
Flavones Isolated from Scutellariae radix Suppress Propionibacterium Acnes-Induced Cytokine Production In Vitro and In Vivo

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results
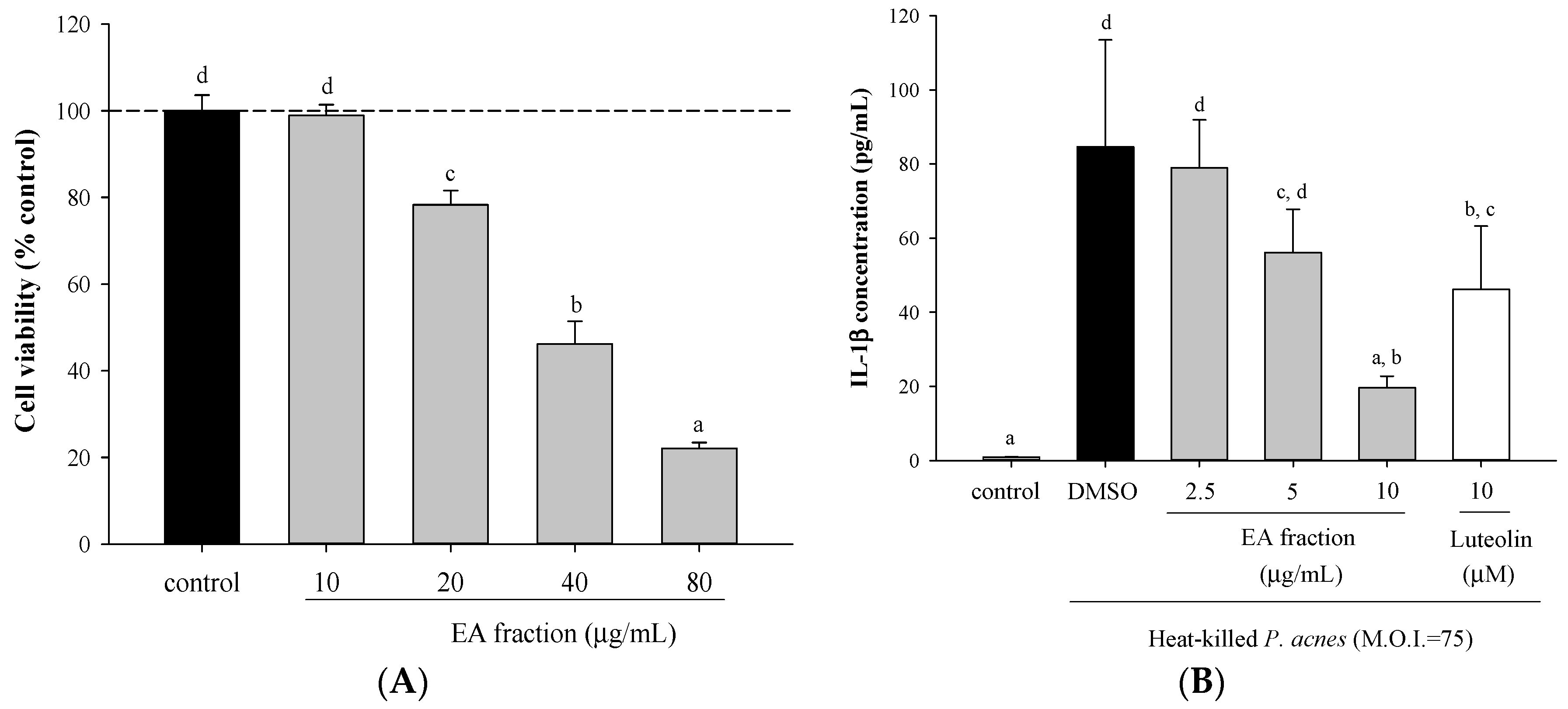
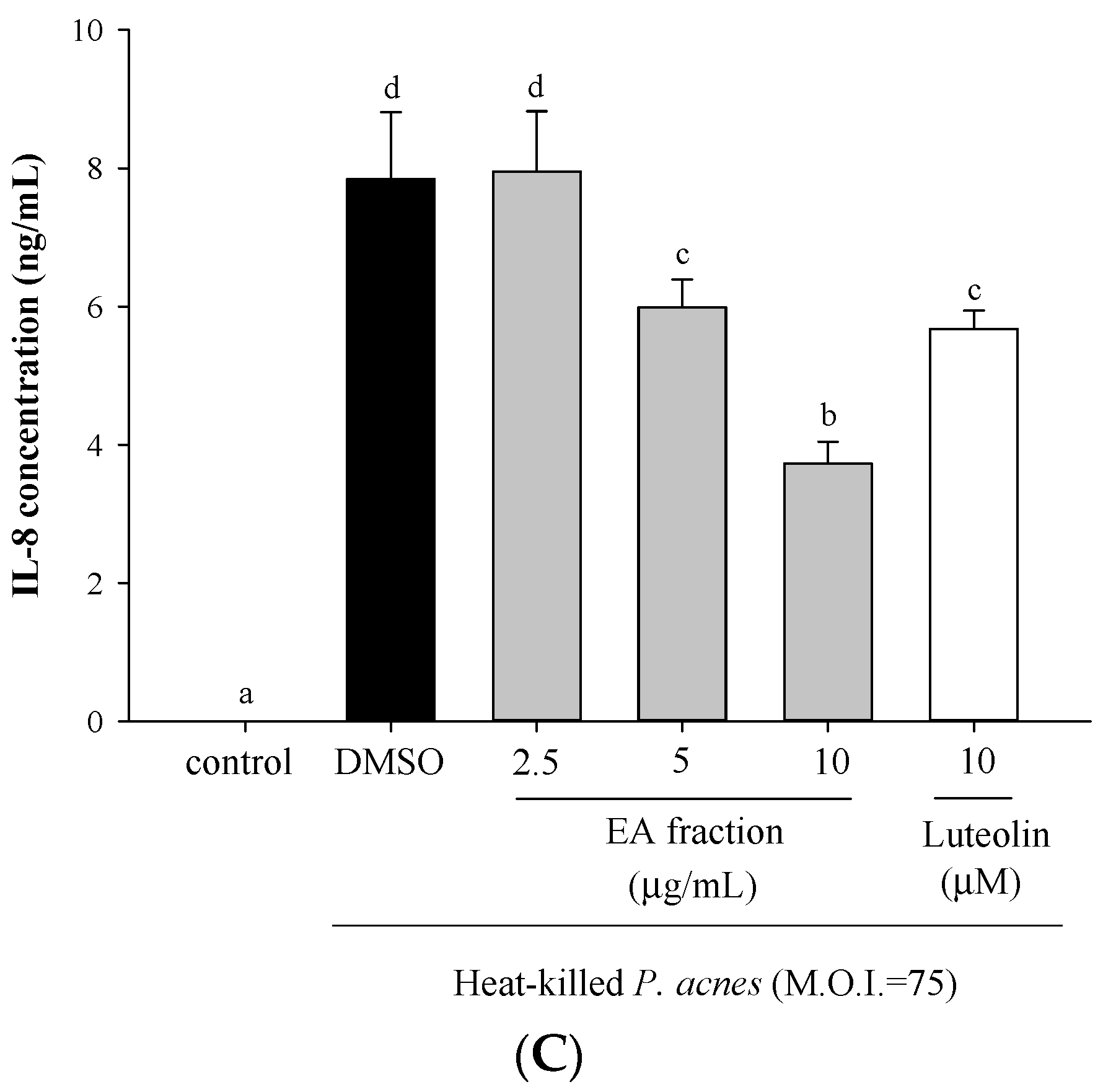
2.1. Effects of Scutellariae Radix Extracts on P. acnes-Induced IL-1β and IL-8 Production in THP-Cells


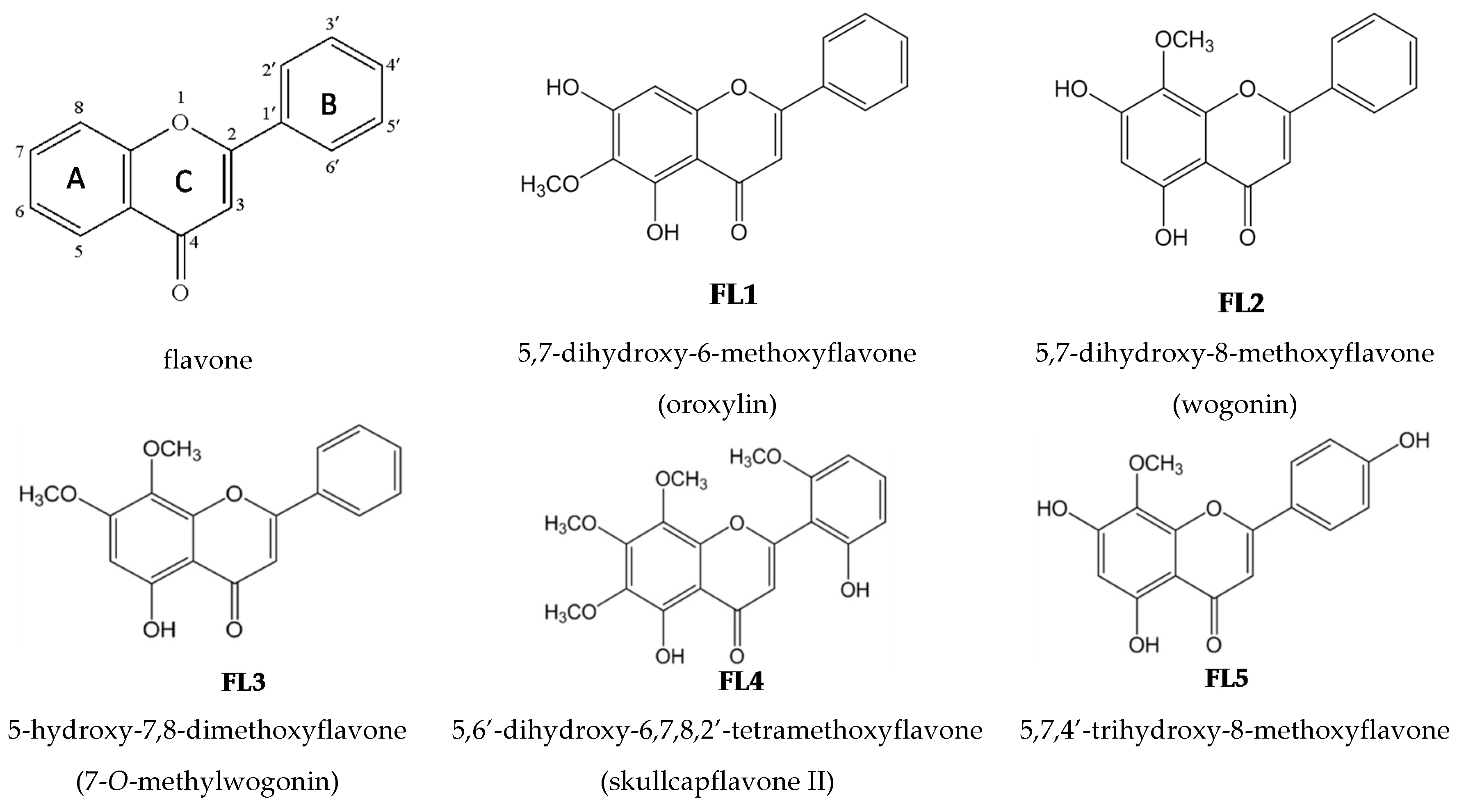
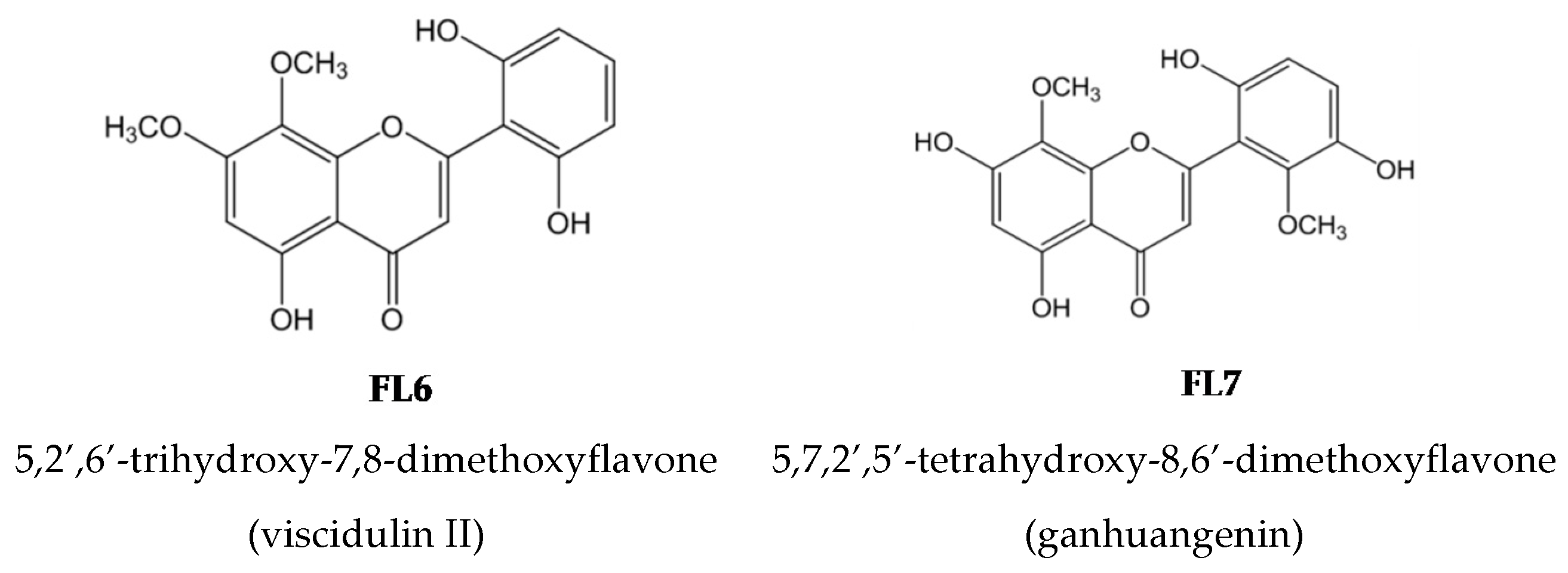
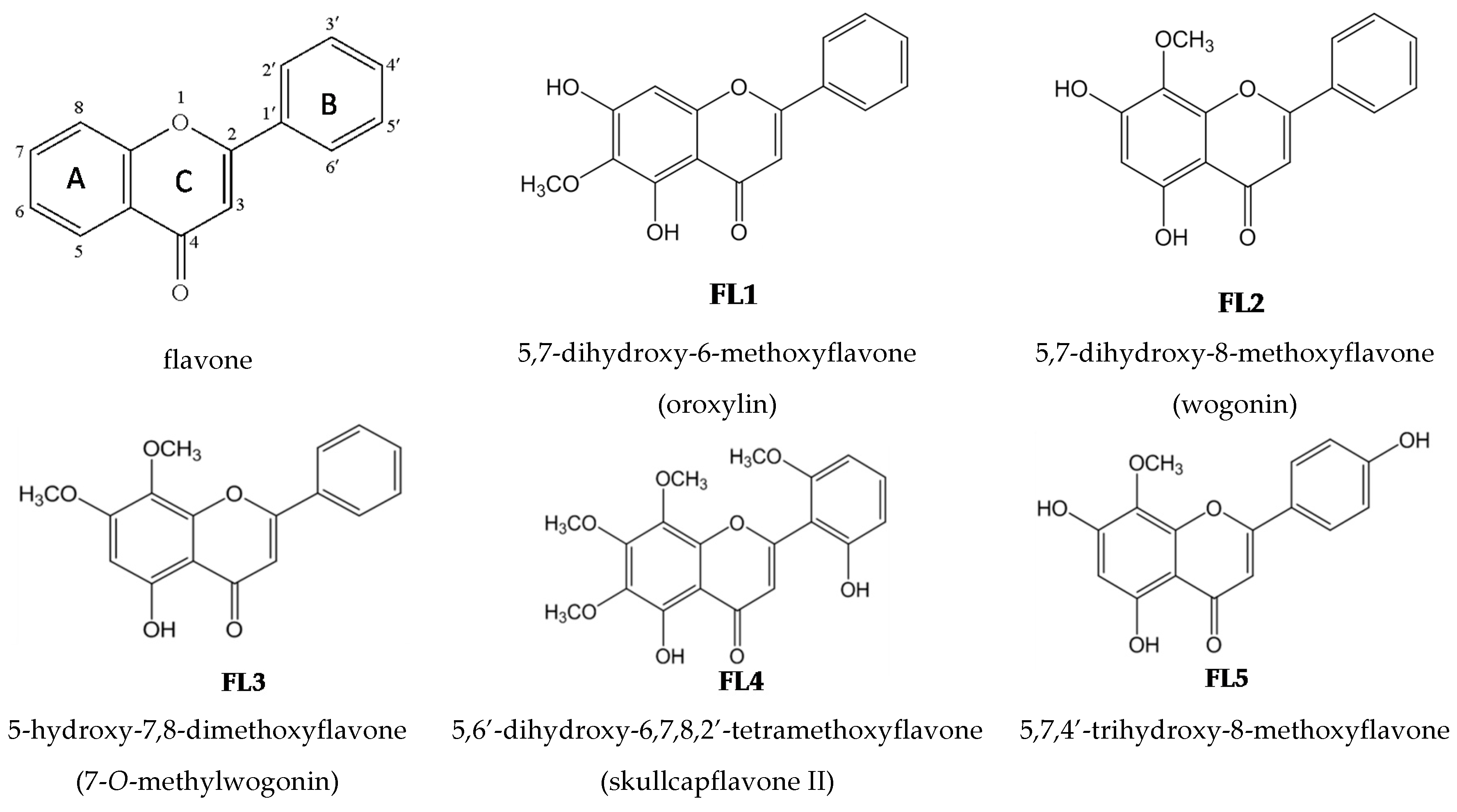
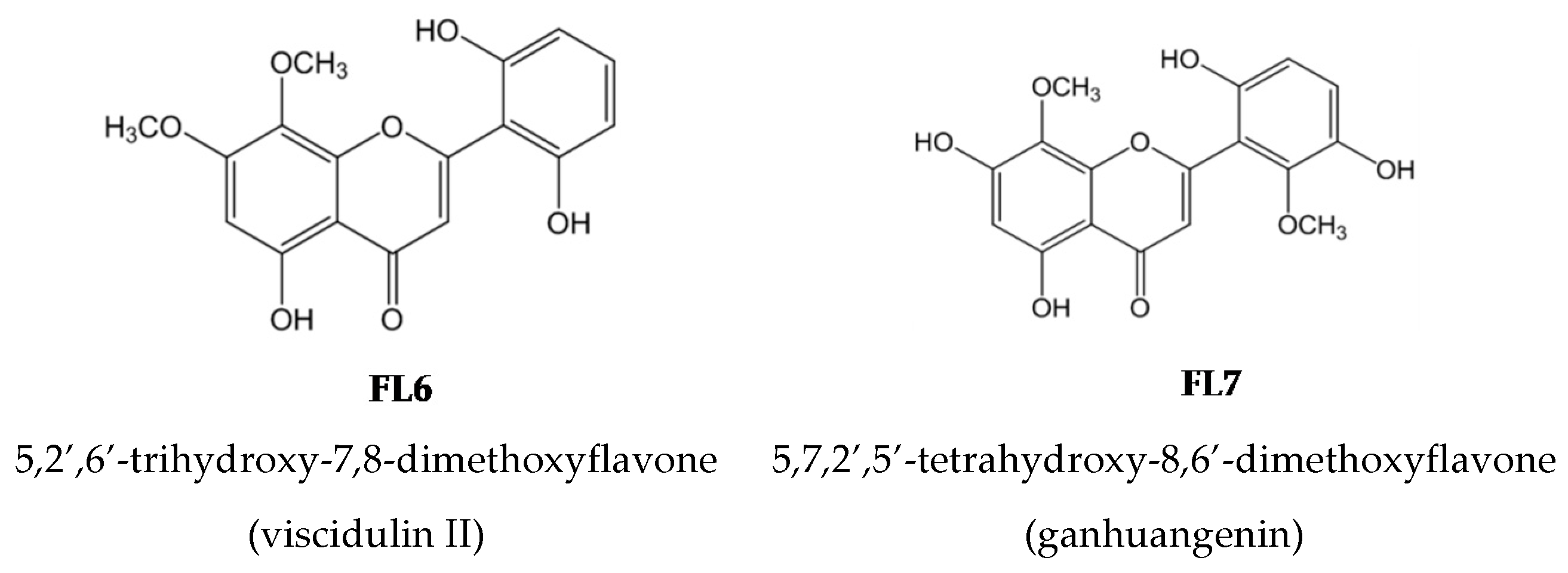
2.2. Effects of the Seven Flavones Isolated from EA Fraction of Scutellariae Radix on P. acnes-Induced IL-8 and IL-1β Production in Human Monocytic THP-1 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
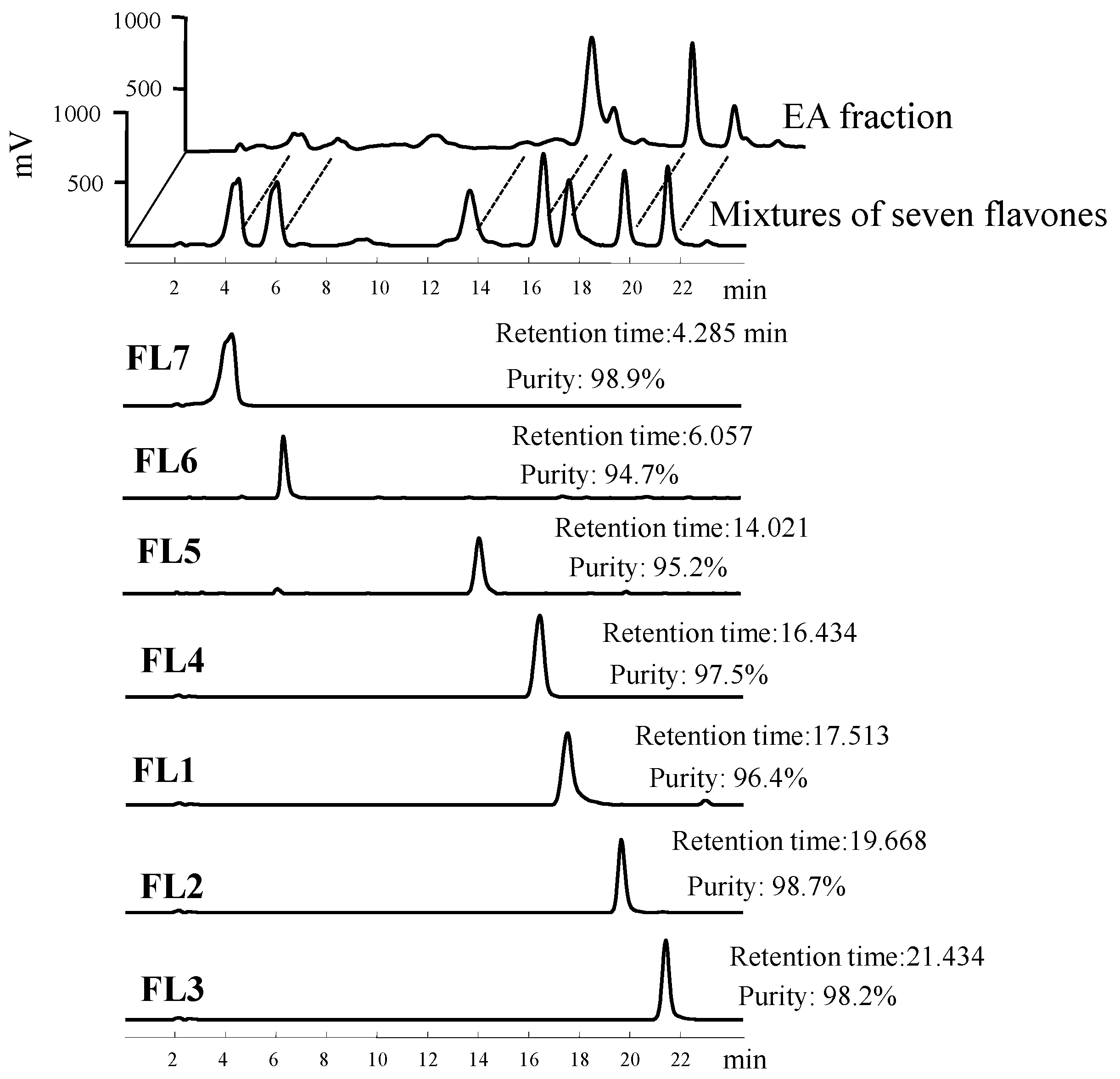
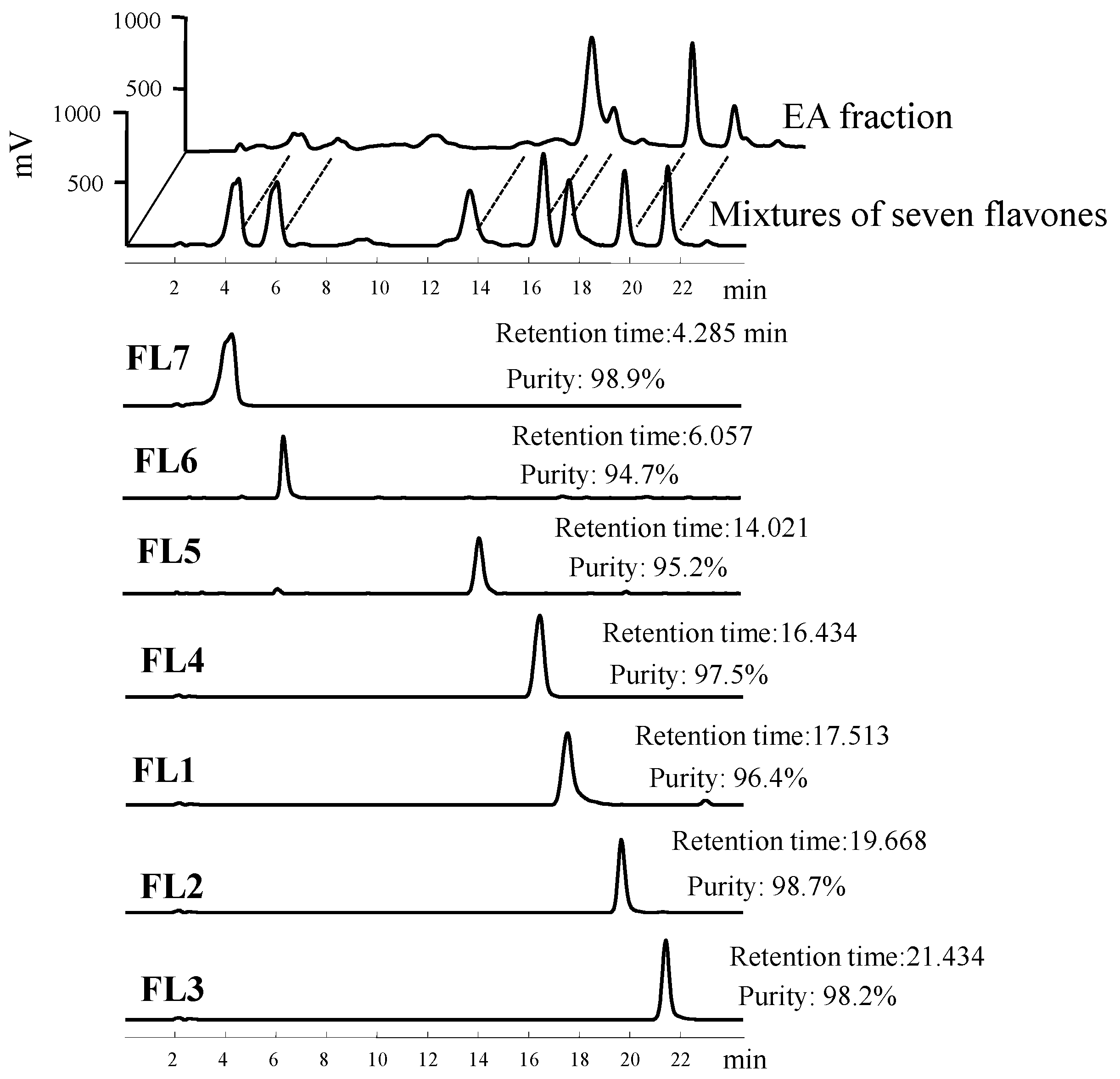{kind=link}
| Compound | Flavone (μM) | Cell Viability (% of Control) | IL-8 Level (ng/mL) | IC50 for IL-8 (μM) | IL-1β Level (ng/mL) | IC50 for IL-1β (μM) |
|---|---|---|---|---|---|---|
| Control | 0 | 93.2 ± 4.5 | 0.2 ± 0.2 ** | 0.008 ± 0.007 ** | ||
| DMSO | 0 | 100.0 ± 2.5 | 52.4 ± 5.1 | 2.4 ± 0.2 | ||
| FL1 | 5 | 101.8 ± 7.8 | 39.2 ± 3.8 * | 13.1 | 1.6 ± 0.2 ** | NA (>15) |
| 10 | 93.3 ± 4.0 | 29.3 ± 5.4 ** | 1.5 ± 0.2 ** | |||
| 15 | 97.2 ± 3.0 | 24.1 ± 2.5 ** | 1.2 ± 0.1 ** | |||
| 30 | 64.5 ± 3.0 | ND | ND | |||
| FL2 | 5 | 98.6 ± 12.1 | 55.8 ± 8.0 | 8.7 | 1.2 ± 0.1 ** | 4.9 |
| 10 | 112.0 ± 13.0 | 18.9 ± 3.5 ** | 0.6 ± 0.1 ** | |||
