
The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins


Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source Protein | Assays | Sequence | Ref. * |
|---|---|---|---|
| Chicken egg white | ORAC | AEERYP, DEDTQAMP | [9] |
| Egg white | oxygen radical scavenging, DPPH radical scavenging. | DHTKE, MPDAHL, FFGFN | [10] |
| Rice residue protein | DPPH and ABTS radical scavenging, FRAP-Fe3+ reducing assay. | RPNYTDA, TSQLLSDQ, TRTGDPFF, NFHPQ | [11] |
| Grass carp (Ctenopharyngodon idella) skin | DPPH radical, hydroxyl radical, ABTS radical scavenging, Inhibiting lipid peroxidation. | PYSFK, GFGPEL, GGRP | [12] |
| Bluefin leatherjacket skin (Navodon septentrionalis) | DPPH, HO, O2− radical scavenging. | GSGGL, GPGGFI, FIGP | [6] |
| Palm kernel cake proteins | DPPH radical scavenging, Metal chelating ability. | GIFE LPWRPATNVF | [13] |
| Blood clam (Tegillarca granosa) muscle | lipid peroxidation, radical scavenging activity. | WPP | [14] |
| Sweet potato | OH radical scavenging. | YYIVS | [15] |
| Croceine croaker (Pseudosciaena crocea) muscle | DPPH, superoxide, ABTS and hydroxyl radical scavenging, lipid peroxidation. | WLMSA, VLWEE, MILMR | [16] |
| Spotless smoothhound (Mustelus griseus) muscle | Hydroxyl, ABTS, superoxide radical scavenging. | GIISHR, ELLI, KFPE, GFVG, GAA | [17] |
| Bluefin leatherjacket (Navodon septentrionalis) heads | DPPH, hydroxyl, ABTS, superoxide radicals scavenging. | WEGPK, GPP, GVPLT | [18] |
| Hemp (Cannabis sativa L.) seed | DPPH radicals scavenging. | PSLPA, WVYY | [19] |
| Chickpea protein | DPPH radicals scavenging. | VGDI, DHG | [20] |
| Marine Sepia brevimana mantle | DPPH radicals scavenging, lipid peroxidation. | I/LNI/LCCN | [21] |
| Sphyrna lewini muscle | ABTS, DPPH radicals scavenging. | WDR, PYFNK | [22] |
| Tilapia (Oreochromis niloticus) gelatin | Hydroxyl radicals scavenging. | LSGYGP | [23] |
| Corn gluten meal | DPPH, ABTS, and hydroxyl radicals scavenging. | LLPF | [24] |
| Oyster (Saccostrea cucullata) | DPPH radicals scavenging, Inhibiting human colon carcinoma (HT-29) cell lines. | LANAK, PSLVGRPPVGKLTL, VKVLLEHPVL | [25] |
| Corp | Preventing hepatic fibrosis. | LLPF, FLPE | [26] |
2. The Mechanism of the Antioxidation and the Evaluating Methods of Antioxidant Capacity
