
Effects of Fusarium solani and F. oxysporum Infection on the Metabolism of Ginsenosides in American Ginseng Roots

 and
and
Abstract
:1. Introduction
2. Results and Discussion
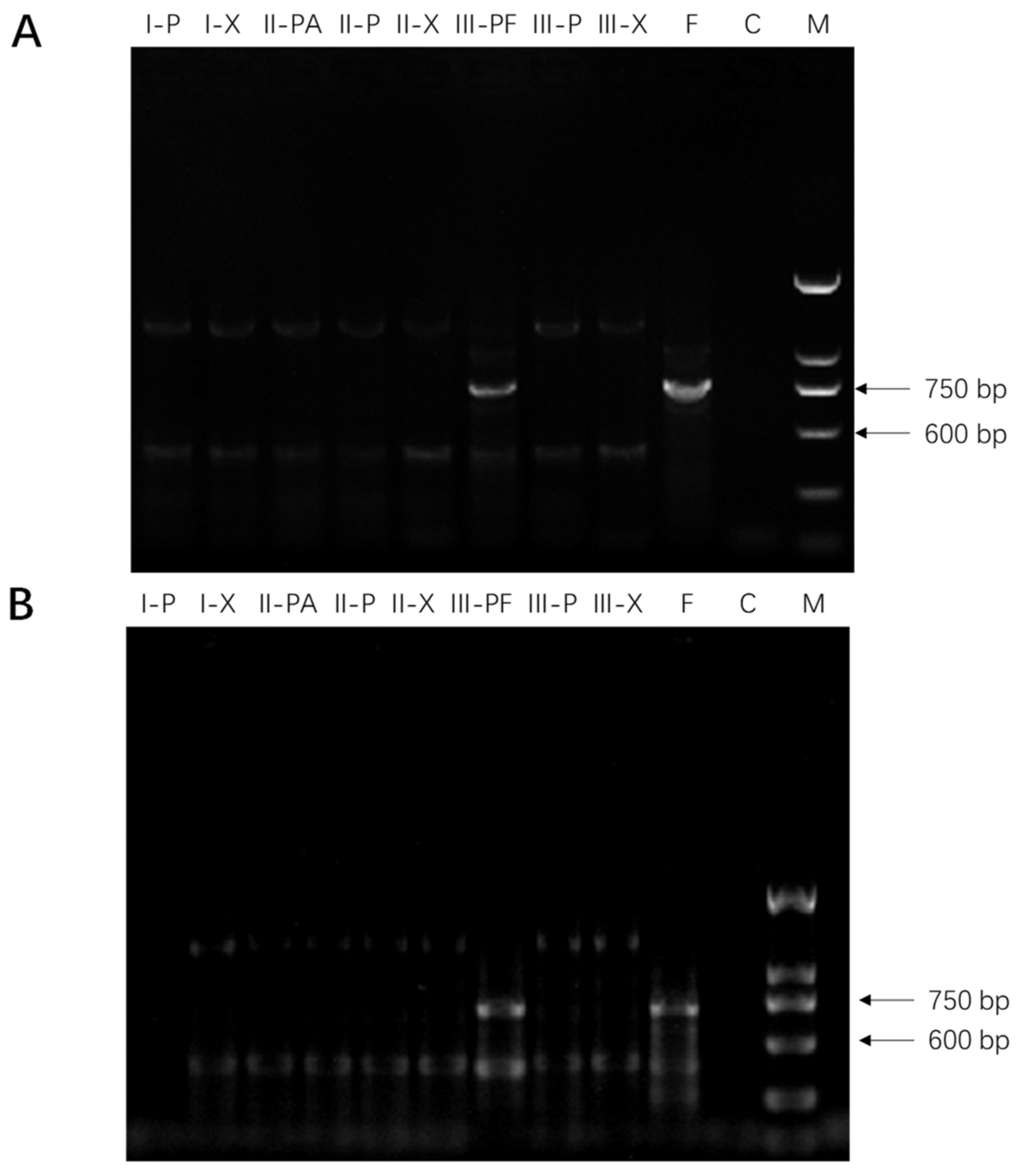
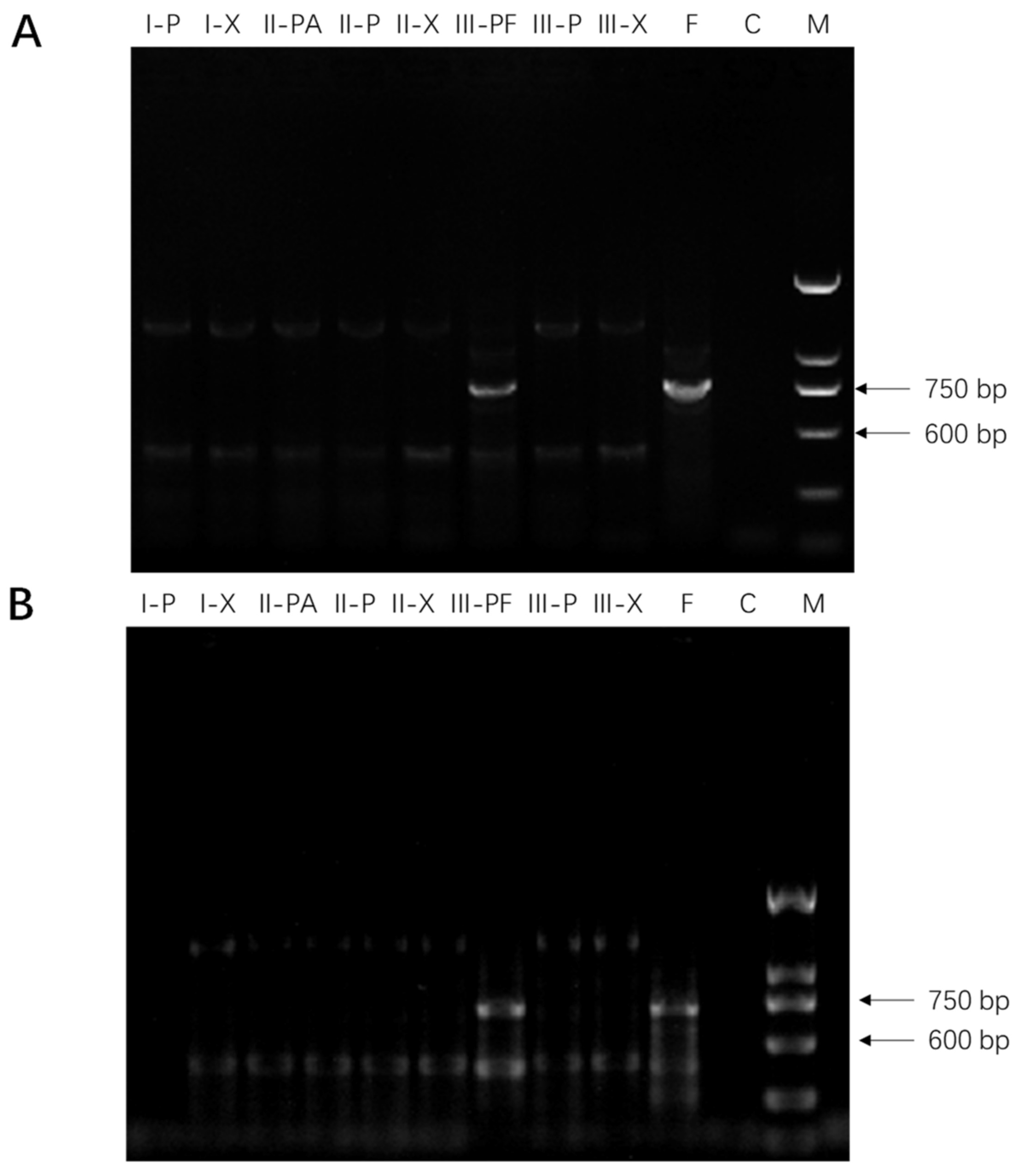
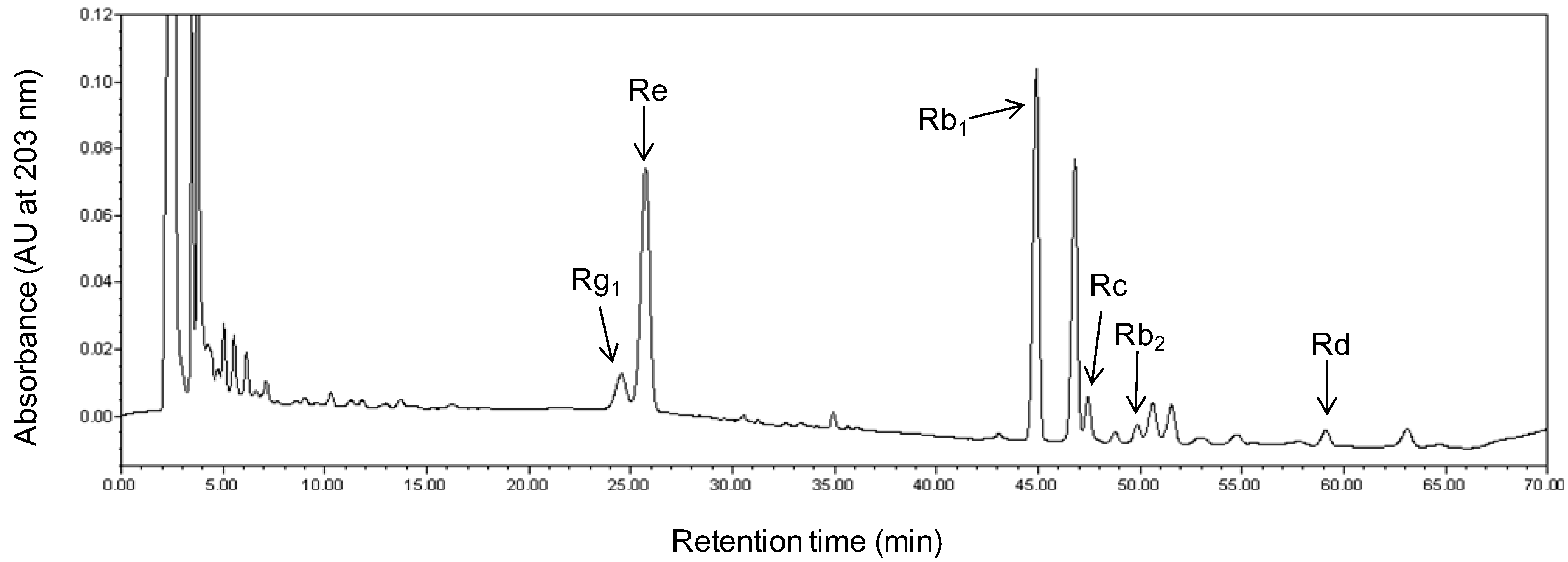
2.1. Molecular Detection of Fusarium spp. in American Ginseng Roots
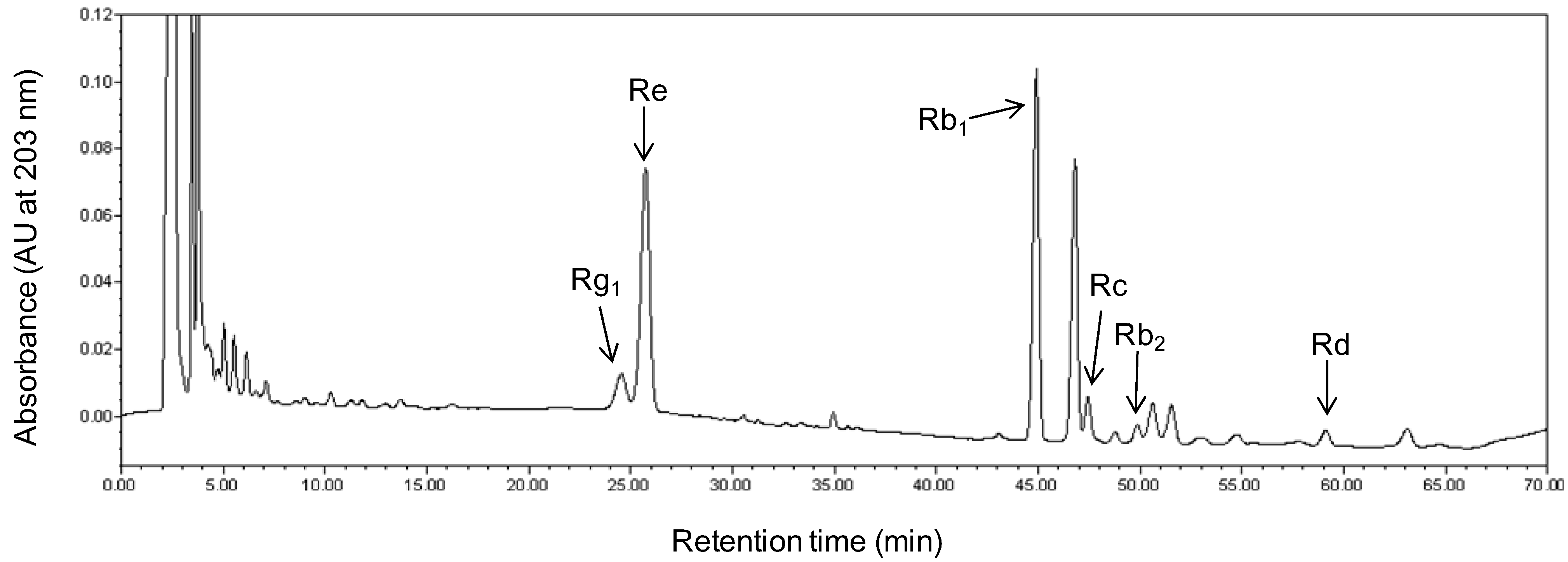
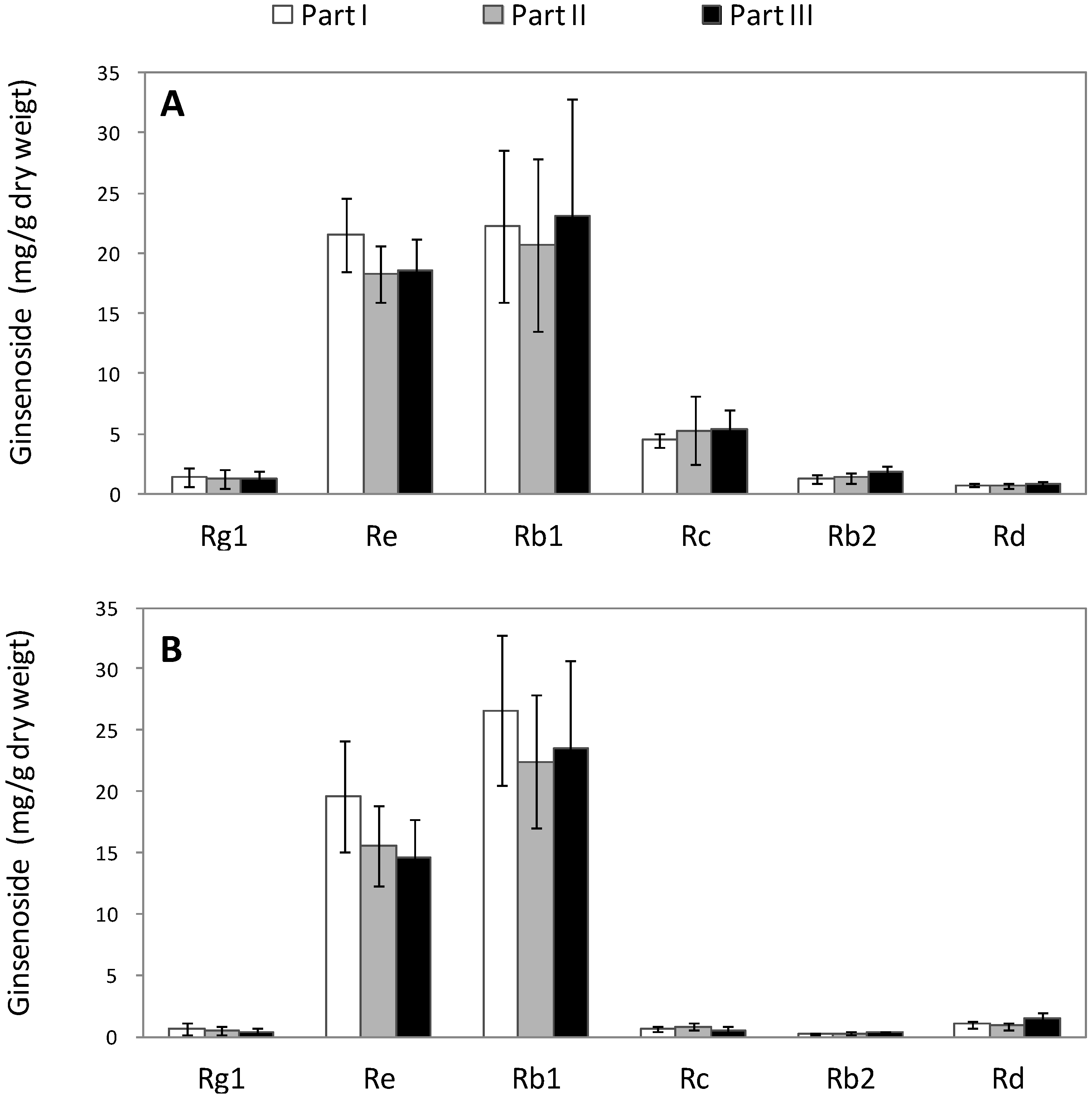
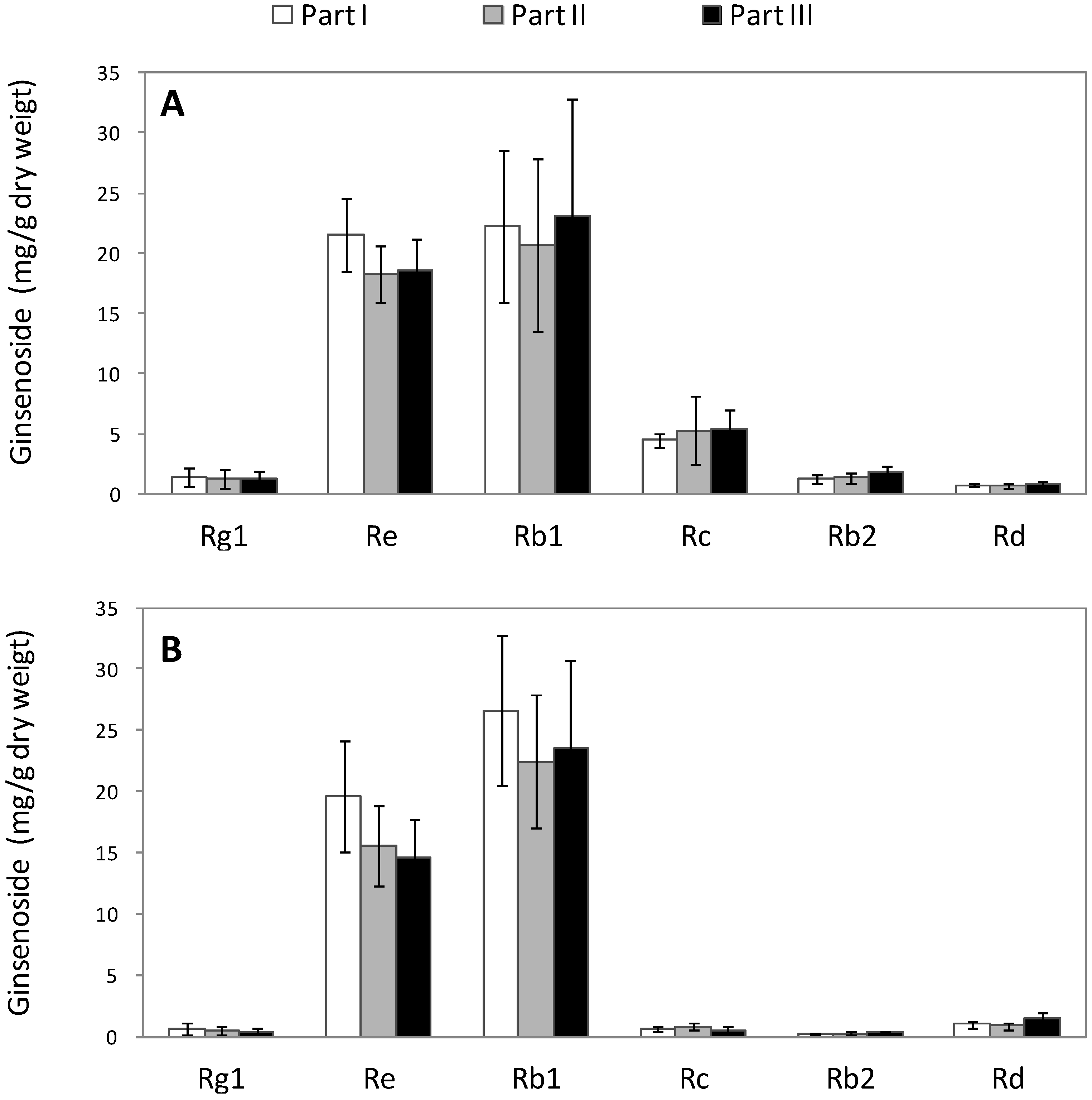
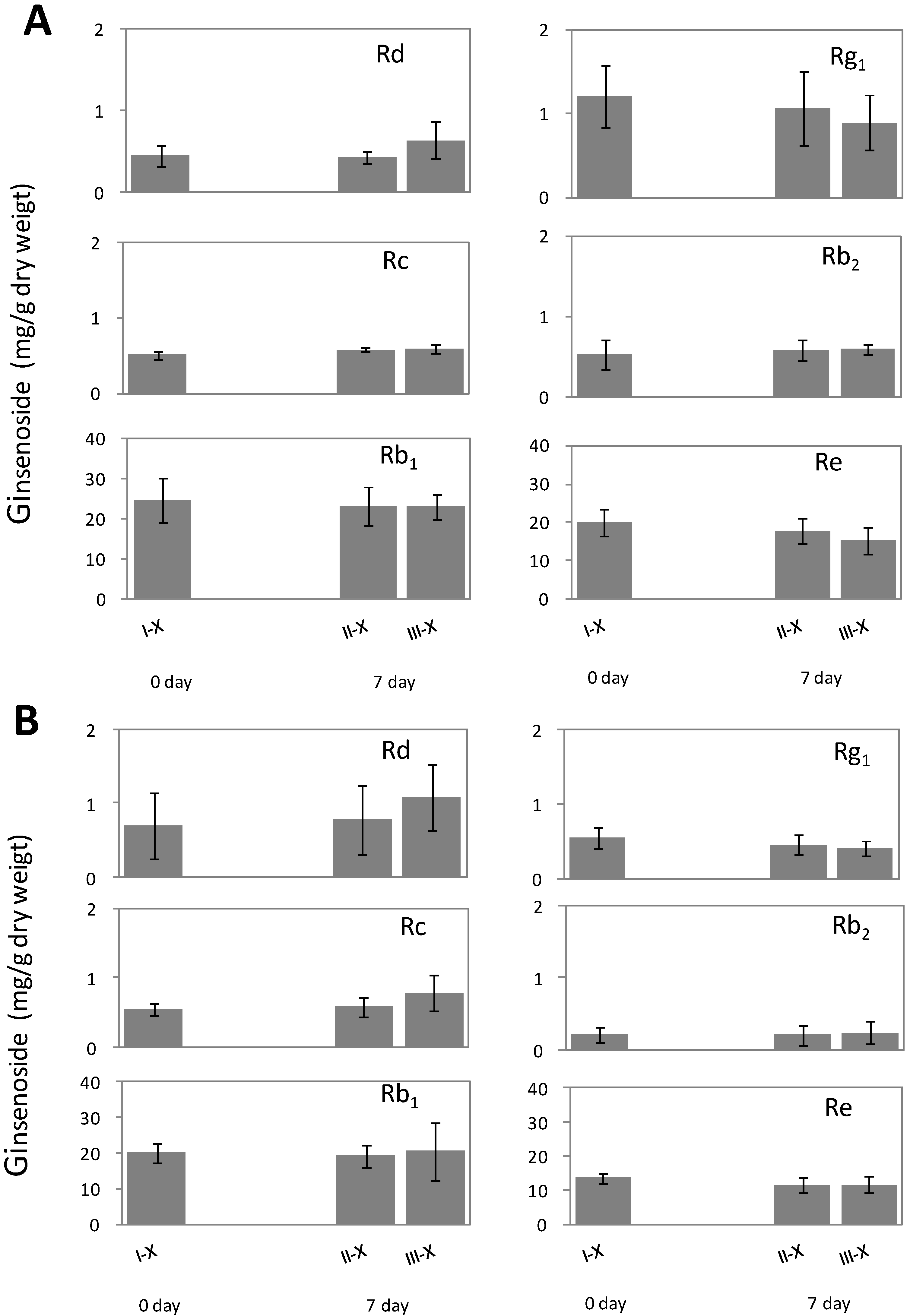
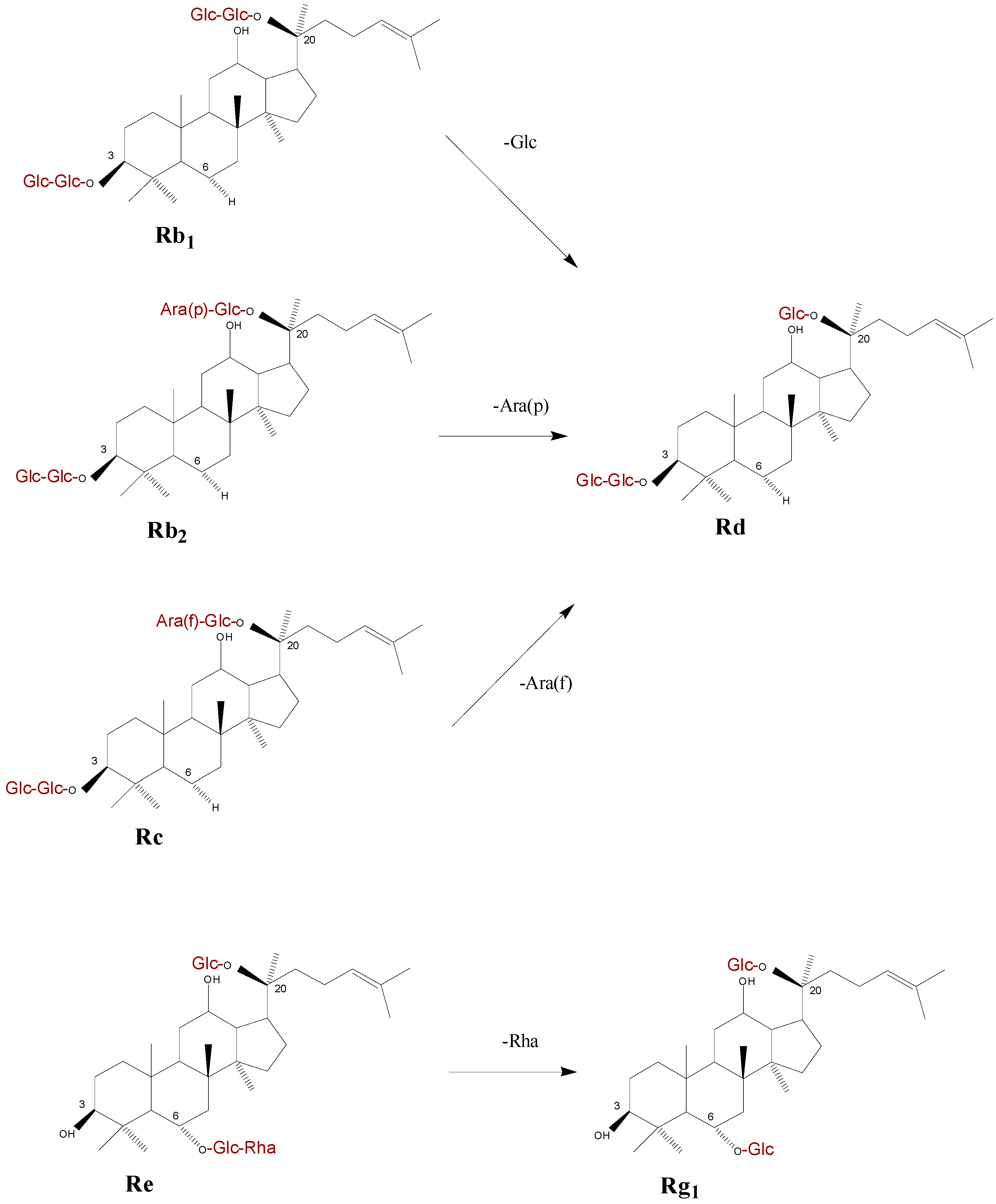
2.2. Alteration of Ginsenoside in American Ginseng Roots Inoculated with Fusarium spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rg1 | Re | Rb1 | Rc | Rb2 | Rd | |
|---|---|---|---|---|---|---|
| Part | 0.698 | 0.051 | 0.659 | 0.656 | 0.071 | 0.079 |
| Tissue type | 0.001 | 0.011 | 0.406 | 0.000 | 0.000 | 0.005 |
| Interaction between part and tissue type | 0.998 | 0.898 | 0.831 | 0.676 | 0.235 | 0.398 |
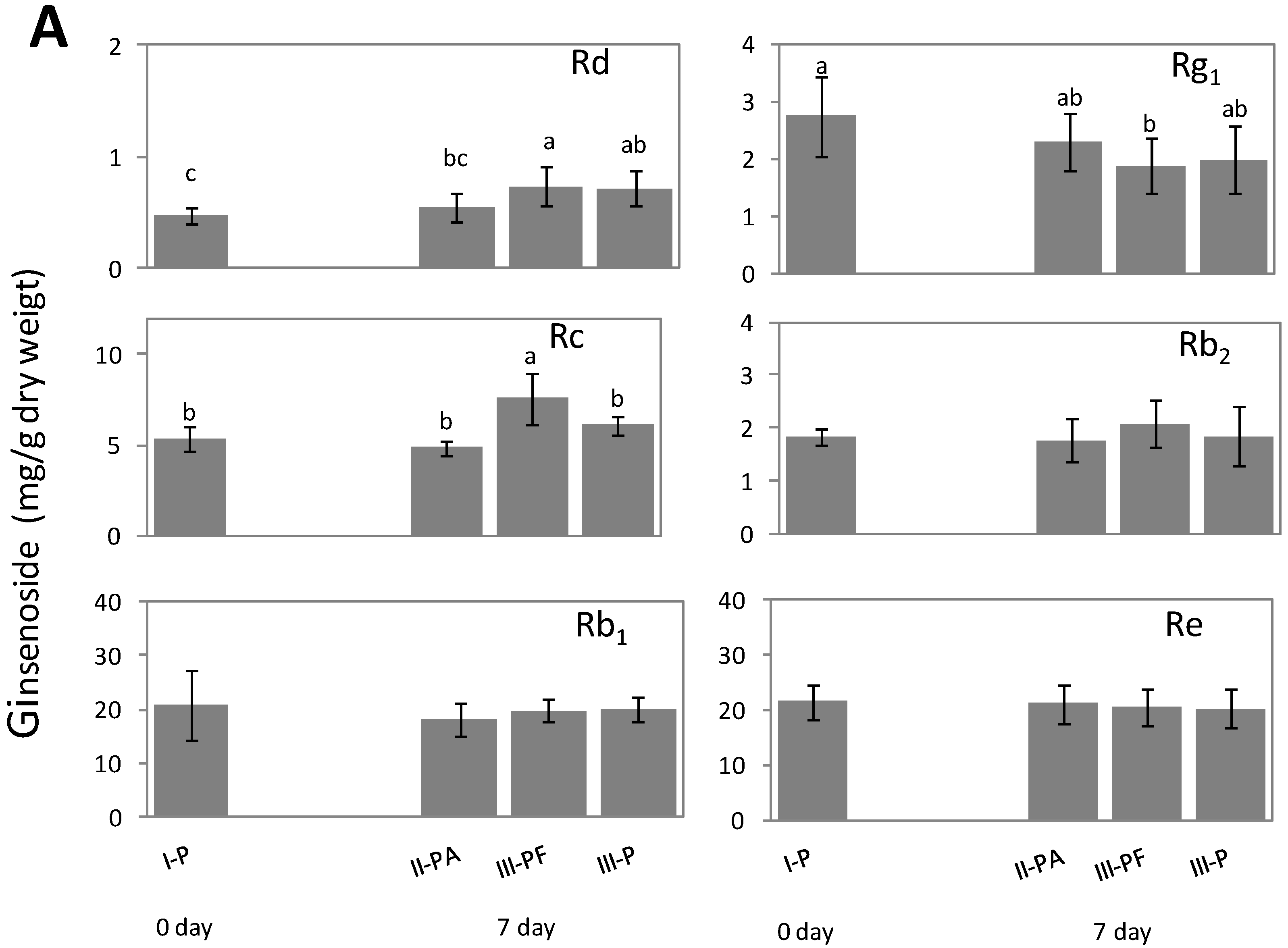
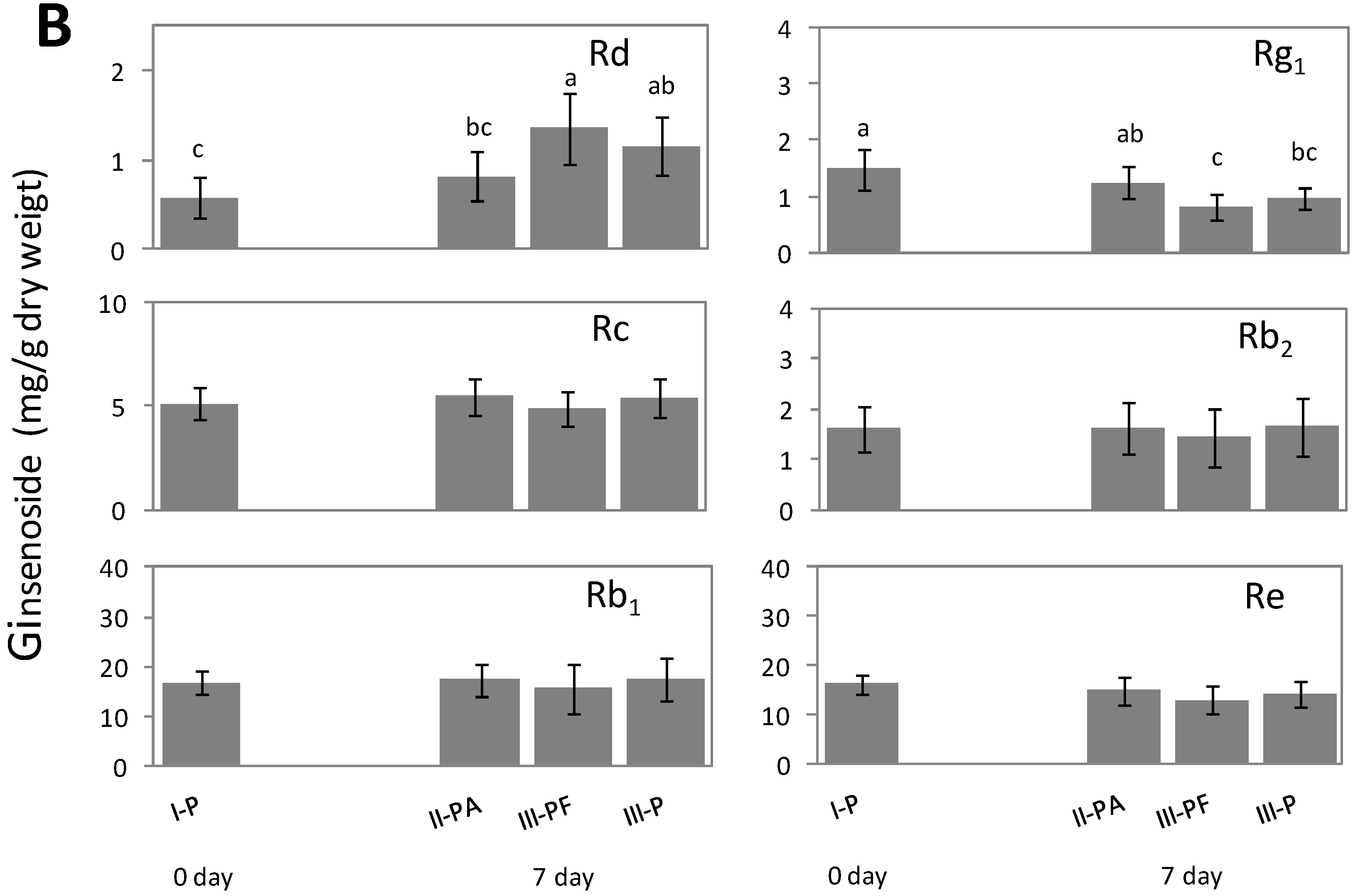
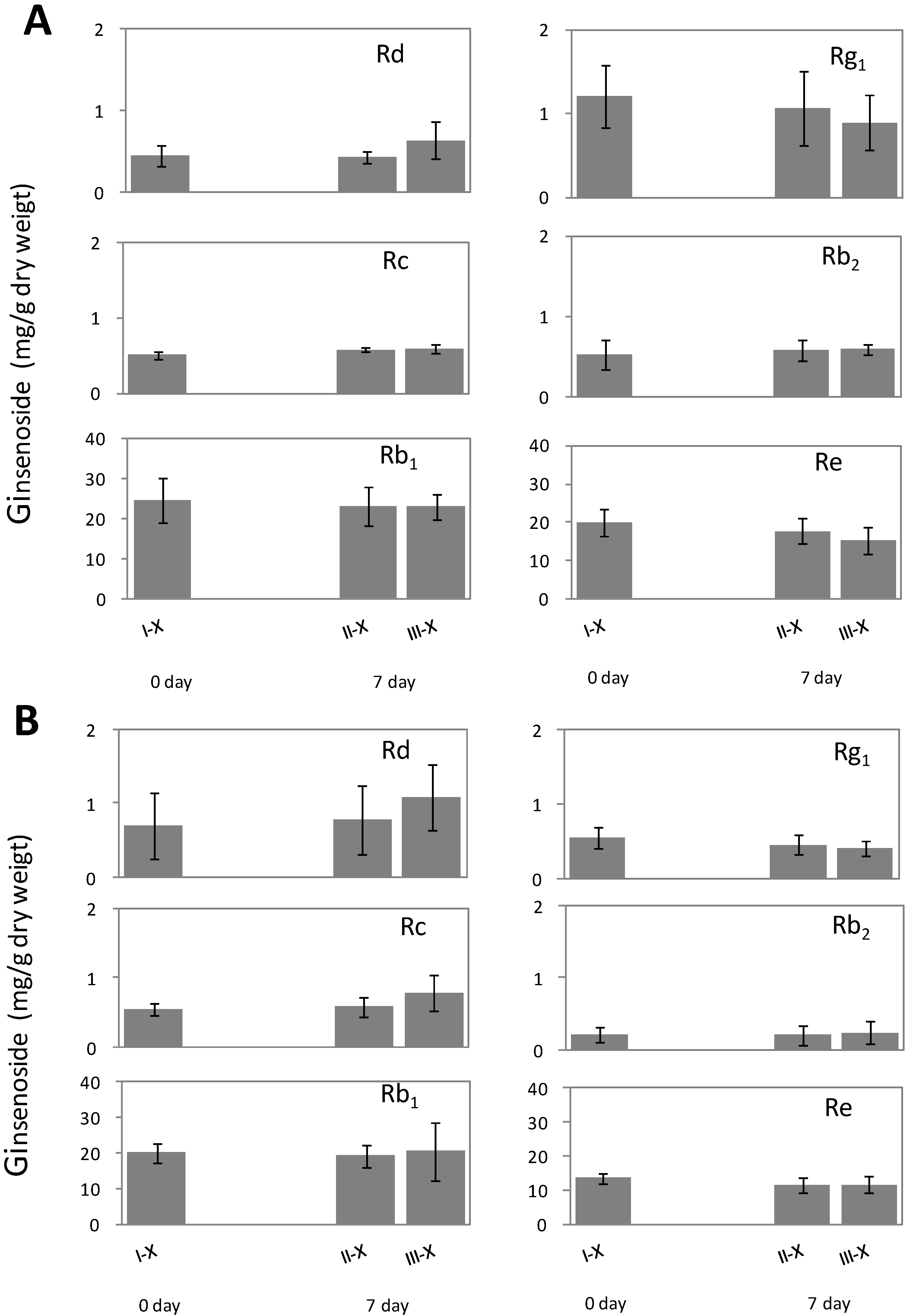
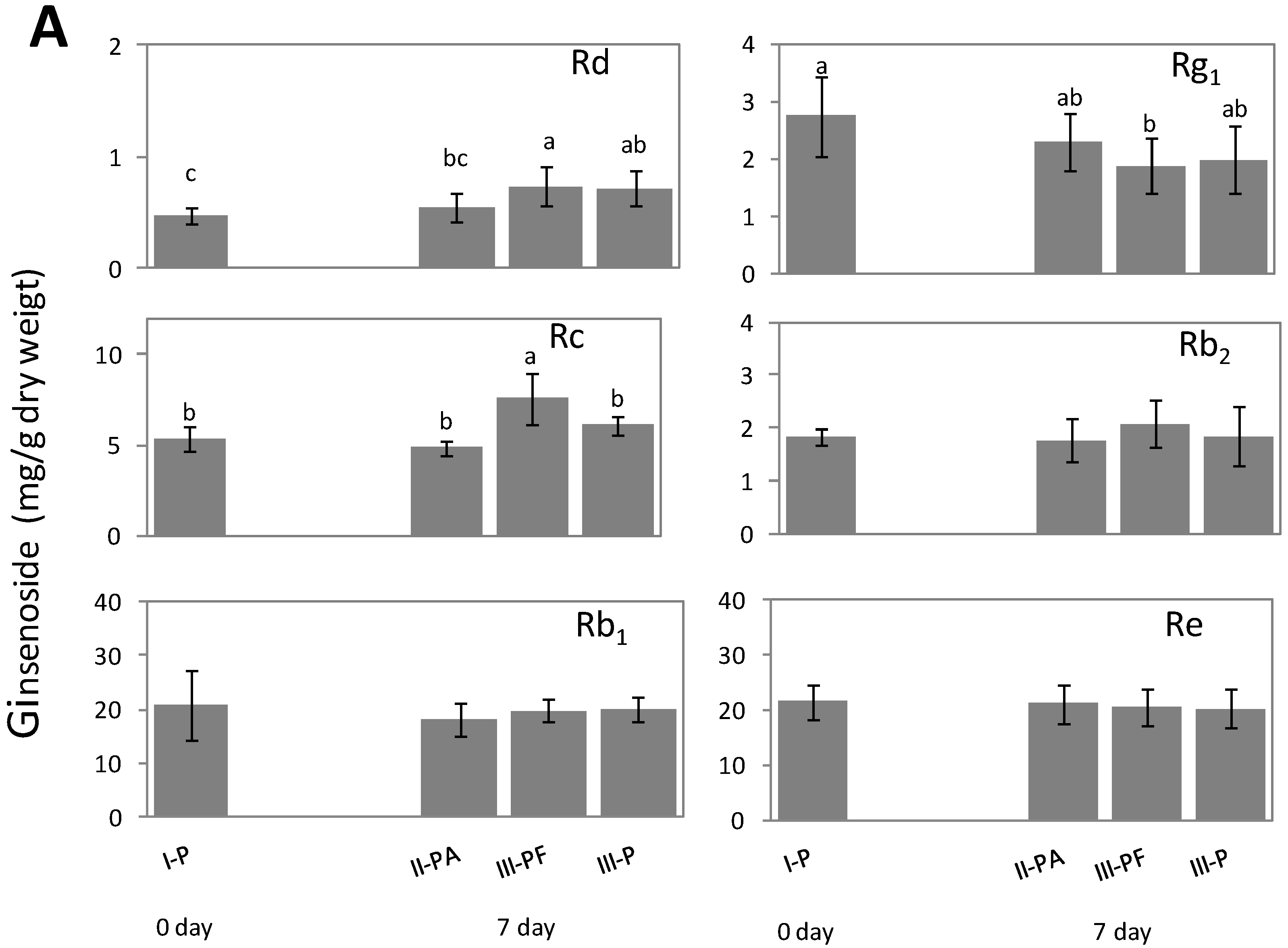
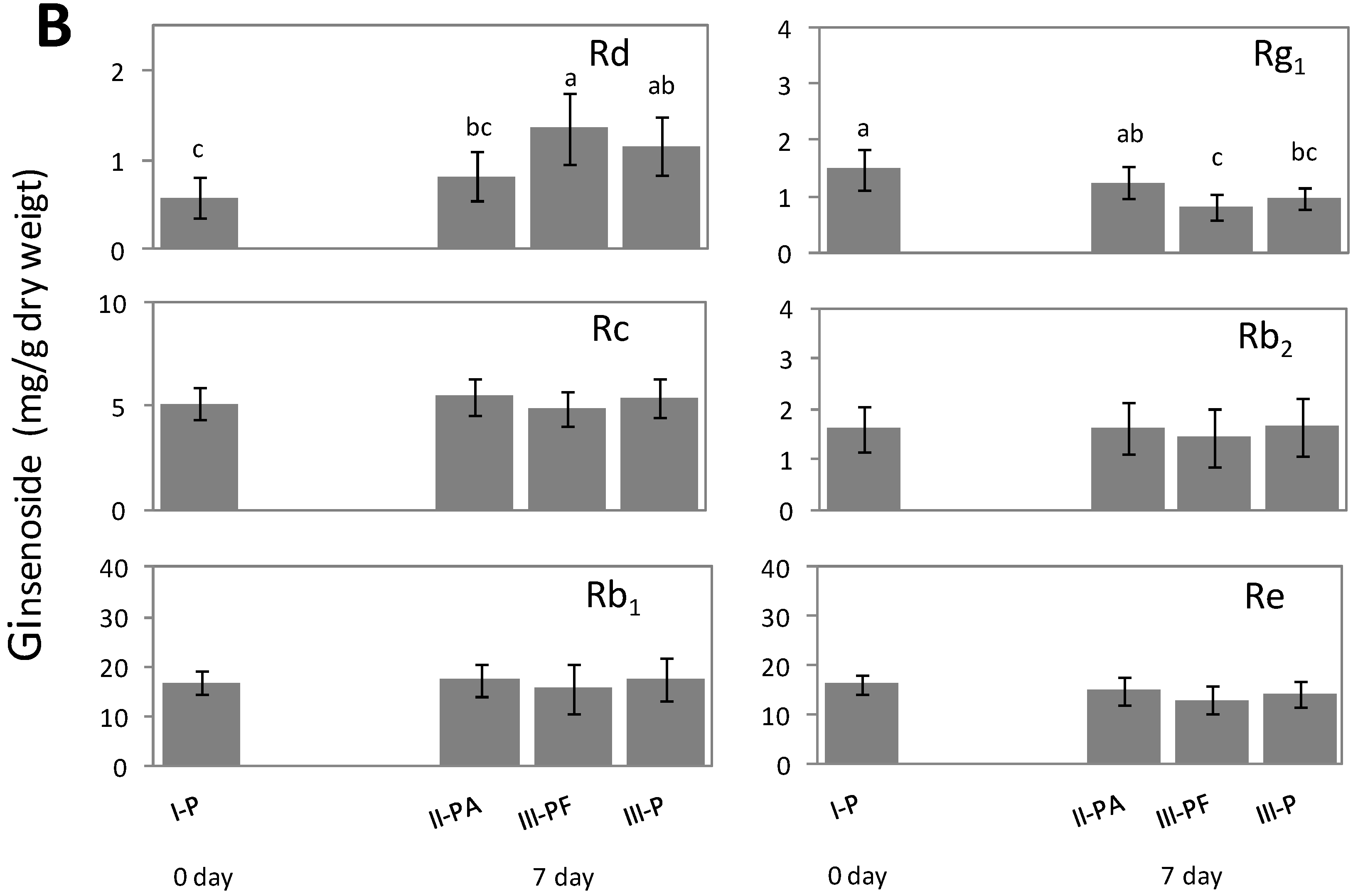
