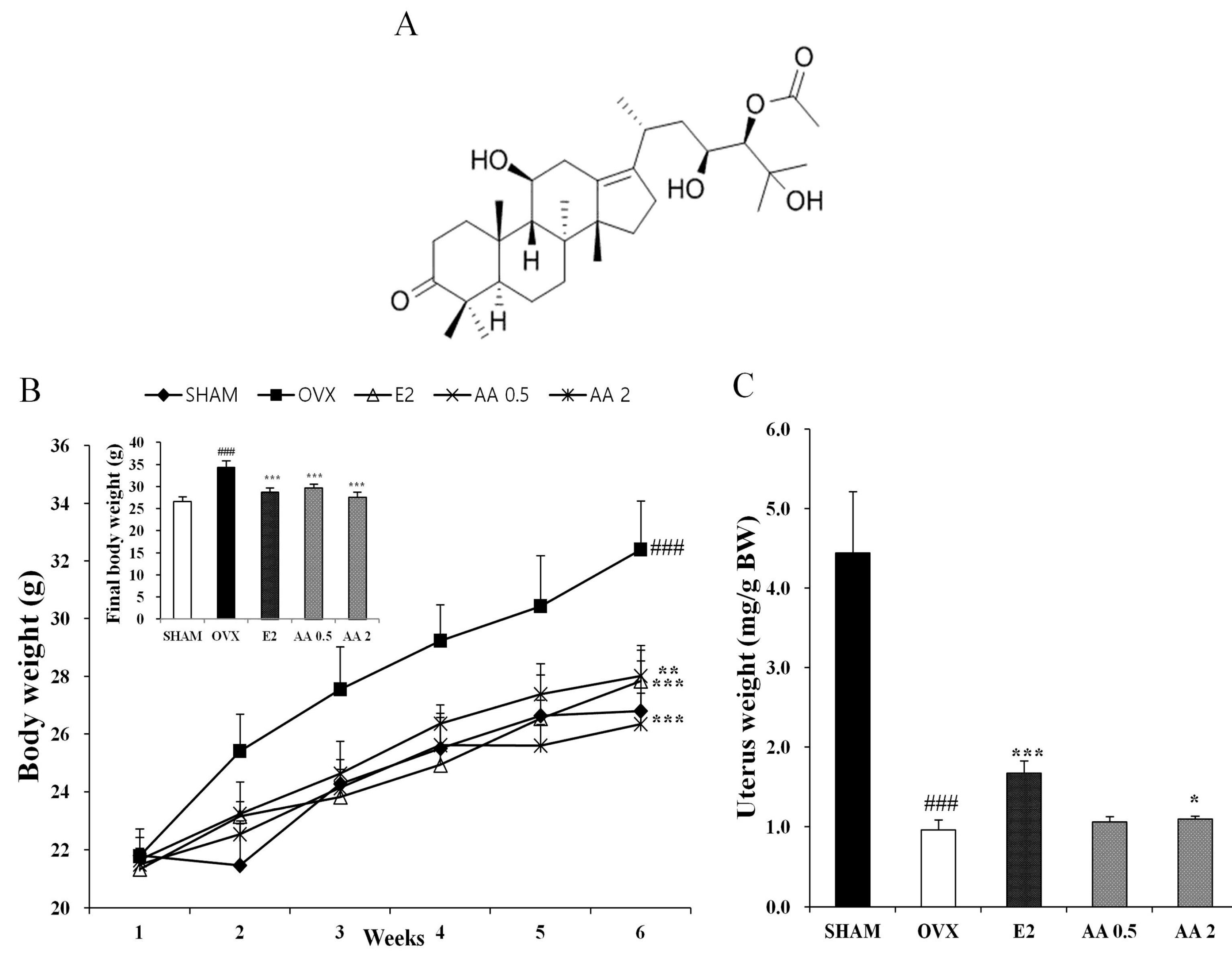
3.1. Effects of AA on Body, Uterus, and Bone Weights in OVX Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Length (mm) | Weight (mg) | |||
|---|---|---|---|---|
| Tibia | Femur | Tibia | Femur | |
| SHAM | 19.528 ± 0.606 | 15.986 ± 0.074 | 45.9 ± 3.937 | 59.68 ± 2.791 |
| OVX | 18.338 ± 0.638 # | 15.538 ± 0.349 # | 40.3 ± 4.702 | 51.48 ± 4.827 ## |
| E2 | 18.784 ± 0.071 | 16.344 ± 0.129 ** | 45.26 ± 1.024 | 57.84 ± 1.44 * |
| AA 0.5 | 18.556 ± 0.099 | 16.18 ± 0.154 ** | 43.9 ± 2.747 | 56.32 ± 2.681 |
| AA 2 | 18.592 ± 0.112 | 15.826 ± 0.119 | 47.22 ± 2.734 * | 58.84 ± 2.855 * |
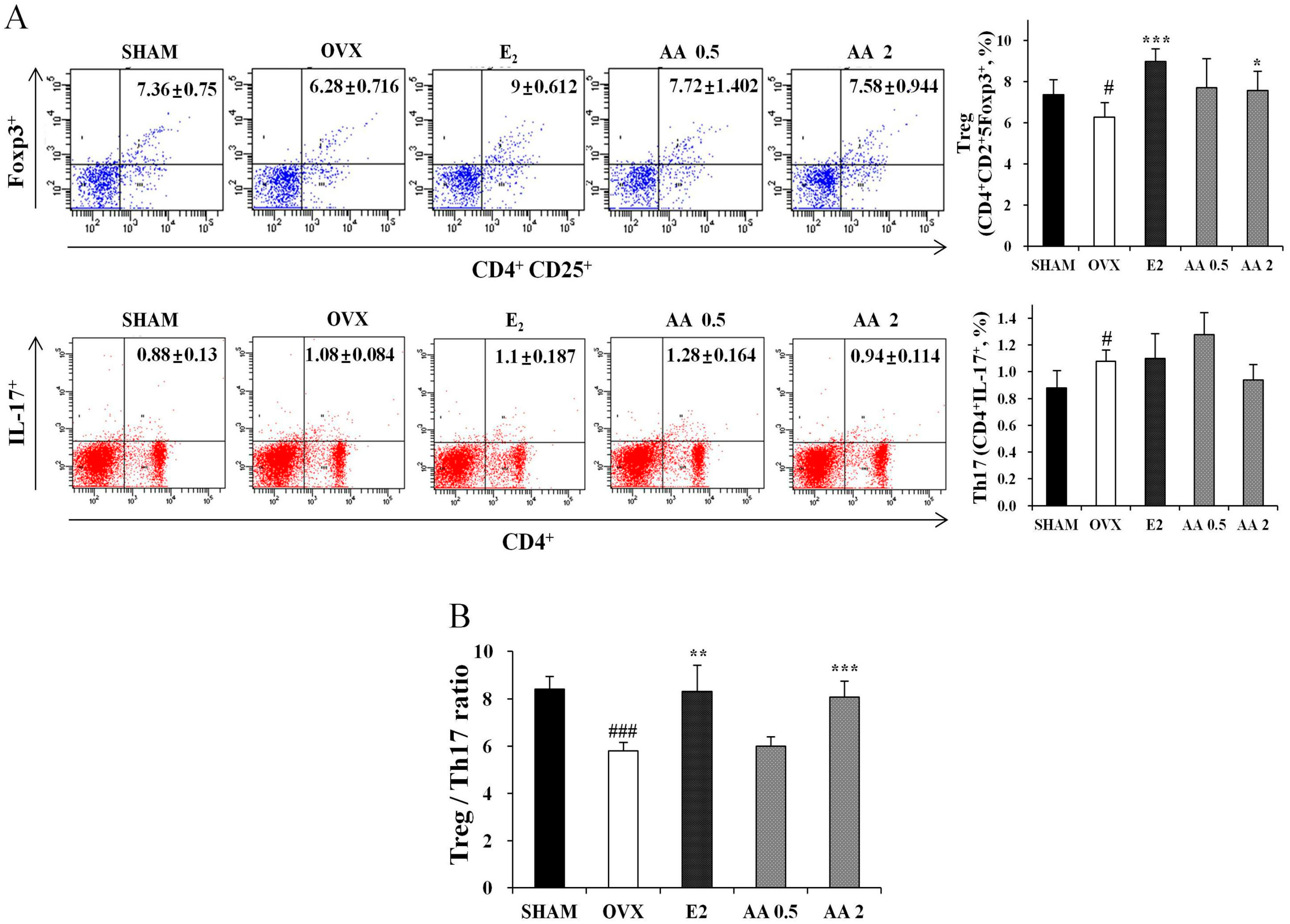
3.2. Effects of AA on the Balance between Th17 and Regulatory T Cell

3.3. Effects of AA on Serum Biochemical Markers (Ca and IP) and TCHO
| SHAM | OVX | E2 | AA 0.5 | AA 2 | |
|---|---|---|---|---|---|
| Calcium (mg/dl) | 10.32 ± 0.303 | 10.8 ± 0.173 # | 10.38 ± 0.277 * | 10.4 ± 0.212 * | 10.22 ± 0.205 ** |
| Phosphorus (mg/dl) | 5.22 ± 0.507 | 7.7 ± 1.632 # | 7.22 ± 0.963 | 6.86 ± 1.033 | 7.32 ± 1.094 |
| Total cholesterol (mg/dl) | 118.8 ± 9.834 | 197.4 ± 6.656 ### | 156.6 ± 5.459 *** | 177.8 ± 11.432 * | 162.2 ± 14.533 ** |
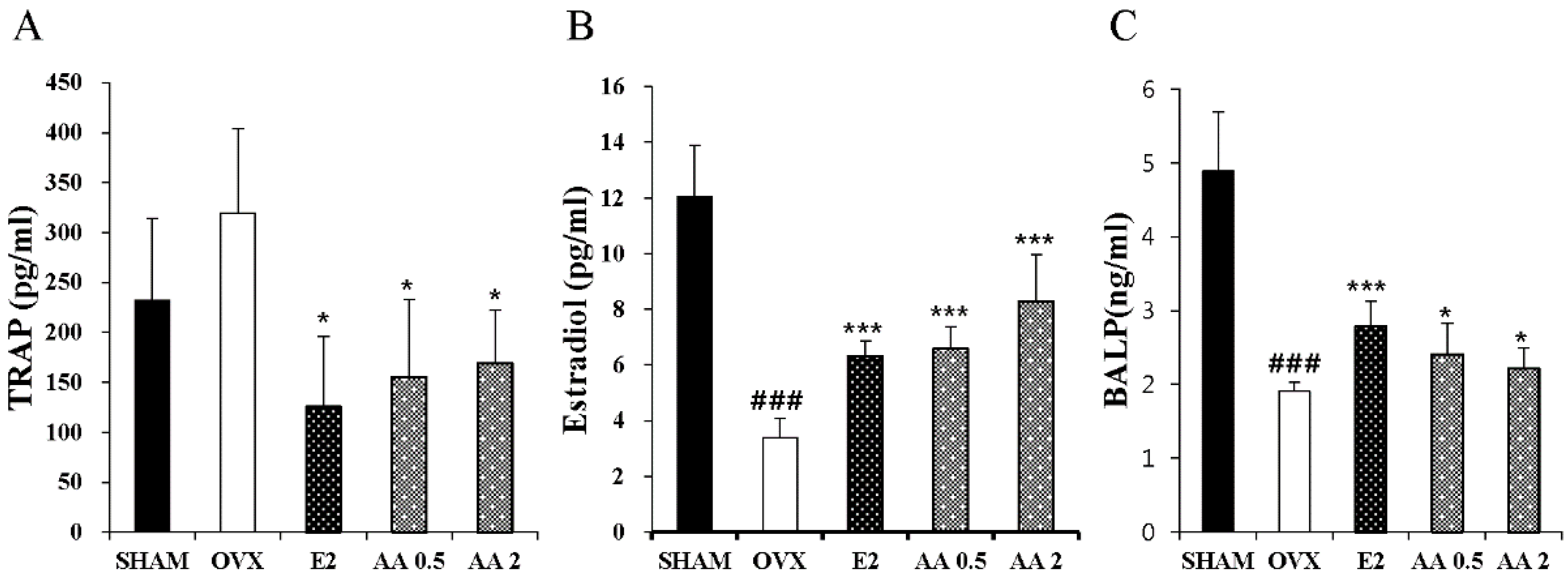
3.4. Effects of AA on Serum TRAP, E2, and BALP
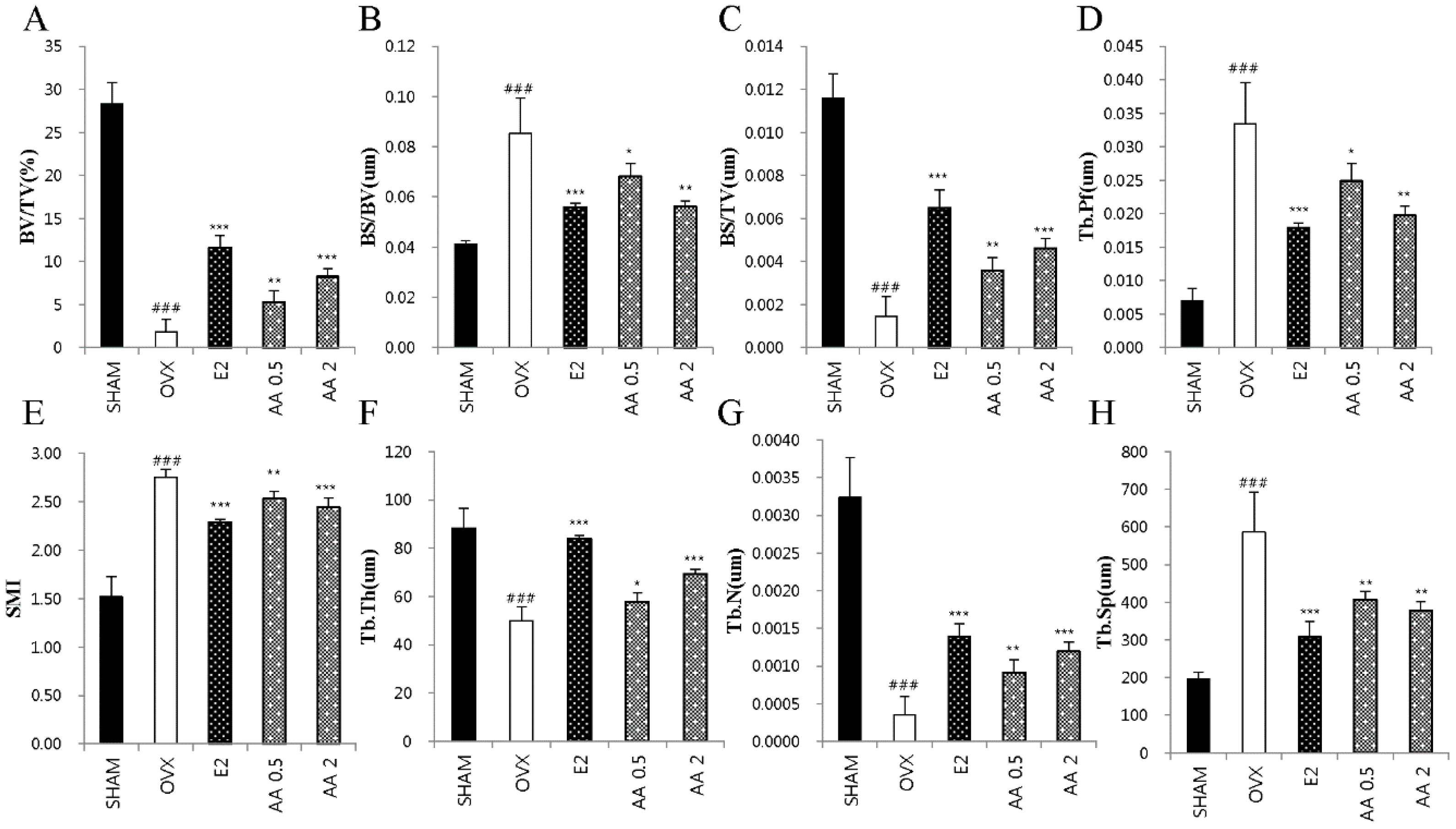
3.5. Effects of AA on Bone Microarchitecture


3.6. Discussion
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [PubMed]