3. The Relationship of Chemical Structure and the Antioxidant Ability of Peptides
3.1. Effect of the Structure of Precursor Proteins and the Hydrolytic Process on the Antioxidant Activities
3.2. The Relationship of Peptide Structure and Its Antioxidant Activity
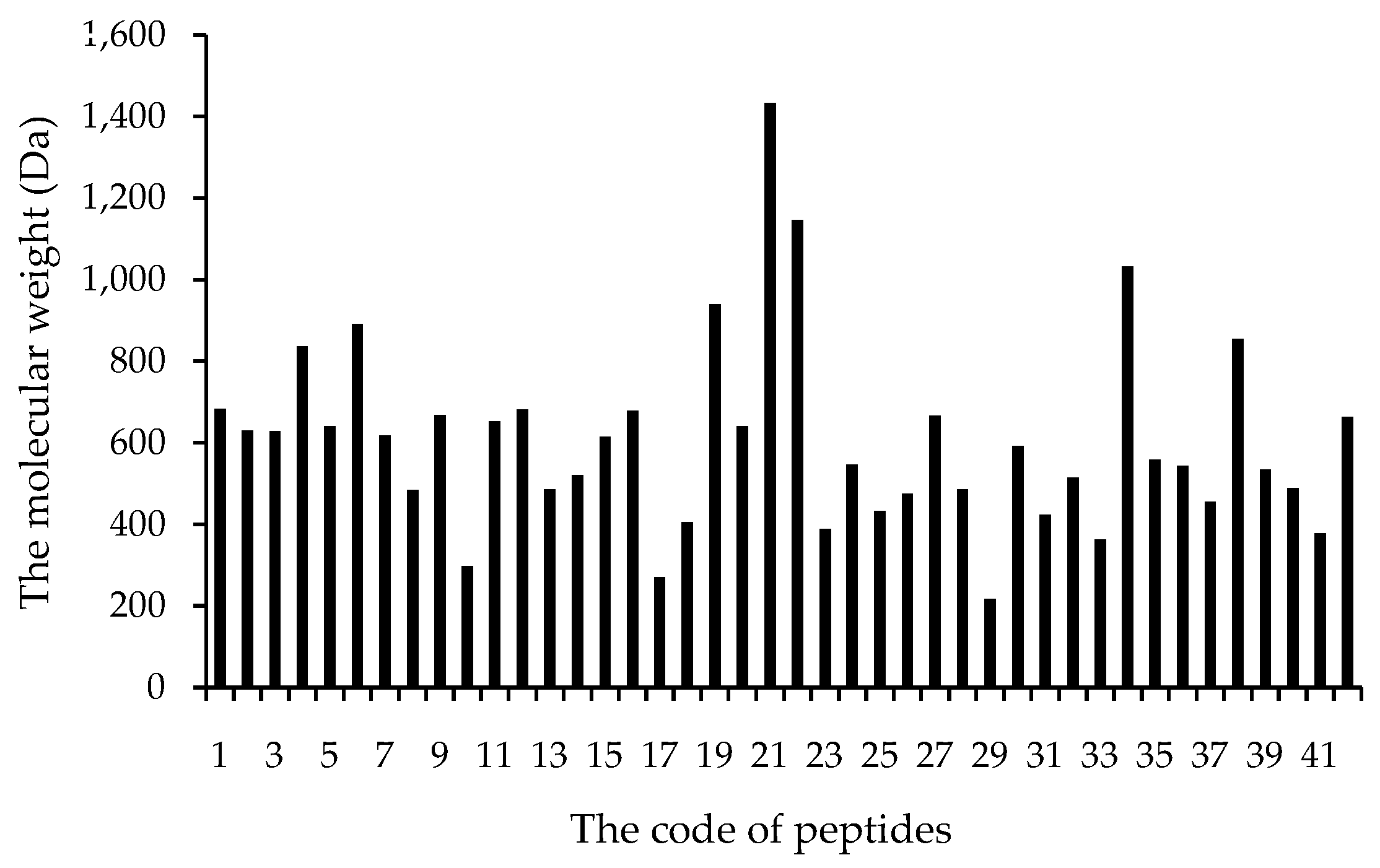
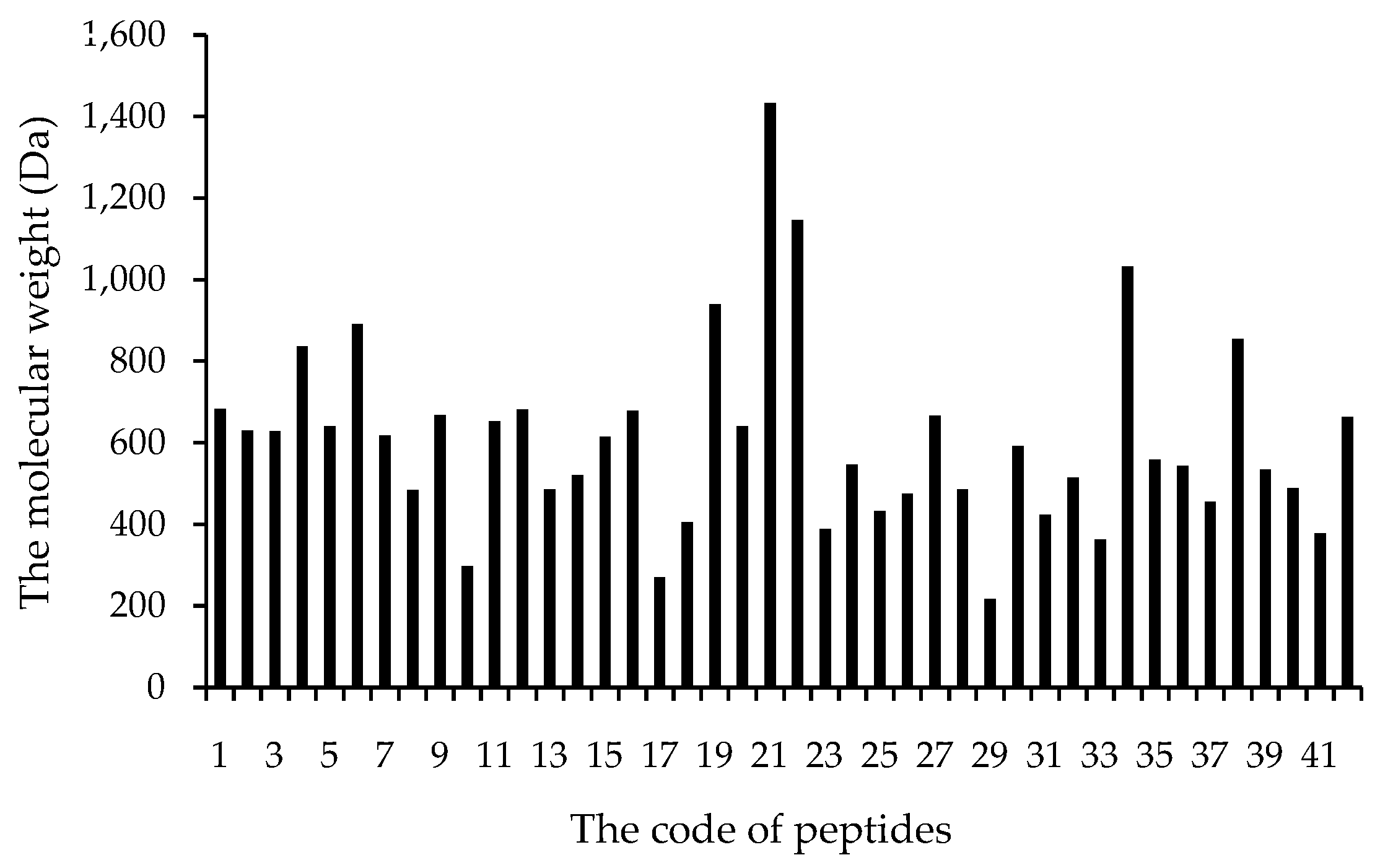
3.2.1. Molecular Weight
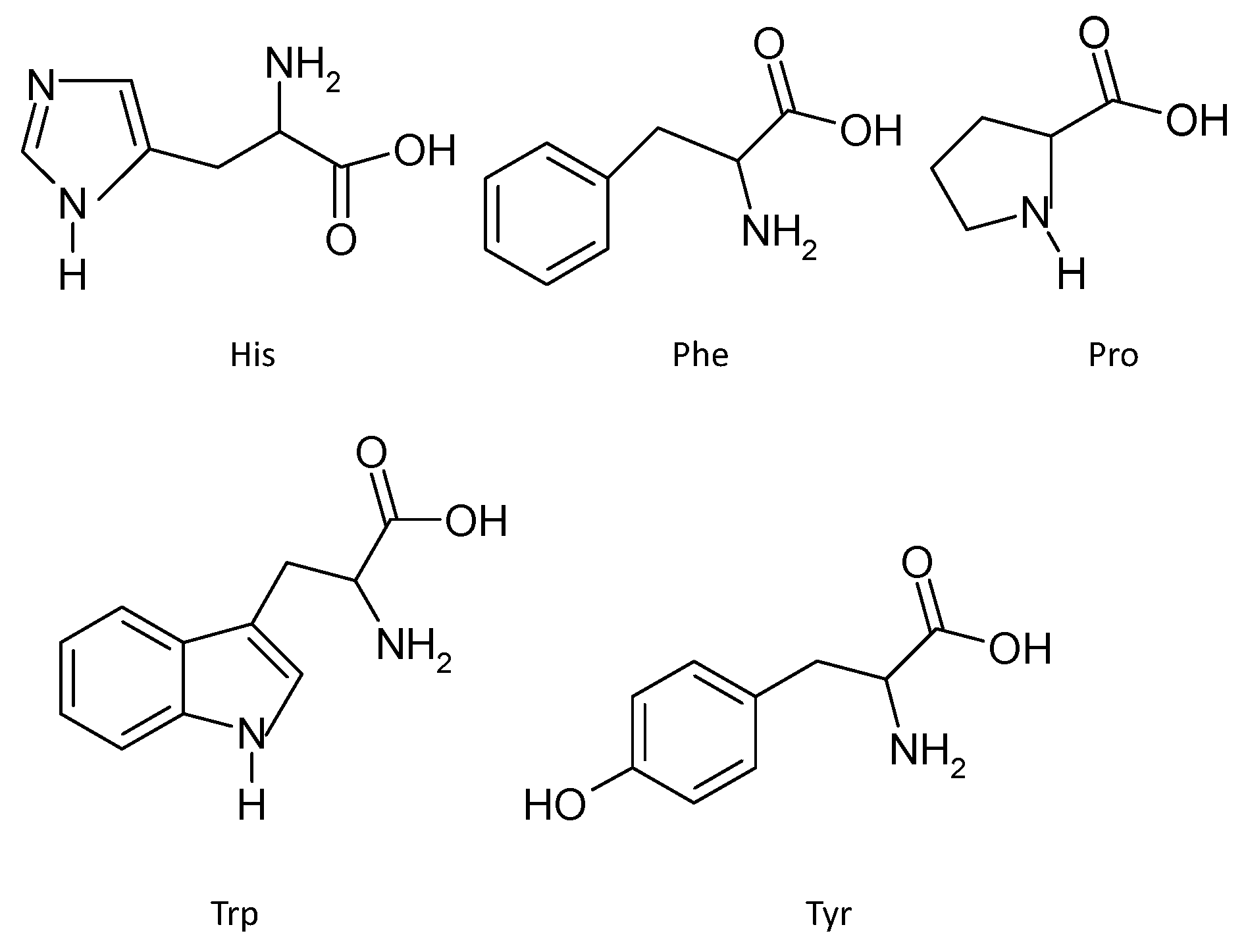
3.2.2. Amino Acid Composition
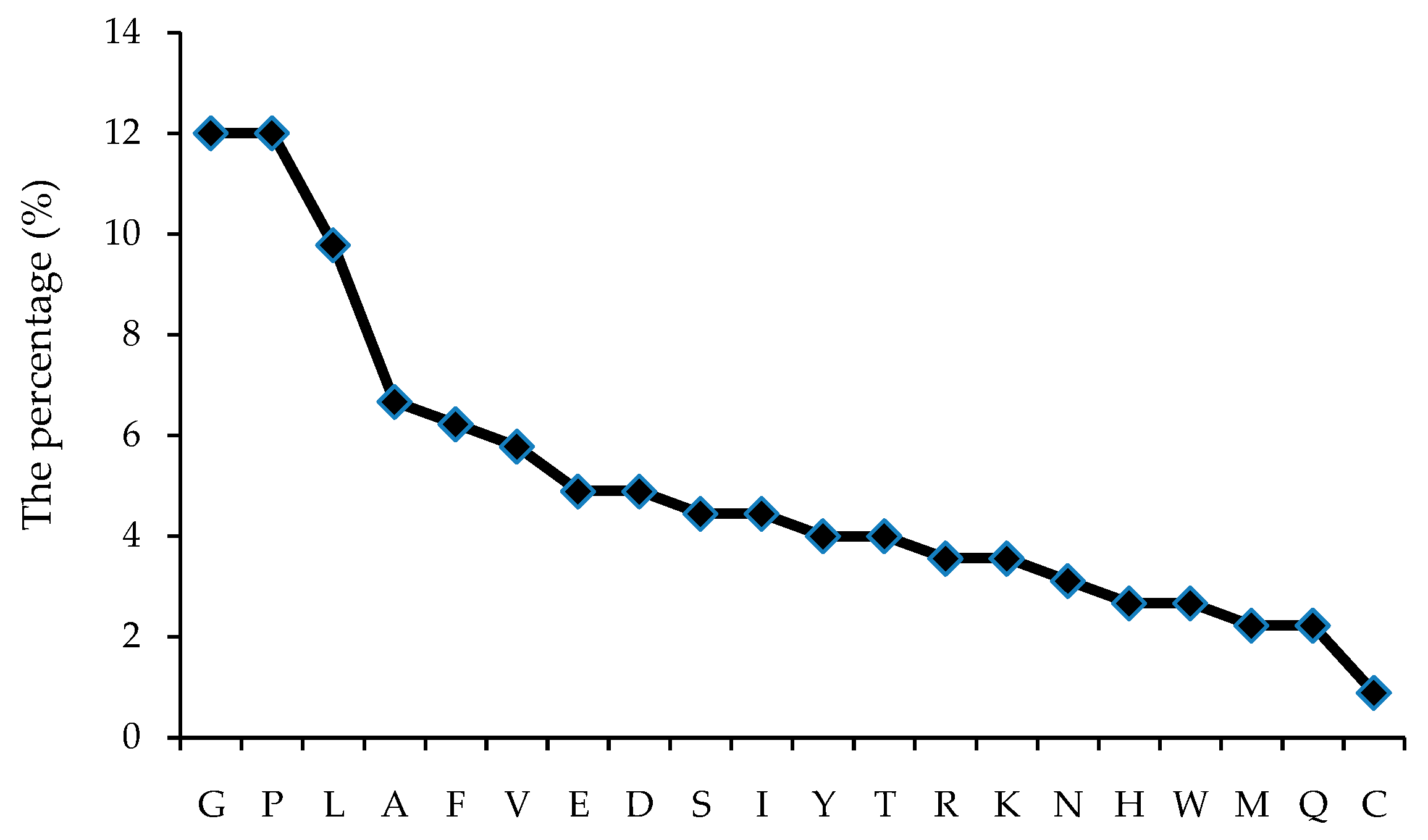
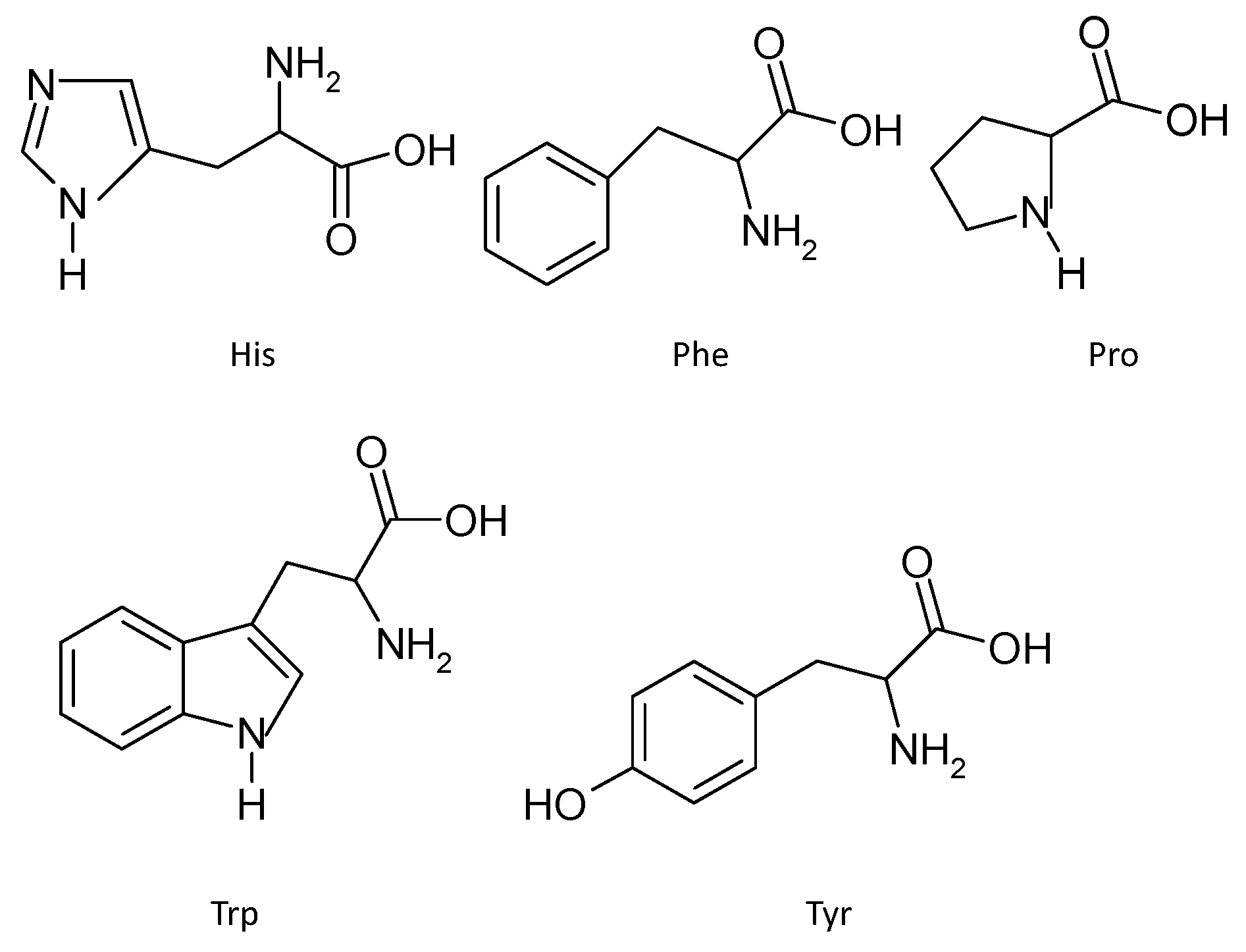
3.2.3. Amino Acids Sequence
3.2.4. Secondary Structure
3.2.5. Antioxidant Peptides’ Stability and Their Synergistic Effects
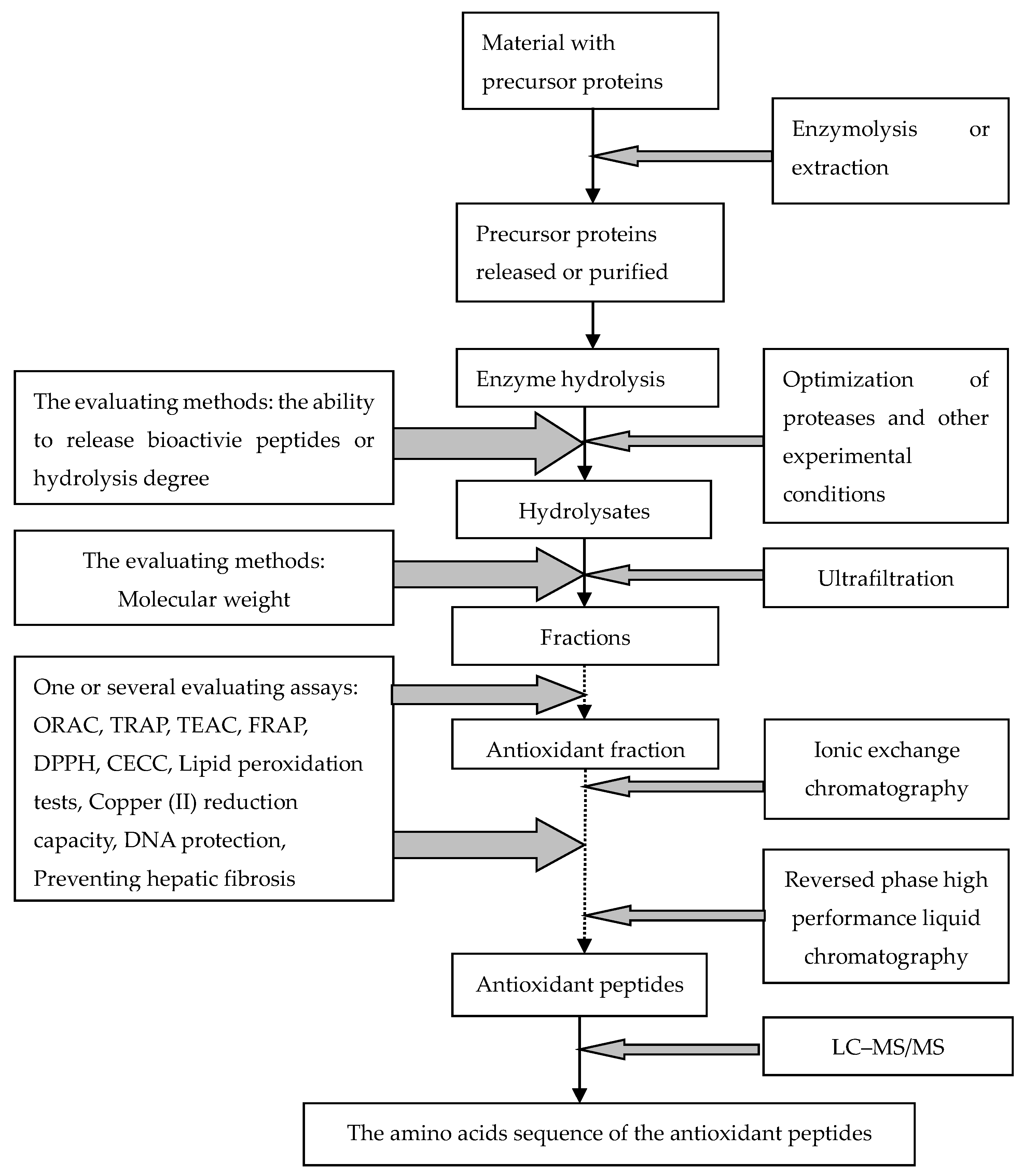
4. Systematic Schemes to Predict the Antioxidative Activity of Peptides
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Montoya-Rodriguez, A.; de Mejia, E.G. Pure peptides from amaranth (Amaranthus hypochondriacus) proteins inhibit LOX-1 receptor and cellular markers associated with atherosclerosis development in vitro. Food Res. Int. 2015. [Google Scholar] [CrossRef]
- Moller, N.P.; Scholz-Ahrens, K.E.; Roos, N.; Schrezenmeir, J. Bioactive peptides and proteins from foods: Indication for health effects. Eur. J. Nutr. 2008, 47, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Samaranayaka, A.G.P.; Li-Chan, E.C.Y. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J. Funct. Foods 2011, 3, 229–254. [Google Scholar] [CrossRef]
- Vásquez-Villanueva, R.; Marina, M.L.M.; Garcia, C. Identification by hydrophilic interaction and reversed-phase liquid chromatography-tandem mass spectrometry of peptides with antioxidant capacity in food residues. J. Chromatogr. A 2015. [Google Scholar] [CrossRef]
- Dziuba, J.; Niklewicz, M.; Iwaniak, I.; Darewicz, M.; Minkiewicz, P. Bioinformatic-aided prediction for release possibilities of bioactive peptides from plant proteins. Acta. Aliment. 2004, 33, 227–235. [Google Scholar] [CrossRef]
- Chi, C.F.; Wang, B.; Hu, F.Y.; Wang, Y.M.; Zhang, B.; Deng, S.J.; Wu, C.W. Purification and identification of three novel antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) skin. Food Res. Int. 2015, 73, 124–129. [Google Scholar] [CrossRef]