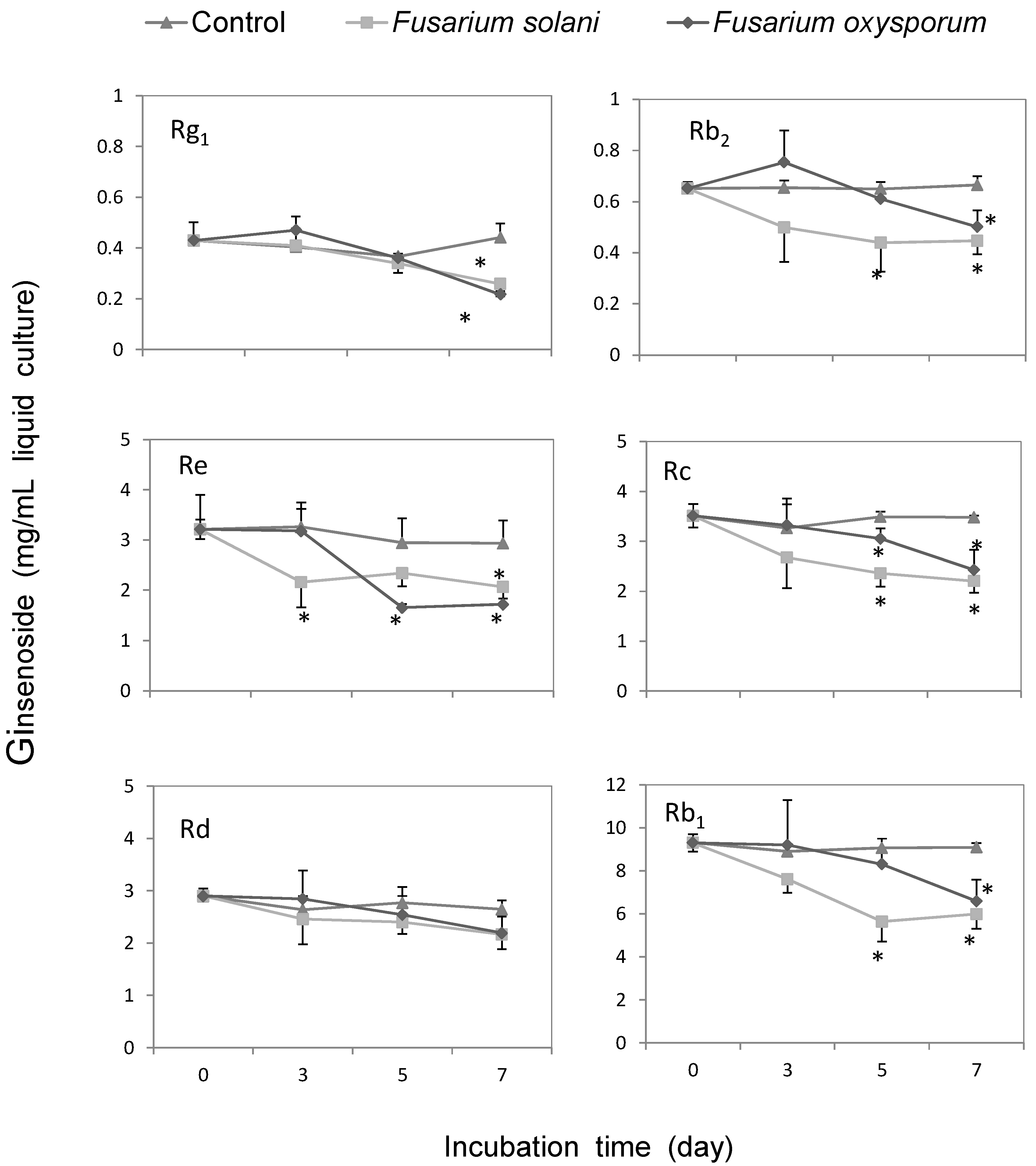
2.3. Metabolic Effect of Fusarium spp. on the Concentrations of Ginsenoside
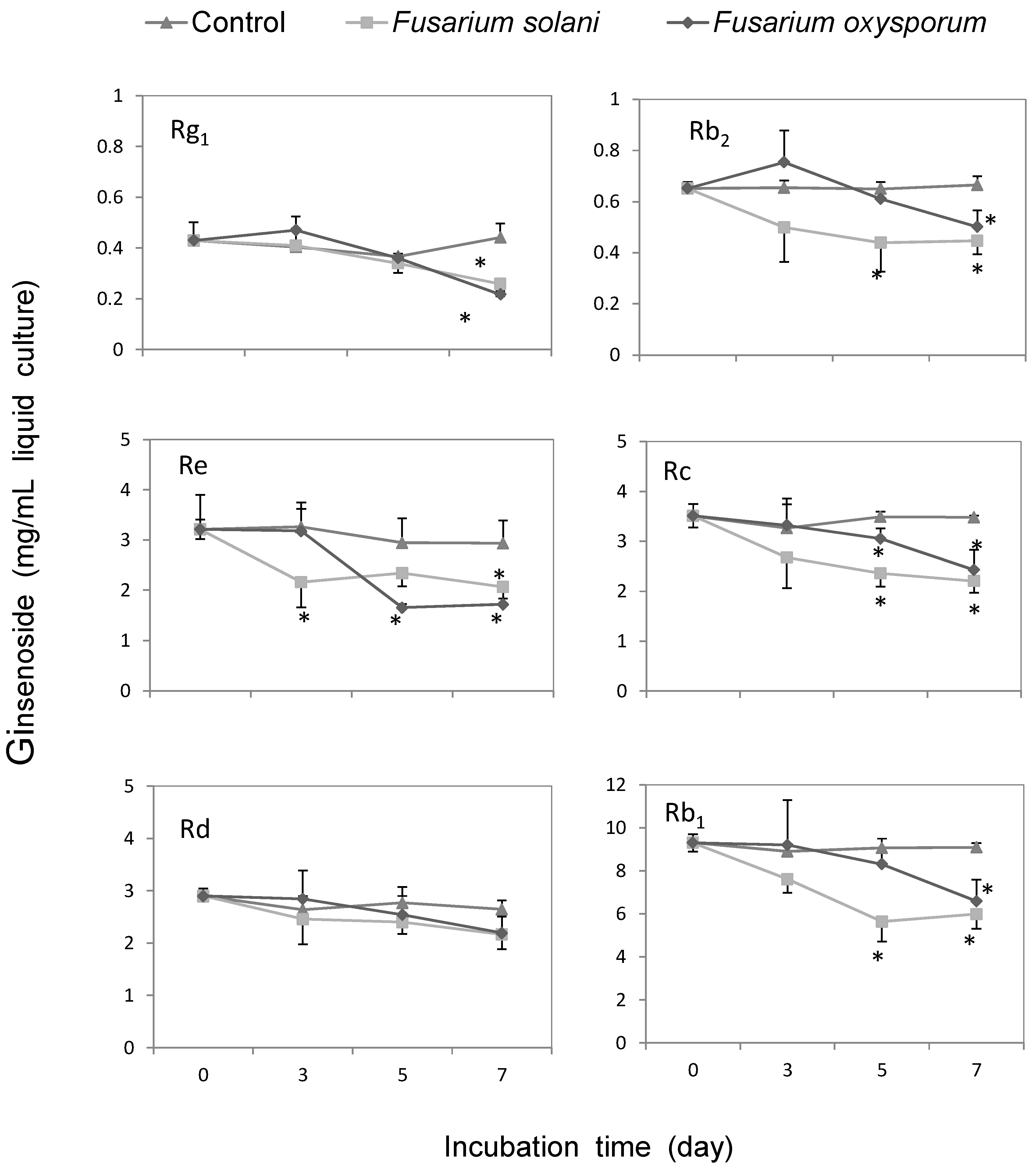
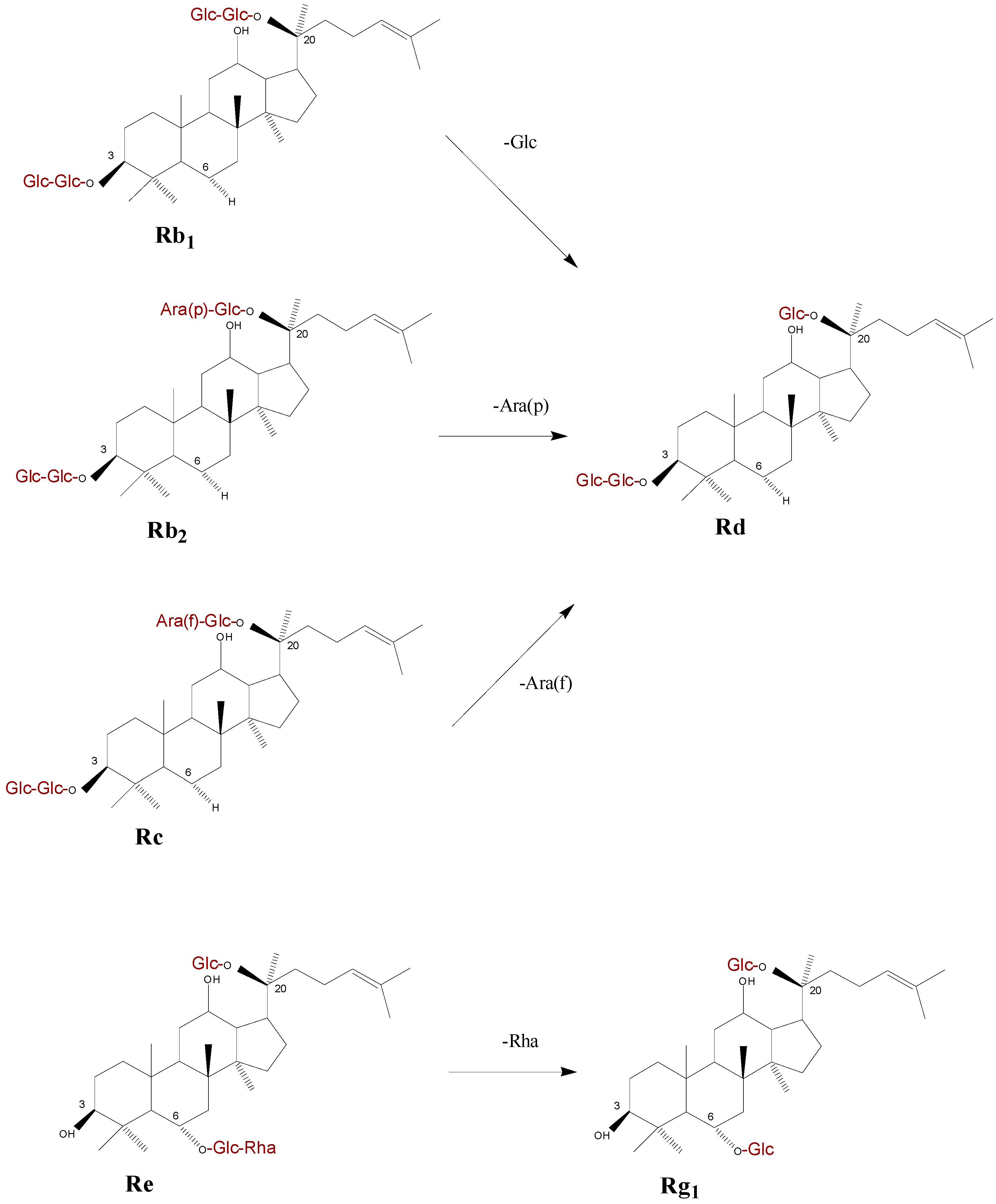
2.4. Discussion
3. Experimental Section
3.1. American Ginseng Roots and Fungal Inoculums
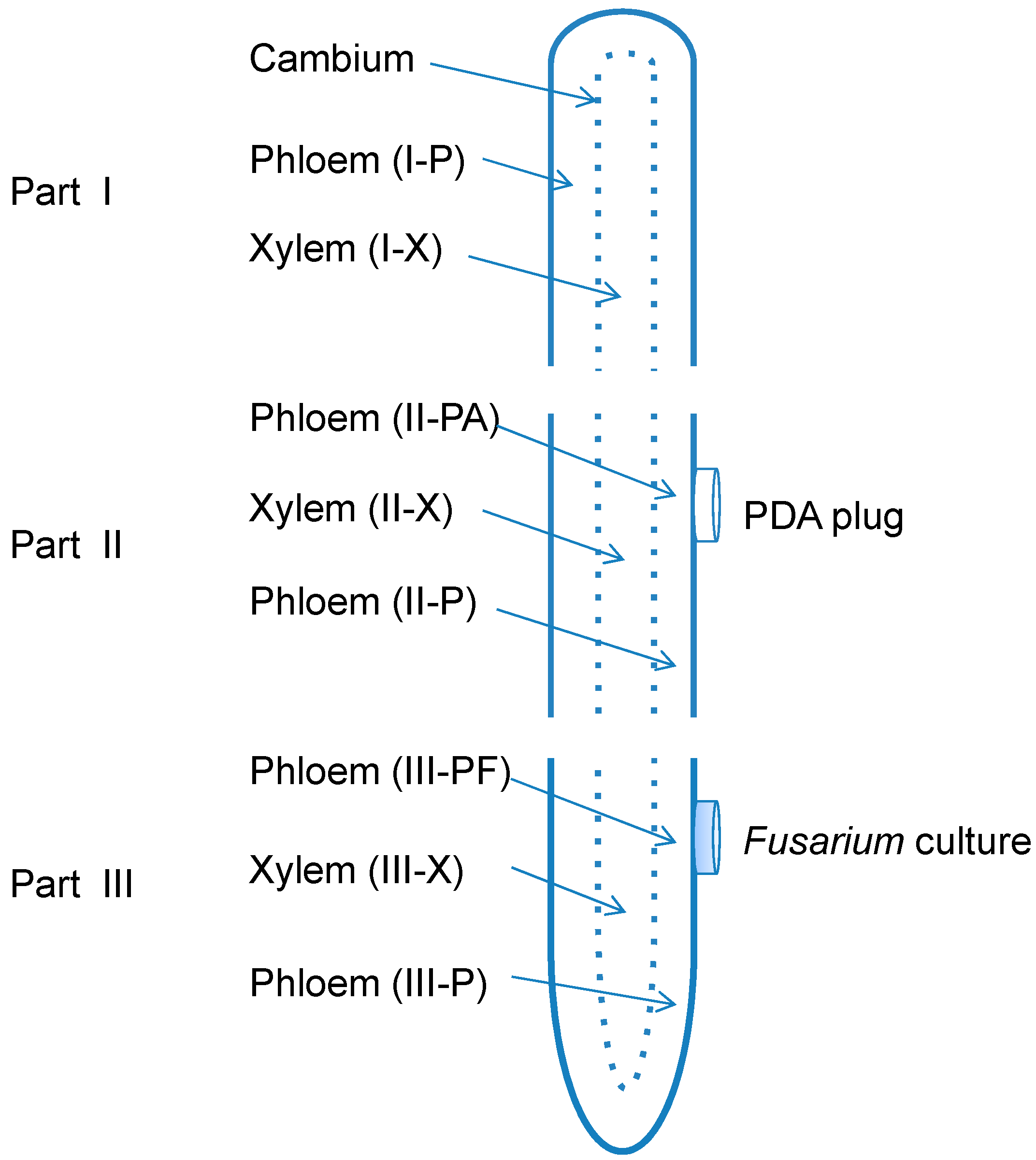
3.2. Root Treatments
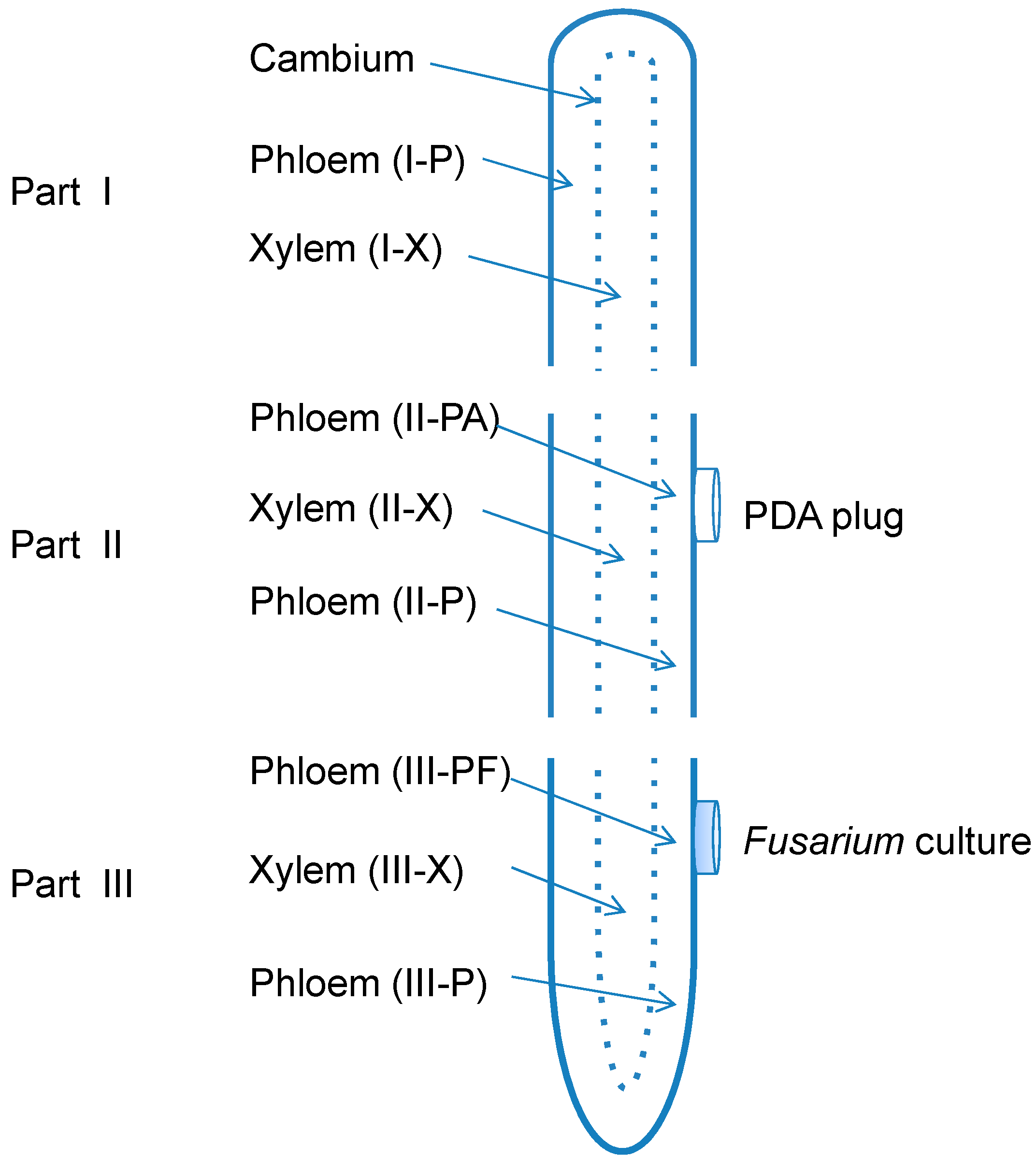
3.3. Molecular Detection of Fusarium spp. in American Ginseng Roots
3.4. Quantification of Ginsenoside from Root Extracts
3.5. Metabolic Effect of Fusarium spp. on the Concentrations of Ginsenoside
3.6. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, F.Y. The origin and classification of Chinese ginseng and American ginseng. In Chinese Ginseng and American Ginseng, 1st ed.; China Agricultural Scientech Press: Beijing, China, 2002; pp. 1–20. [Google Scholar]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Leung, K.W.; Wong, A.S. Pharmacology of ginsenosides: A literature review. Chin. Med. 2010, 5, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, J. Effects of ginsenoside Rb1 and Rg1 on synaptosomal free calcium level, ATPase and calmodulin in rat hippocampus. Chin. Med. J. 1995, 108, 544–547. [Google Scholar] [PubMed]