| 15 | 94.7 ± 2.1 | 5.9 ± 1.3 ** | 0.3 ± 0.1 ** | |||
| 30 | 88.4 ± 4.0 | ND | ND | |||
| FL3 | 30 | 99.9 ± 7.1 | 39.1 ± 12.0 * | 55.2 | 1.4 ± 0.1 ** | 72.8 |
| 60 | 101.0 ± 7.8 | 24.7 ± 7.4 ** | 1.4 ± 0.1 ** | |||
| 120 | 103.6 ± 4.5 | 15.8 ± 3.0 ** | 1.0 ± 0.1 ** | |||
| 240 | 65.6 ± 2.4 | ND | ND | |||
| FL4 | 5 | 102.8 ± 2.4 | 56.4 ± 1.9 | NA (>15) | 1.6 ± 0.1 ** | 9.1 |
| 10 | 98.8 ± 3.9 | 39.3 ± 1.6 * | 1.1 ± 0.1 ** | |||
| 15 | 99.6 ± 0.5 | 29.6 ± 2.2 ** | 1.0 ± 0.1 ** | |||
| 30 | 87.1 ± 1.8 | ND | ND | |||
| FL5 | 5 | 103.6 ± 2.0 | 67.7 ± 10.1 ** | 10.2 | 2.0 ± 0.1 * | 11.3 |
| 10 | 100.1 ± 2.0 | 32.4 ± 2.5 ** | 1.3 ± 0.2 ** | |||
| 15 | 100.8 ± 2.5 | 13.5 ± 0.6 ** | 0.8 ± 0.1 ** | |||
| 30 | 78.2 ± 2.8 | ND | ND | |||
| FL6 | 15 | 104.3 ± 5.8 | 41.5 ± 8.2 * | 26.1 | 2.0 ± 0.2 ** | NA (>60) |
| 30 | 101.3 ± 4.6 | 23.8 ± 1.5 ** | 1.6 ± 0.1 ** | |||
| 60 | 100.9 ± 4.2 | 20.3 ± 3.4 ** | 1.4 ± 0.1 ** | |||
| 120 | 87.5 ± 3.1 | ND | ND | |||
| FL7 | 60 | 106.7 ± 6.0 | 33.5 ± 2.3 ** | 124.3 | 1.7 ± 0.1 ** | 84.3 |
| 90 | 110.9 ± 1.1 | 28.6 ± 3.0 ** | 1.0 ± 0.1 ** | |||
| 120 | 120.7 ± 1.2 | 26.3 ± 1.2 ** | 0.8 ± 0.1 ** | |||
| 240 | 89.3 ± 3.2 | ND | ND |
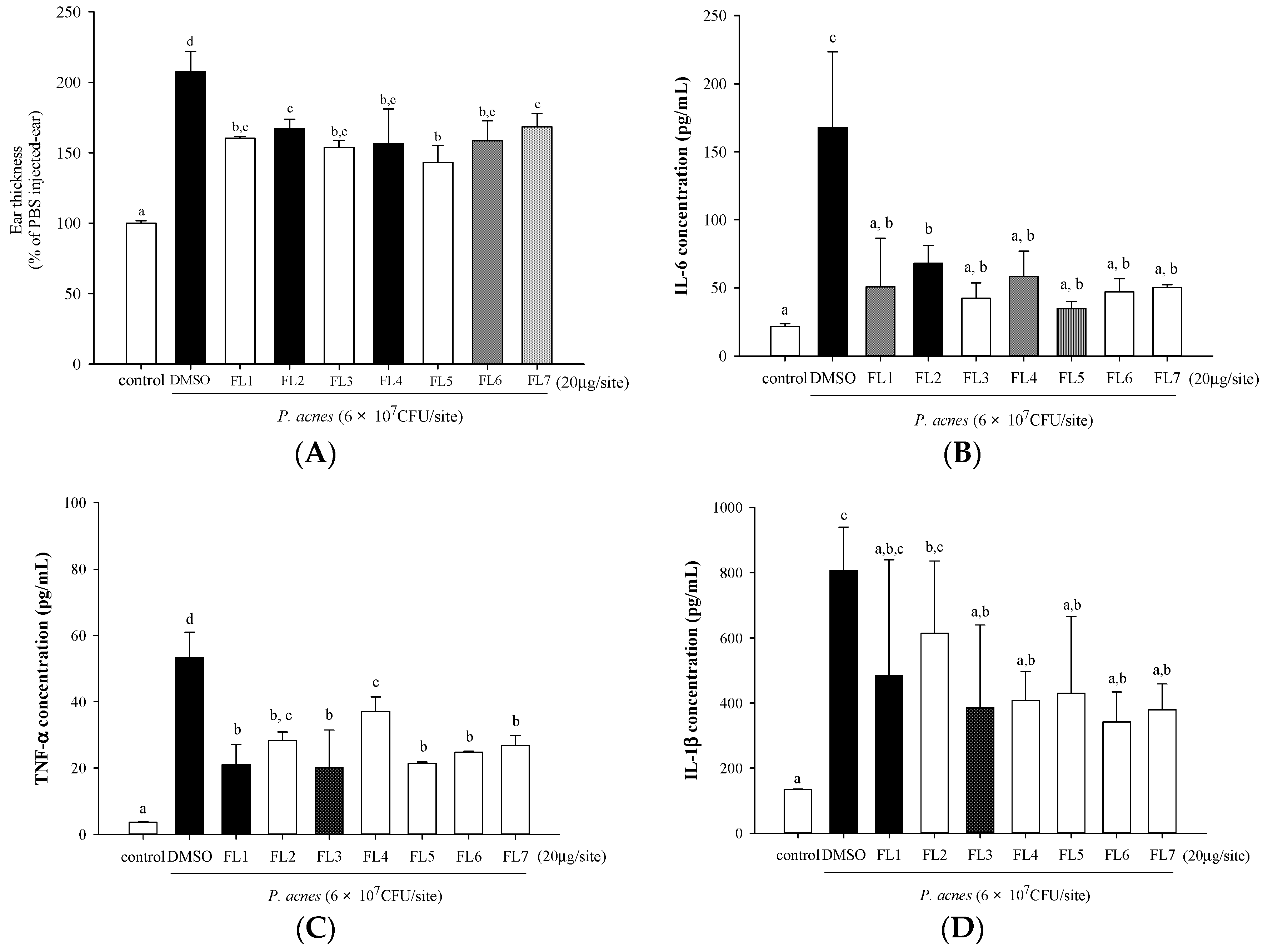
2.3. Effects of the Seven Flavones Isolated from Scutellariae Radix on P. acnes-Induced Ear Edema and Cytokine Production in Mouse Ear Homogenates

3. Discussion
4. Experimental Section
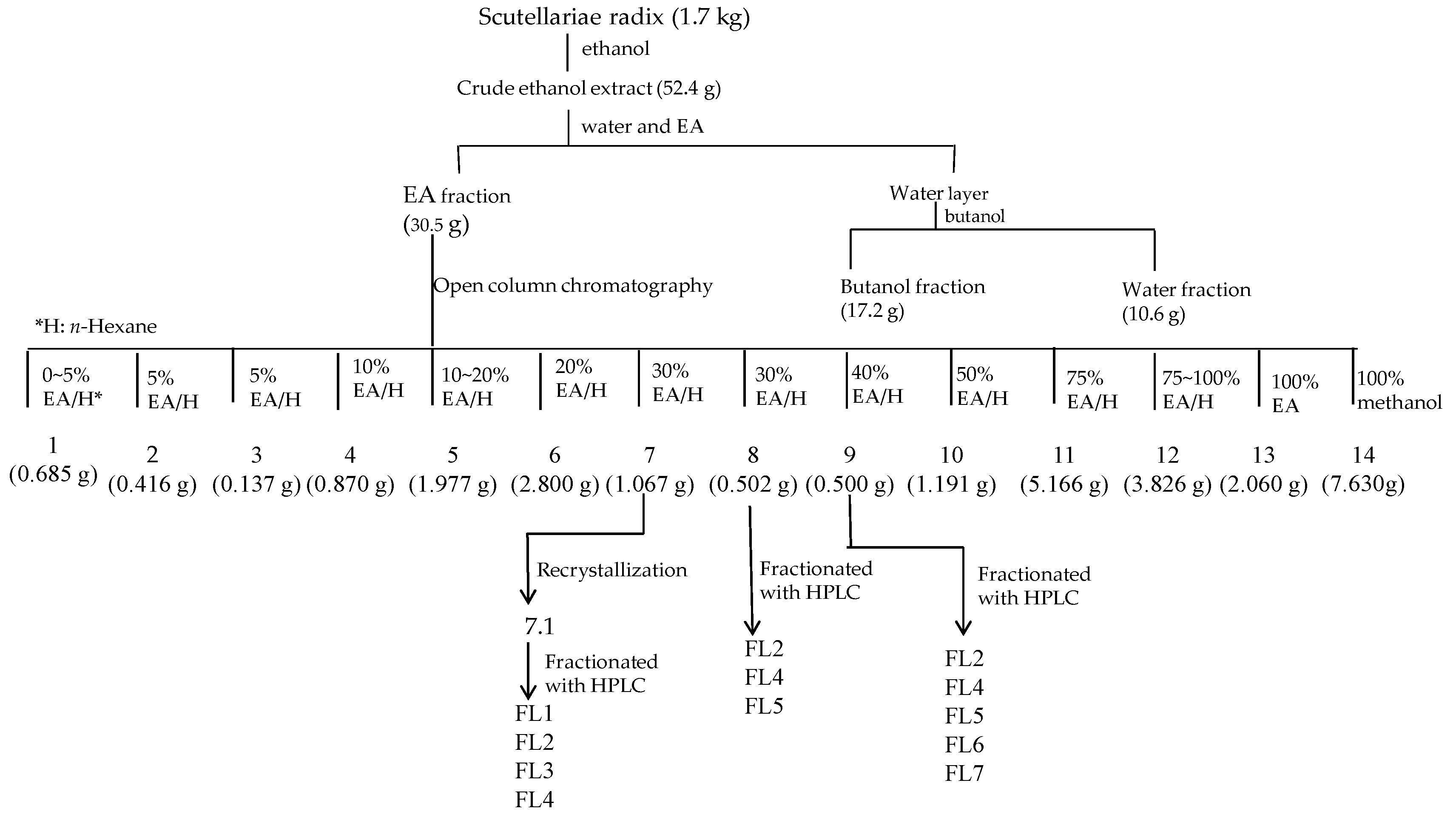
4.1. Isolation and Structural Elucidation