- Tseng, S.H.; Sung, C.H.; Chen, L.G.; Lai, Y.J.; Chang, W.S.; Sung, H.C.; Wang, C.C. Comparison of chemical compositions and osteoprotective effects of different sections of velvet antler. J. Ethnopharmacol. 2014, 151, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Tantikanlayaporn, D.; Wichit, P.; Weerachayaphorn, J.; Chairoungdua, A.; Chuncharunee, A.; Suksamrarn, A.; Piyachaturawat, P. Bone sparing effect of a novel phytoestrogen diarylheptanoid from Curcuma comosa Roxb. in ovariectomized rats. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Raisz, L.G. Pathogenesis of osteoporosis: Concepts, conflicts, and prospects. J. Clin. Investig. 2005, 115, 3318–3325. [Google Scholar] [CrossRef] [PubMed]
- Lerner, U.H. Bone remodeling in post-menopausal osteoporosis. J. Dent. Res. 2006, 85, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.M.; Srivastava, K.; Mansoori, M.N.; Trivedi, R.; Chattopadhyay, N.; Singh, D. Estrogen deficiency induces the differentiation of IL-17 secreting Th17 cells: A new candidate in the pathogenesis of osteoporosis. PLoS ONE 2012. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Paul, W.E. Heterogeneity and plasticity of T helper cells. Cell Res. 2010, 20, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.O.; Zhang, H.; Kim, B.S.; Niu, X.; Peng, J.; Chen, Y.; Kerketta, R.; Lee, Y.H.; Chang, S.H.; Corry, D.B.; et al. The signaling suppressor CIS controls proallergic T cell development and allergic airway inflammation. Nat. Immunol. 2013, 14, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Bedoya, S.K.; Lam, B.; Lau, K.; Larkin, J. Th17 cells in immunity and autoimmunity. Clin. Dev. Immunol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Afzali, B.; Lombardi, G.; Lechler, R.I.; Lord, G.M. The role of T helper 17 (Th17) and regulatory T cells (Treg) in human organ transplantation and autoimmune disease. Clin. Exp. Immunol. 