- Yokozawa, T.; Satoh, A.; Cho, E.J. Ginsenoside-Rd attenuates oxidative damage related to aging in senescence-accelerated mice. J. Pharm. Pharmacol. 2004, 56, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Li, D.M.; Wang, F.X.; Yan, F.Q.; Sun, H.P. In vitro anti-infection of ginseng saponin Rb2. Chin. J. Nosocomiol. 2011, 21, 1284–1286. [Google Scholar]
- Yuan, Z.Y.; Meng, Z.; Chai, Y.S.; Lan, J.Q.; Lei, F.; Li, H.Y.; Xing, D.M.; Li, H.Y.; Du, L.J. Ginsenoside Rg1 reduced the invasion of S. aureus into respiratory epithelial cells involving pro-inflammatory cytokines and glucocorticoid receptor. J. Chin. Pharm. Sci. 2011, 20, 266–274. [Google Scholar] [CrossRef]
- Chen, J.C.; Chen, L.D.; Tsauer, W.; Tsai, C.C.; Chen, B.C.; Chen, Y.J. Effects of ginsenoside Rb2 and Rc on inferior human sperm motility in vitro. Am. J. Chin. Med. 2001, 29, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Battinelli, L.; Mascellino, M.T.; Martino, M.C.; Lu, M.; Mazzanti, G. Antimicrobial activity of ginsenosides. Pham. Pharmacol. Commun. 1998, 4, 411–413. [Google Scholar]
- Bi, W.; Chen, J.; Jiao, X.L.; Gao, W.W. Identification of the pathogens causing the root rot and their pathogenicity on American ginseng in Beijing. Plant Prot. 2011, 37, 135–138. [Google Scholar]