4.2. Culture of P. acnes and Preparation of Heat-Killed Bacteria
4.3. Determination of the Viability of THP-1 Cells
4.4. Measurement of Cytokine Production in Human Monocytic THP-1 Cells
4.5. P. acnes-Induced Ear Edema and Measurement of Cytokine Levels In Vivo
4.6. Measurement of Cytokine Levels In Vivo
4.7. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Farrar, M.D.; Ingham, E. Acne: Inflammation. Clin. Dermatol. 2004, 22, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Koreck, A.; Pivarcsi, A.; Dobozy, A.; Kemény, L. The role of innate immunity in the pathogenesis of acne. Dermatology 2003, 20, 96–105. [Google Scholar] [CrossRef]
- Kurokawa, I.; Danby, F.W.; Ju, Q.; Wang, X.; Xiang, L.F.; Xia, L.; Chen, W.; Nagy, I.; Picardo, M.; Suh, D.H.; et al. New developments in our understanding of acne pathogenesis and treatment. Exp. Dermatol. 2009, 18, 821–832. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, N.R.; Gilliland, K.L.; Zhao, W.; Liu, W.; Thiboutot, D.M. Gene array expression profiling in acne lesions reveals marked upregulation of genes involved in inflammation and matrix remodeling. J. Investig. Dermatol. 2006, 126, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Kistowska, M.; Gehrke, S.; Jankovic, D.; Kerl, K.; Fettelschoss, A.; Feldmeyer, L.; Fenini, G.; Kolios, A.; Navarini, A.; Ganceviciene, R.; et al. IL-1beta drives inflammatory responses to Propionibacterium acnes in vitro and in vivo. J. Investig. Dermatol. 