- Song, R.; Wei, R.B.; Ruan, G.Q.; Luo, H.Y. Isolation and identification of antioxidant peptides from peptic hydrolysates of half-fin anchovy (Setipinna taty). LWT-Food Sci. Technol. 2015, 60, 221–229. [Google Scholar] [CrossRef]
- Duan, X.; Ocen, D.; Wu, F.F.; Li, M.; Yang, N.; Xu, J.; Chen, H.Y.; Huang, L.Q.; Jin, Z.Y.; Xu, X.M. Purification and characterization of a natural antioxidant peptide from fertilized eggs. Food Res. Int. 2014, 56, 18–24. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Bandara, N.; Wu, J.P. Purification and characterization of antioxidant peptides from enzymatically hydrolyzed chicken egg white. Food Chem. 2015, 188, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.B.; Jin, Y.; Lin, S.Y.; Jones, G.S.; Chen, F. Purification and identification of novel antioxidant peptides from egg white protein and their antioxidant activities. Food Chem. 2015, 175, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.J.; Huang, L.H.; Sun, Q.; Jiang, Z.Q.; Wu, X. Isolation, identification and synthesis of four novel antioxidant peptides from rice residue protein hydrolyzed by multiple proteases. Food Chem. 2015, 179, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.Y.; Wu, X.S.; Zhang, Y.H.; Li, X.X.; Ma, S.; Li, J.R. Purification and characterization of three antioxidant peptides from protein hydrolysate of grass carp (Ctenopharyngodon idella) skin. J. Funct. Foods 2015, 16, 234–242. [Google Scholar] [CrossRef]
- Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Anwar, F.; Bakar, F.A.; Philip, R.; Saari, N. Identification and characterization of papain-generated antioxidant peptides from palm kernel cake proteins. Food Res. Int. 2014, 62, 726–734. [Google Scholar] [CrossRef]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Li, T.; Ding, G.F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Zhang, M.; Mu, T.H.; Sun, M.J. Purification and identification of antioxidant peptides from sweet potato protein hydrolysates by Alcalase. J. Funct. Foods 2014, 7, 191–200. [Google Scholar] [CrossRef]
- Chi, C.F.; Hu, F.Y.; Wang, B.; Ren, X.J.; Deng, S.J.; Wu, C.W. Purification and characterization of three antioxidant peptides from protein hydrolyzate of croceine croaker (Pseudosciaena crocea) muscle. Food Chem. 2015, 168, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gong, Y.D.; Li, Z.R.; Yu, D.; Chi, C.F.; Ma, J.Y. Isolation and characterisation of five novel antioxidant peptides from ethanol-soluble proteins hydrolysate of spotless smoothhound (Mustelus griseus) muscle. J. Funct. Foods 2014, 6, 176–185. [Google Scholar] [CrossRef]