- Punja, Z.K.; Wan, A.; Goswami, R.S.; Verma, N.; Rahman, M.; Barasubiye, T.; Seifert, K.A.; Lévesque, C.A. Diversity of Fusarium species associated with discolored ginseng roots in British Columbia. Can. J. Plant Pathol. 2007, 29, 340–353. [Google Scholar] [CrossRef]
- Rahman, M.; Punja, Z.K. Biochemistry of ginseng root tissues affected by rusty root symptoms. Plant Physiol. Biochem. 2005, 43, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Reeleder, R.D. Rhexocercosporidium panacis sp. nov., a new anamorphic species causing rusted root of ginseng (Panax quinquefolius). Mycologia 2007, 99, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Punja, Z.K. Fungal pathogens of American ginseng (Panax quinquefolium) in British Columbia. Can. J. Plant Pathol. 1997, 19, 301–306. [Google Scholar] [CrossRef]
- Reeleder, R.D.; Bramm all, R.A. Pathogenicity of Pythium species, Cylindrocarpon destructans and Rhizoctonia solani to ginseng seedling in Ontario. Can. J. Plant Pathol. 1994, 16, 311–316. [Google Scholar] [CrossRef]
- Darmono, T.W.; Owen, M.L.; Parke, J.L. Isolation and pathogenicity of Phytophthora cactorum from forest and ginseng garden soils in Wisconsin. Plant Dis. 1991, 75, 610–612. [Google Scholar] [CrossRef]
- Gao, W.W.; Jiao, X.L.; Bi, W.; He, C.N. The variation of ginsenosides in the infected roots of American ginseng (Panax quinquefolium). China J. Chin. Mat. Med. 2008, 33, 48–51. [Google Scholar]
- Turner, E.M. The nature of the resistance of oats to the take-all fungus. J. Exp. Bot. 1953, 11, 403–412. [Google Scholar] [CrossRef]
- Bernards, M.A.; Yousef, L.F.; Nicol, R.W. The allelopathic potential of ginsenosides. In Allelochemicals: Biological Control of Plant Pathogens and Diseases, 1st ed.; Inderjit, Mukerji, K.G., Eds.; Springer Press: Dordrecht, The Netherlands, 2006; pp. 157–175. [Google Scholar]
- Nicol, R.W.; Traquair, J.A.; Bernards, M.A. Ginsenosides as host resistance factors in American ginseng (Panax quinquefolius). Can. J. Bot. 2002, 80, 557–562. [Google Scholar] [CrossRef]
- Jiao, X.L.; Bi, W.; Li, M.; Luo, Y.; Gao, W.W. Dynamic response of ginsenosides in American ginseng. Plant Soil 2011, 339, 317–327. [Google Scholar] [CrossRef]