2014, 134, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; He, X.; He, X.; Li, M.; Zhang, R.; Fan, P.; Zhang, Q.; Jia, Z. The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2010, 128, 279–313. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Jiang, Y.; Chen, F. Separation methods used for Scutellaria baicalensis active components. J. Chromatogr. B 2004, 812, 277–290. [Google Scholar] [CrossRef]
- Li, C.; Lin, G.; Zuo, Z. Pharmacological effects and pharmacokinetics properties of Radix Scutellariae and its bioactive flavones. Biopharm. Drug Dispos. 2011, 32, 427–445. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bauer, R.; Melchart, D.; Xiao, P.G.; Staudinger, A. Radix Scutellariae—Huangqin. In Chromatographic Fingerprint Analysis of Herbal Medicines; Wagner, H., Bauer, R., Melchart, D., Xiao, P.G., Staudinger, A., Eds.; Springer: Vienna, Austria, 2011; Volume 2, Chapter 63; pp. 755–765. [Google Scholar]
- Li, K.L.; Sheu, S.J. Determination of flavonoids and alkaloids in the scute-coptis herb couple by capillary electrophoresis. Anal. Chim. Acta 1995, 313, 113–120. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, L.; Lee, T.J. Oroxylin A inhibition of lipopolysaccharide-induced iNOS and COX-2 gene expression via suppression of nuclear factor-κB activation. Biochem. Pharmacol. 2000, 59, 1445–1457. [Google Scholar] [CrossRef]
- Song, X.; Chen, Y.; Sun, Y.; Lin, B.; Qin, Y.; Hui, H.; Li, Z.; You, Q.; Lu, N.; Guo, Q. Oroxylin A, a classical natural product, shows a novel inhibitory effect on angiogenesis induced by lipopolysaccharide. Pharmacol. Rep. 2012, 64, 1189–1199. [Google Scholar] [CrossRef]