2007, 148, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Alves, C.H.; Farrell, E.; Vis, M.; Colin, E.M.; Lubberts, E. Animal Models of Bone Loss in Inflammatory Arthritis: From Cytokines in the Bench to Novel Treatments for Bone Loss in the Bedside-a Comprehensive Review. Clin. Rev. Allergy Immunol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, A.M.; Mansoori, M.N.; Srivastava, K.; Khan, M.P.; Kureel, J.; Dixit, M.; Shukla, P.; Trivedi, R.; Chattopadhyay, N.; Singh, D. Enhanced immunoprotective effects by anti-IL-17 antibody translates to improved skeletal parameters under estrogen deficiency compared with anti-RANKL and anti-TNF-α antibodies. J. Bone Miner. Res. 2014, 29, 1981–1992. [Google Scholar] [CrossRef] [PubMed]
- Downey, P.A.; Siegel, M.I. Bone biology and the clinical implications for osteoporosis. Phys. Ther. 2006, 86, 77–91. [Google Scholar] [PubMed]
- Lee, J.W.; Jhee, O.; Yuan, H.; Kim, T.; Kim, D.; Lee, M.; Om, A.; Lee, B.; Park, S.K.; Kang, J. Effect of Korean oriental medicine extract on bone mass as compared with alendronate in ovariectomized rats. J. Med. Food. 2005, 8, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.E.; Kim, G.M.; Lee, S.K.; Yang, C.J. Growth performance, meat yield, oxidative stability, and Fatty Acid composition of meat from broilers fed diets supplemented with a medicinal plant and probiotics. Asian-Australas. J. Anim. Sci. 2012, 25, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.E.; Ko, S.Y.; Kim, G.M.; Firman, J.D.; Yang, C.J. Evaluation of probiotic strains for development of fermented Alisma canaliculatum and their