- Bais, P.H.; Prithiviraj, B.; Jha, K.A.; Ausubel, M.F.; Vivanco, M.J. Mediation of pathogen resistance by exudation of antimicrobials from roots. Nature 2005, 434, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J.; Ausubel, F.M. Isolation of phytoalexin-deficient mutants of Arabidopsis thaliana and characterization of their interactions with bacterial pathogens. PNAS 1994, 91, 8955–8959. [Google Scholar] [CrossRef] [PubMed]
- Toyomasu, T.; Kagahara, T.; Okada, K.; Koga, J.; Hasegawa, M.; Mitsuhashi, W.; Sassa, T.; Yamane, H. Diterpene phytoalexins are biosynthesized in and exuded from the roots of rice seedlings. Biosci. Biotechnol. Biochem. 2008, 72, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.S.; Gao, J.; Song, C.C.; Fang, Q.; Wang, N.; Zhao, T.J.; Liu, D.B.; Zhou, Y.F. Fungal sensitivity to and enzymatic deglycosylation of ginsenosides. Phytochemistry 2012, 78, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Punja, Z.K. Molecular and biochemical characterization of defense responses in ginseng (Panax quinquefolius) roots challenged with Fusarium equiseti. Physiol. Mol. Plant Pathol. 2008, 72, 10–20. [Google Scholar] [CrossRef]
- Yousef, L.F.; Bernards, M.A. In vitro metabolism of ginsenosides by the ginseng root pathogen Pythium irregulare. Phytochemistry 2006, 67, 1740–1749. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.S.; Wang, J.; Li, J.; Fu, L.; Gao, J.; Du, X.L.; Bi, H.T.; Zhou, Y.F.; Tai, G.H. Highly selective biotransformation of ginsenoside Rb1 to Rd by the phytopathogenic fungus Cladosporium fulvum (syn. Fulvia fulva). J. Ind. Microbiol. Biotechnol. 2009, 36, 721–726. [Google Scholar] [CrossRef]
- Fäldt, J.; Solheim, H.; Långström, B.; Borg-karlson, A.K. Influence of fungal infection and wounding on contents and enantiomeric compositions of monoterpenes in phloem of Pinus sylvestris. J. Chem. Ecol. 2006, 32, 1779–1795. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef]
- van Burik, J.A.; Schreckhise, R.W.; White, T.C.; Bowden, R.A.; Myerson, D. Comparison of six extraction techniques for isolation of DNA from filamentous fungi. Med. Mycol. 1998, 36, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Mirete, S.; Vazquez, C.; Mule, G.; Jurado, M.; Gonzalez-jaen, M.T. Differentiation of Fusarium verticillioides from banana fruits by IGS and EF-1α sequence analyses. Eur. J. Plant Pathol. 2004, 110, 515–523. [Google Scholar] [CrossRef]
- Li, T.S.C.; Mazza, G.; Cottrell, A.C.; Gao, L. Ginsenosides in roots and leaves of American ginseng. J. Agric. Food Chem. 1996, 44, 717–720. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds ginsenoside Rb1, Rb2, Rc, Rd, Re, and Rg1 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, X.; Lu, X.; Chen, A.J.; Luo, Y.; Hao, J.J.; Gao, W. Effects of Fusarium solani and F. oxysporum Infection on the Metabolism of Ginsenosides in American Ginseng Roots. Molecules 2015, 20, 10535-10552. https://doi.org/10.3390/molecules200610535
Jiao X, Lu X, Chen AJ, Luo Y, Hao JJ, Gao W. Effects of Fusarium solani and F. oxysporum Infection on the Metabolism of Ginsenosides in American Ginseng Roots. Molecules. 2015; 20(6):10535-10552. https://doi.org/10.3390/molecules200610535
Chicago/Turabian StyleJiao, Xiaolin, Xiaohong Lu, Amanda Juan Chen, Yi Luo, Jianjun J. Hao, and Weiwei Gao. 2015. "Effects of Fusarium solani and F. oxysporum Infection on the Metabolism of Ginsenosides in American Ginseng Roots" Molecules 20, no. 6: 10535-10552. https://doi.org/10.3390/molecules200610535
APA StyleJiao, X., Lu, X., Chen, A. J., Luo, Y., Hao, J. J., & Gao, W. (2015). Effects of Fusarium solani and F. oxysporum Infection on the Metabolism of Ginsenosides in American Ginseng Roots. Molecules, 20(6), 10535-10552. https://doi.org/10.3390/molecules200610535