- Kubo, M.; Matsuda, H.; Tanaka, M.; Kimura, Y.; Okuda, H.; Higashino, M.; Tani, T.; Namba, K.; Arichi, S. Studies on Scutellaria radix. VII. Anti-arthritic and anti-inflammatory actions of methanol extract and flavonoid components from Scutellaria radix. Chem. Pharm. Bull. 1984, 32, 2724–2729. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, K.; Takido, M.; Takeuchi, M.; Nakagawa, S. Effect of chemical constituents from plants on 12-O-tetradecanoylphorbol-13-acetate induced inflammation in mice. Chem. Pharm. Bull. 1989, 37, 1071–1073. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.S.; Lim, H.; Park, H.; Kim, H.P. Effects of wogonin, a plant flavone from Scutellaria radix, on skin inflammation: In vivo regulation of inflammation-associated gene expression. Biochem. Pharmacol. 2003, 66, 1271–1278. [Google Scholar] [CrossRef]
- Chandrasekaran, C.V.; Thiyagarajan, P.; Deepak, H.B.; Agarwal, A. In vitro modulation of LPS/calcimycin induced inflammatory and allergic mediators by pure compounds of Andrographis paniculata (King of bitters) extract. Int. Immunopharmacol. 2011, 11, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.W.; Kuo, Y.H.; Lin, B.F. Anti-inflammatory activity of new compounds from Andrographis paniculata by NF-κB transactivation inhibition. J. Agric. Food Chem. 2010, 58, 2505–2512. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.Y.; Ahn, K.S.; Park, M.J.; Kwon, O.K.; Lee, H.K.; Oh, S.R. Skullcapflavone II inhibits ovalbumin-induced airway inflammation in a mouse model of asthma. Int. Immunopharmacol. 2012, 12, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Suzuki, Y.; Tomimori, T.; Yamada, H. Antiviral activity of plant flavonoid, 5,7,4′-trihydroxy-8-methoxyflavone, from the roots of Scutellaria baicalensis against influenza A (H3N2) and B viruses. Biol. Pharm. Bull. 1995, 18, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, T.; Ohmoto, T.; Sankawa, U.; Tomimori, T.; Miyaichi, Y.; Imoto, Y. Inhibition of adenosine 3′,5′-cyclic monophosphate phosphodiesterase by flavonoids. II. Chem. Pharm. Bull. 1988, 36, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.O. Effect of ganhuangenin obtained from Scutellaria radix on the chemical mediator production of peritoneal exudate cells and immunoglobulin E level of mesenteric lymph node lymphocytes in Sprague-Dawley rats. Phytother. Res. 2002, 16, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Clahsen, T.; Schaper, F. Interleukin-6 acts in the fashion of a classical chemokine on monocytic cells by inducing integrin activation, cell adhesion, actin polymerization, chemotaxis, and transmigration. J. Leukoc. Biol. 2008, 84, 1521–1529. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.; Sansonetti, P.J. IL-8 is a key chemokine regulating neutrophil recruitment in a new mouse model of Shigella-induced colitis. J. Immunol. 