- Chi, C.F.; Wang, B.; Wang, Y.M.; Zhang, B.; Deng, S.J. Isolation and characterization of three antioxidant peptides from protein hydrolysate of bluefin leatherjacket (Navodon septentrionalis) heads. J. Funct. Foods 2015, 12, 1–10. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Malomo, S.; Offengenden, M.; Wu, J.P.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Ghribi, A.M.; Sila, A.; Przybylski, R.; Nedjar-Arroume, N.; Makhlouf, I.; Blecker, C.; Attia, H.; Dhulster, P.; Bougatef, A.; Besbes, S. Purification and identification of novel antioxidant peptides from enzymatic hydrolysate of chickpea (Cicer arietinum L.) protein concentrate. J. Funct. Foods 2015, 12, 516–525. [Google Scholar] [CrossRef]
- Sudhakar, S.; Nazeer, R.A. Preparation of potent antioxidant peptide from edible part of shortclub cuttlefish against radical mediated lipid and DNA damage. LWT-Food Sci. Technol. 2015, 64, 593–601. [Google Scholar] [CrossRef]
- Wang, B.; Li, Z.R.; Chi, C.F.; Zhang, Q.H.; Luo, H.Y. Preparation and evaluation of antioxidant peptides from ethanol-soluble proteins hydrolysate of Sphyrna lewini muscle. Peptides 2012, 36, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Zhang, Y.F.; Zhuang, Y.L. Antiphotoaging effect and purification of an antioxidant peptide from tilapia (Oreochromis niloticus) gelatin peptides. J. Funct. Foods 2013, 5, 154–162. [Google Scholar] [CrossRef]
- Zhuang, H.; Tang, N.; Yuan, Y. Purification and identification of antioxidant peptides from corn gluten meal. J. Funct. Food 2013, 5, 1810–1821. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Sivagami, G.; Balasubramanian, T. Antioxidant activity and anticancer effect of bioactive peptide from enzymatic hydrolysate of oyster (Saccostrea cucullata). Biomed. Prev. Nutr. 2014, 4, 343–353. [Google Scholar] [CrossRef]
- Wang, C.; He, H.; Zhang, J.L.; Li, X.; Ma, Z.L. High performance liquid chromatography (HPLC) fingerprints and primary structure identification of corn peptides by HPLC-diode array detection and HPLC-electrospray ionization tandem mass spectrometry. J. Food Drug Anal. 2015. [Google Scholar] [CrossRef]