effects on broiler chickens. Poult. Sci. 2012, 91, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Mikamo, H.; Kawazoe, K.; Izumi, K.; Sato, Y.; Tamaya, T. Effects of crude herbal ingredients on intrauterine infection in a rat model. Curr. Ther. Res. 1998, 59, 122–127. [Google Scholar] [CrossRef]
- Huang, Y.T.; Huang, D.M.; Chueh, S.C.; Teng, C.M.; Guh, J.H. Alisol B acetate, a triterpene from Alismatis rhizoma, induces Bax nuclear translocation and apoptosis in human hormone-resistant prostate cancer PC-3 cells. Cancer Lett. 2006, 231, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.K.; Han, Y.R.; Nam, J.S.; Han, C.W.; Kim, B.J.; Jeong, H.S.; Ha, K.T.; Jung, M.H. Protective Effects of Alisma orientale Extract against Hepatic Steatosis via Inhibition of Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2015, 16, 26151–26165. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.J.; Leutou, A.S.; Yeon, J.T.; Choi, S.W.; Kim, S.H.; Yee, S.T.; Choi, K.H.; Nam, S.J.; Son, Y.J. The Inhibitory Effect of Alisol A 24-Acetate from Alisma canaliculatum on Osteoclastogenesis. Int. J. Endocrinol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Ray, N.F.; Chan, J.K.; Thamer, M.; Melton, L.J. Medical expenditures for the treatment of osteoporotic fractures in the United States in 1995: Report from the National Osteoporosis Foundation. J. Bone Miner. Res. 1997, 12, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Canonico, M.; Plu-Bureau, G.; Lowe, G.D.; Scarabin, P.Y. Hormone replacement therapy and risk of venous thromboembolism in postmenopausal women: Systematic review and meta-analysis. BMJ 2008, 336, 1227–1231. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.G.; Zhang, R.; Li, C.; Ma, X.; Liu, L.; Wang, J.P.; Mei, Q.B. The osteoprotective effect of Radix Dipsaci extract in ovariectomized rats. J. Ethnopharmacol. 2009, 123, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Wronski, T.J.; Dann, L.M.; Qi, H.; Yen, C.F. Skeletal effects of withdrawal of estrogen and diphosphonate treatment in ovariectomized rats. Calcif. Tissue Int. 1993, 53, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Notomi, T.; Okimoto, N.; Okazaki, Y.; Nakamura, T.; Suzuki, M. Tower climbing exercise started 3 months after ovariectomy recovers bone strength of the femur and lumbar vertebrae in aged osteopenic rats. J. Bone Miner. Res. 2003, 18, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Hidaka, S.; Okamoto, Y.; Nakajima, K.; Suekawa, M.; Liu, S.Y. Preventive effects of traditional Chinese (Kampo) medicines on experimental osteoporosis induced by ovariectomy in rats. Calcif. Tissue Int. 1997, 61, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Park, J.A.; Ha, S.K.; Kang, T.H.; Oh, M.S.; Cho, M.H.; Lee, S.Y.; Park, J.H.; Kim, S.Y. Protective effect of apigenin on ovariectomy-induced bone loss in rats. Life Sci. 2008, 82, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Sims, N.A.; Morris, H.A.; Moore, R.J.; Durbridge, T.C. Estradiol treatment transiently increases trabecular bone volume in ovariectomized rats. Bone 1996, 19, 455–461. [Google Scholar] [CrossRef]
- Swaminathan, R. Biochemical markers of bone turnover. Clin. Chim. Acta. 2001, 313, 95–105. [Google Scholar] [CrossRef]