2004, 173, 4197–4206. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Tsai, T.H.; Huang, C.J.; Li, Y.Y.; Chyuan, J.H.; Chuang, L.T.; Tsai, P.J. Inhibitory effects of wild bitter melon leaf extract on Propionibacterium acnes-induced skin inflammation in mouse and cytokine production in vitro. Food Funct. 2015, 6, 2550–2560. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.H.; Chien, P.Y.; Yang, C.H.; Lee, A.R. Novel synthesis of flavonoids of Scutellaria baicalensis Georgi. Chem. Pharm. Bull. 2003, 51, 339–340. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.L.; Ou, J.C.; Chen, C.F.; Kuo, Y.H. Flavonoids from the roots of Scutellaria luzonica Rolfe. J. Chin. Chem. Soc. 1991, 38, 619–623. [Google Scholar] [CrossRef]
- Hua, Y.; Wang, H.Q. Chemical components of Anaphalis sinica Hance. J. Chin. Chem. Soc. 2004, 51, 409–415. [Google Scholar] [CrossRef]
- Furukawa, M.; Suzuki, H.; Makino, M.; Ogawa, S.; Iida, T.; Fujimoto, Y. Studies on the constituents of Lagochilus leiacanthus (Labiatae). Chem. Pharm. Bull. 2011, 59, 1535–1540. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Kim, H.P.; Park, H. Structure and antiinflammatory activity relationships of wogonin derivatives. Arch. Pharm. Res. 2005, 28, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Stevens, J.F.; Wollenweber, E.; Ivancic, M.; Hsu, V.L.; Sundberg, S.; Deinzer, M.L. Leaf surface flavonoids of Chrysothamnus. Phytochemistry 1999, 51, 771–780. [Google Scholar] [CrossRef]
