1. Introduction
Blueberry, also known as huckleberry or mustikka, belongs to the
Ericaceae Vaccinium group of deciduous shrub plants and is popular because of its high health care value, as it is recommended by the FAO as one of the five major healthy fruits [
1,
2]. At present, the biological activity of blueberry extract is widely studied [
3,
4,
5,
6,
7]. However, no reports are available describing the effect of soluble polysaccharides, derived from blueberries, on physiological activity.
Recently, many studies have shown that polysaccharides possess antiviral activity, anti-aging properties, resistance to bone marrow suppression, anti-tumor, and immunoregulatory activities [
8,
9,
10] with limited side effects or adverse reactions [
11,
12], thus making research on polysaccharide immunomodulatory effects popular [
13,
14,
15].
A growing amount of research has shown that polysaccharides possess antitumor activity and in some cases these antitumor effects may be mediated indirectly, possibly through the innate and/or adaptive immune system. Should this be the case there are a number of immune effector functions that polysaccharides have been shown to promote and therefore may be implicated in contributing to the observed antitumor activity, and these include; the enhancement of phagocytosis of macrophage [
16], the activation of T and B lymphocytes [
11], the activation of natural killers cells (NK cells) and lymphokine activated killer (LAK) cells [
17]. In addition, polysaccharides (or peptide-polysaccharides) have been shown to promote the secretion of a range of important immunoregulatory cytokines [
18], such as interleukins [
19], tumor necrosis factor-α (TNF-α) [
20] and interferon-γ (IFN-γ) [
16,
21]. Hence, finding a new polysaccharide from natural source with anticancer and immunopotentiation activities could be a new tool for cancer therapy [
22]. This study aimed to ascertain the anti-tumor activities of a polysaccharide purified from blueberries (BBP3-1) using a sarcoma-180 tumor-bearing mouse model and the possible immune mechanism is discussed.
3. Experimental Section
3.1. Materials
S180 tumor cells were kept in RPMI-1640 with 10% fetal calf serum (FCS) at 37 °C in a 5% CO2 atmosphere to sustain its bio-activity.
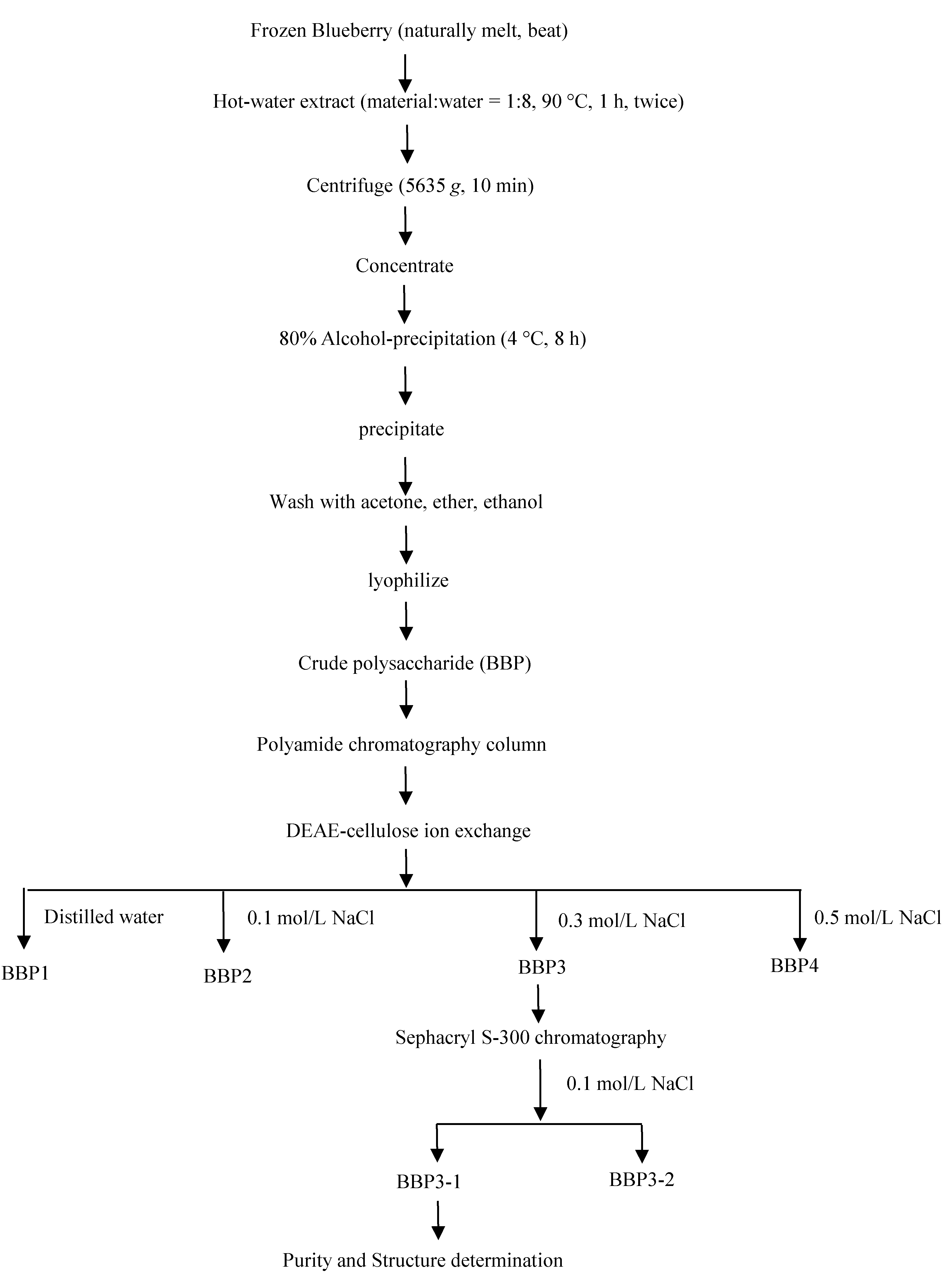
The extraction flow diagram of BBP3-1 was presented in
Figure 1. A detailed analysis Purity Determination and Composition property of BBP3-1 were previously presented [
23].