- Borges, R.S.; Castle, S.L. The antioxidant properties of salicylate derivatives: A possible new mechanism of anti-inflammatory activity. Bioorgan. Med. Chem. Lett. 2015, 25, 4808–4811. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assay. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, M.; Mirdamadi, S.; Ehsani, M.R.; Aminlari, M.; Hosseini, E. Purification and identification of antioxidant and ACE-inhibitory peptide from Saccharomyces cerevisiae protein hydrolysate. J. Funct. Foods 2015, 19, 259–268. [Google Scholar] [CrossRef]
- Pouzo, L.B.; Descalzo, A.M.; Zaritzky, N.E.; Rossetti, L.; Pavan, E. Antioxidant status, lipid and color stability of aged beef from grazing steers supplemented with corn grain and increasing levels of flaxseed. Meat Sci. 2016, 111, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Kovacs-Nolan, J.; Jiang, B.; Tsao, R.; Mine, Y. Peptides derived from eggshell membrane improve antioxidant enzyme activity and glutathione synthesis against oxidative damage in Caco-2 cells. J. Funct. Foods 2014, 11, 571–580. [Google Scholar] [CrossRef]
- Zheng, L.; Zhao, M.; Xiao, C.; Zhao, Q.; Su, G. Practical problems when using ABTS assay to assess the radical-scavenging activity of peptides: Importance of controlling reaction pH and time. Food Chem. 2016, 192, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Fang, B.; Jiang, L.; Guo, H.; Cui, J.Y. Structure-activity relationship of a series of antioxidant tripeptides derived from β-Lactoglobulin using QSAR modeling. Dairy Sci. Technol. 2015, 176, 1815–1833. [Google Scholar] [CrossRef]
- Carbonaro, M.; Nardini, M.; Maselli, P.; Nucara, A. Chemico-physical and nutritional properties of traditional legumes (lentil, Lens culinaris L., and grass pea, Lathyrus sativus L.) from organic agriculture: An explorative study. Org. Agric. 2015, 16, 334–335. [Google Scholar] [CrossRef]
- Ahmed, A.S.; El-Bassiony, T.; Elmalt, L.M.; Ibrahim, H.R. Identification of potent antioxidant bioactive peptides from goat milk proteins. Food Res. Int. 2015, 74, 80–88. [Google Scholar] [CrossRef]