- Fahmy, S.R.; Soliman, A.M.; Sayed, A.A.; Marzouk, M. Possible antiosteoporotic mechanism of Cicer arietinum extract in ovariectomized rats. Int. J. Clin. Exp. Pathol. 2015, 8, 3477–3490. [Google Scholar] [PubMed]
- Su, S.J.; Yeh, Y.T.; Shyu, H.W. The preventive effect of biochanin A on bone loss in ovariectomized rats: Involvement in regulation of growth and activity of osteoblasts and osteoclasts. Evid. Based Complement. Altern. Med. 2013. [Google Scholar] [CrossRef]
- Mohamed, M.K.; Abdel-Rahman, A.A. Effect of long-term ovariectomy and estrogen replacement on the expression of estrogen receptor gene in female rats. Eur. J. Endocrinol. 2000, 142, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.W.; Lee, Y.; Kim, Y.T. Preventive effects of Citrus unshiu peel extracts on bone and lipid metabolism in OVX rats. Molecules 2014, 19, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.W.; Ross, P.D.; Johnson, N.E.; Wasnich, R.D. Estrogen and calcium supplement use among Japanese-American women: Effects upon bone loss when used singly and in combination. Bone 1995, 17, 369–373. [Google Scholar] [CrossRef]
- Lai, N.; Zhang, Z.; Wang, B.; Miao, X.; Guo, Y.; Yao, C.; Wang, Z.; Wang, L.; Ma, R.; Li, X.; et al. Regulatory effect of traditional Chinese medicinal formula Zuo-Gui-Wan on the Th17/Treg paradigm in mice with bone loss induced by estrogen deficiency. J. Ethnopharmacol. 2015, 166, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.; Zhou, C.H.; Zhang, X.; Chen, Y.; Xu, B.L.; Cui, L.; Xu, D.H. Effect of 1,25-dihydroxyvitamin D3 on regulatory T cells in ovariectomized mice. Biomed. Environ. Sci. 2014, 27, 779–785. [Google Scholar] [PubMed]