- Tanka, T.; Iinuma, M.; Mizuno, M. Synthesis of flavonoids in Scutellaria spp. II. Synthesis of 2′,6′-dioxygenated flavones. Yakugaku Zasshi 1987, 107, 827–829. [Google Scholar]
- Iinuma, M.; Tanaka, T.; Mizuno, M. Flavonoids synthesis II. Synthesis of flavones with a 2′,3′,6′-trioxygenated ring B. Chem. Pharm. Bull. 1985, 33, 4034–4036. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, P.-J.; Huang, W.-C.; Hsieh, M.-C.; Sung, P.-J.; Kuo, Y.-H.; Wu, W.-H. Flavones Isolated from Scutellariae radix Suppress Propionibacterium Acnes-Induced Cytokine Production In Vitro and In Vivo. Molecules 2016, 21, 15. https://doi.org/10.3390/molecules21010015
Tsai P-J, Huang W-C, Hsieh M-C, Sung P-J, Kuo Y-H, Wu W-H. Flavones Isolated from Scutellariae radix Suppress Propionibacterium Acnes-Induced Cytokine Production In Vitro and In Vivo. Molecules. 2016; 21(1):15. https://doi.org/10.3390/molecules21010015
Chicago/Turabian StyleTsai, Po-Jung, Wen-Cheng Huang, Ming-Chi Hsieh, Ping-Jyun Sung, Yueh-Hsiung Kuo, and Wen-Huey Wu. 2016. "Flavones Isolated from Scutellariae radix Suppress Propionibacterium Acnes-Induced Cytokine Production In Vitro and In Vivo" Molecules 21, no. 1: 15. https://doi.org/10.3390/molecules21010015
APA StyleTsai, P.-J., Huang, W.-C., Hsieh, M.-C., Sung, P.-J., Kuo, Y.-H., & Wu, W.-H. (2016). Flavones Isolated from Scutellariae radix Suppress Propionibacterium Acnes-Induced Cytokine Production In Vitro and In Vivo. Molecules, 21(1), 15. https://doi.org/10.3390/molecules21010015