- Capriotti, A.L.; Caruso, G.; Cavaliere, C.; Samperi, R.; Ventura, S.; Chiozzi, R.Z.; Laganà, A. Identification of potential bioactive peptides generated by simulated gastrointestinal digestion of soybean seeds and soy milk proteins. J. Food Compos. Anal. 2015, 44, 205–213. [Google Scholar] [CrossRef]
- Tironi, V.A.; Anon, M.C. Amaranth proteins as a source of antioxidant peptides: Effect of proteolysis. Food Res. Int. 2010, 43, 315–322. [Google Scholar] [CrossRef]
- Ren, Y.; Wu, H.; Li, X.; Lai, F.; Xiao, X. Purification and characterization of high antioxidant peptides from duck egg white protein hydrolysates. Biochem. Biophys. Res. Commun. 2014, 452, 888–894. [Google Scholar] [CrossRef] [PubMed]
- Esteve, C.; Marina, M.L.; García, M.C. Novel strategy for the revalorization of olive (Olea europaea) residues based on the extraction of bioactive peptides. Food Chem. 2015, 167, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidant and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Li, B. The effect of molecular weights on the survivability of casein-derived antioxidant peptides after the simulated gastrointestinal digestion. Innov. Food Sci. Emerg. Technol. 2012, 16, 341–348. [Google Scholar] [CrossRef]
- Lin, S.Y.; Jin, Y.; Liu, M.Y.; Yang, Y.; Zhang, M.S.; Guo, Y.; Jones, G.; Liu, J.B.; Yin, Y.G. Research on the preparation of antioxidant peptides derived from egg white with assisting of high-intensity pulsed electric field. Food Chem. 2013, 139, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Saidi, S.; Deratani, A.; Belleville, M.P.; Amar, R.B. Antioxidant properties of peptide fractions from tuna dark muscle protein by-product hydrolysate produced by membrane fractionation process. Food Res. Int. 2014, 65, 329–336. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Contreras, M.M.; Recio, I.; Alaiz, M.; Vioque, J. Identification and characterization of antioxidant peptides from chickpea protein hydrolysates. Food Chem. 2015, 180, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, A.; Nedjar-Arroume, N.; Manni, L.; Ravallec, R.; Barkia, A.; Guillochon, D.; Nasri, M. Purification and identification of novel antioxidant peptides from enzymatic hydrolysates of sardinelle (Sardinella aurita) by-products proteins. Food Chem. 2010, 118, 559–565. [Google Scholar] [CrossRef]