- Recker, R.; Masarachia, P.; Santora, A.; Howard, T.; Chavassieux, P.; Arlot, M.; Rodan, G.; Wehren, L.; Kimmel, D. Trabecular bone microarchitecture after alendronate treatment of osteoporotic women. Curr. Med. Res. Opin. 2005, 21, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Wu, J.; Wang, S.; Wei, Y.; Chen, F.; Chen, J.; Shi, J.; Xia, J. Pycnogenol® treatment inhibits bone mineral density loss and trabecular deterioration in ovariectomized rats. Int. J. Clin. Exp. Med. 2015, 8, 10893–10901. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compound Alisol A 24 acetate is available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, Y.-H.; Kang, K.-Y.; Lee, S.-J.; Nam, S.-J.; Son, Y.-J.; Yee, S.-T. The Protective Effects of Alisol A 24-Acetate from Alisma canaliculatum on Ovariectomy Induced Bone Loss in Vivo. Molecules 2016, 21, 74. https://doi.org/10.3390/molecules21010074
Hwang Y-H, Kang K-Y, Lee S-J, Nam S-J, Son Y-J, Yee S-T. The Protective Effects of Alisol A 24-Acetate from Alisma canaliculatum on Ovariectomy Induced Bone Loss in Vivo. Molecules. 2016; 21(1):74. https://doi.org/10.3390/molecules21010074
Chicago/Turabian StyleHwang, Yun-Ho, Kyung-Yun Kang, Sung-Ju Lee, Sang-Jip Nam, Young-Jin Son, and Sung-Tae Yee. 2016. "The Protective Effects of Alisol A 24-Acetate from Alisma canaliculatum on Ovariectomy Induced Bone Loss in Vivo" Molecules 21, no. 1: 74. https://doi.org/10.3390/molecules21010074
APA StyleHwang, Y.-H., Kang, K.-Y., Lee, S.-J., Nam, S.-J., Son, Y.-J., & Yee, S.-T. (2016). The Protective Effects of Alisol A 24-Acetate from Alisma canaliculatum on Ovariectomy Induced Bone Loss in Vivo. Molecules, 21(1), 74. https://doi.org/10.3390/molecules21010074