- He, R.; Ju, X.; Yuan, J.; Wang, L.; Girgih, A.T.; Aluko, R.E. Antioxidant activities of rapeseed peptides produced by solid state fermentation. Food Res. Int. 2012, 49, 432–438. [Google Scholar] [CrossRef]
- Chen, H.; Zhao, M.; Lin, L.; Wang, J.; Sun-Waterhouse, D.; Dong, Y.; Zhuang, M.; Su, G. Identification of antioxidant peptides from defatted walnut meal hydrolysate with potential for improving learning and memory. Food Res. Int. 2015, 78, 216–233. [Google Scholar] [CrossRef]
- Eftekharzadeh, B.; Khodagholi, F.; Abdi, A.; Maghsoudi, N. Alginate protects NT2 neurons against H2O2-induced neurotoxicity. Carbohydr. Polym. 2010, 79, 1063–1072. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Isolation, purification and identification of three novel antioxidant peptides from patin (Pangasius sutchi) myofibrillar protein hydrolysates. LWT-Food Sci. Technol. 2015, 60, 452–461. [Google Scholar] [CrossRef]
- Bamdad, F.; Ahmed, S.; Chen, L. Specifically designed peptide structures effectively suppressed oxidative reactions in chemical and cellular systems. J. Funct. Foods 2015, 18, 35–46. [Google Scholar] [CrossRef]
- Canabady-Rochelle, L.L.S.; Harscoat-Schiavo, C.; Kessler, V.; Aymes, A.; Fournier, F.; Girardet, J.M. Determination of reducing power and metal chelating ability of antioxidant peptides: Revisited methods. Food Chem. 2015, 183, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Farvin, K.H.S.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 2—Characterisation of peptide fractions. Food Chem. 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Ren, J.; Zhao, M.; Shi, J.; Wang, J.; Jiang, Y.; Cui, C.; Kakuda, Y.; Xue, S.J. Purification and identification of antioxidant peptides from grass carp muscle hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. 2008, 108, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.W.; Li, B. Characterization of structure-antioxidant activity relationship of peptides in free radical systems using QSAR models: Key sequence positions and their amino acid properties. J. Theor. Biol. 2013, 318, 29–43. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Garg, A.; Raghava, G.P. PEP str: A de ovo method for tertiary structure prediction of small bioactive peptides. Protein Pept. Lett. 2007, 14, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Natarajan, P.; Forte, T.M.; Bielicki, J.K. Thiol-bearing synthetic peptides retain the antioxidant activity of apolipoproteinA-IMilano. Biochem. Biophys. Res. Commun. 2002, 297, 206–213. [Google Scholar] [CrossRef]
- Jiménez-Escrig, A.; Alaiz, M.; Vioque, J.; Rupérez, P. Health-promoting activities of ultrafiltered okara protein hydrolysates released by in vitro gastrointestinal digestion: Identification of active peptide from soybean lipoxygenase. Eur. Food Res Technol. 2010, 230, 655–663. [Google Scholar] [CrossRef]
- Hsieh, C.C.; Hernández-Ledesma, B.; Jeong, H.J.; Park, J.H.; de Lumen, B.O. Complementary roles in cancer prevention: Protease inhibitor makes the cancer preventive peptide lunasin bioavailable. PLoS ONE 2010, 5, e8890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Northfield, S.E.; Wang, C.K.; Schroeder, C.I.; Durek, T.; Kan, M.W.; Swdberg, J.E. Disulfide-rich macrocyclic peptides as templates in drug design. Eur. Food Res Technol. 2014, 77, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, A.; Yue, T.; Wang, J.; Wang, Y.; Su, J. Purification and identification of five novel antioxidant peptides from goat milk casein hydrolysates. J. Dairy Sci. 2013, 96, 4242–4251. [Google Scholar] [CrossRef] [PubMed]
- Wegner, J.K.; Sterling, A.; Guha, R.; Bender, A.; Faulon, J.L.; Hastings, J.; O’Boyle, N.; Overington, J.; van Vlijmen, H.; Willighagen, E. Open-source chemistry software and molecular databases broaden the research horizons in drug discovery. Commun. ACM 2012, 55, 65–75. [Google Scholar] [CrossRef]
- Cheng, Y.; Luo, F.; Zeng, Z.; Wen, L.; Xiao, Z. DFT-based quantitative structure-activity relationship studies for antioxidant peptides. Struct. Chem. 2015, 26, 739–747. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticalsand functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef]
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. https://doi.org/10.3390/molecules21010072
Zou T-B, He T-P, Li H-B, Tang H-W, Xia E-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules. 2016; 21(1):72. https://doi.org/10.3390/molecules21010072
Chicago/Turabian StyleZou, Tang-Bin, Tai-Ping He, Hua-Bin Li, Huan-Wen Tang, and En-Qin Xia. 2016. "The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins" Molecules 21, no. 1: 72. https://doi.org/10.3390/molecules21010072