3.2. Reagents
RPMI-1640, DMEM, and DMEM/high glucose (including ketonic acid sodium), penicillin, streptomycin (HYCLONE, Logan, UT, USA), glutamine (AMRESCO, Solon, Tucson, AZ, USA), HEPES, MTT, Trypan Blue (Sigma Co., St. Louis, MO, USA), fetal bovine serum (Hang Zhou Sijiqing Reagent Co., Hangzhou, China), cyclophosphamide (Cy) (Jiang Su Heng Rui Pharmaceutical Co. Ltd., Lianyugang, China), interferon IFN-γ, tumor necrosis factor TNF-α, and interleukin IL-2 ELISA Kit (RND Co., Minneapolis, MN, USA), S180 tumor cells, NK target cell (YAC-1 mice lymphoma cell) (CAS cell bank, Shanghai, China).
Figure 1.
The extract flow diagram of BBP3-1.
Figure 1.
The extract flow diagram of BBP3-1.
3.3. Instruments and Equipment
CO2 incubator (Thermo Scientific 8000, Vernon Hills, IL, USA), Microplate Reader (BIO-RAD 680, Hercules, CA, USA), inverted microscope (Olympus-CK2, Tokyo, Japan) Precision Electronic Balance (TP-313, Sartorius, Denver, CO, USA), low-speed centrifuge, (SC-2542, Zhengzhou Boke Instrument Equipment Co., Ltd., Zhengzhou, China). 96-well flat culture plate, 12-well flat culture plate, 96-well U-culture plate (Costar, Washington, DC, USA).
3.4. Animals
Kun Ming mice (female), aged 4–5 weeks, body weight 20 ± 2 g, were purchased from Chinese Medial Sciences University. The animals were provided with free access to standard rodent chow and water, and were kept at a temperature of 23 ± 2 °C in a 12-h light-dark cycle. The animals were humanely cared for, according to the Standards for Laboratory Animals of China (GB 14923-94, GB 14922-94, and GB/T 14925-94).
3.5. Tumor-Bearing Mice Modeling Building
Mice were randomly divided into six groups, including a normal control group, model group, positive control group, BBP3-1 high-dose group, BBP3-1 medium-dose group, and BBP3-1 low-dose group, each group consisting of 10 mice.
S180 tumor cells were diluted to 5 × l06/mL in saline and injected into the mouse left axillary subcutaneous tissue under aseptic conditions, with each mouse receiving 0.2 mL.
After 24 h, the mice from the BBP3-1 dose groups were orally administered with the corresponding dose (respectively, 400 mg/kg, 200 mg/kg, and 100 mg/kg body mass), gavage volume for each mouse is 0.2 mL; the positive control group was administered with cyclophosphamide (20 mg/kg body mass) orally; and the normal control and model groups were administered with distilled water of the same gavage volume (0.2 mL) for 15 days.
3.6. Purified Mice Peritoneal Macrophages Cells (PMФ)
24 h after the last drug administration, the mice were sacrificed by cervical dislocation, and blood was taken from the eye and centrifuged (1368 g for 10 min) to obtain serum. The dead mice were then disinfected in ethanol for 5 min.
Under aseptic conditions, the peritoneum was exposed, injected intraperitoneally with RPMI-1640 culture solution without serum, and irrigated adequately (to prevent the viscera from breaking). This was repeated 3–4 times, after which the suspension of peritoneal macrophages (PMФ) was collected, centrifuged (152 g for 5 min) and re-suspended in RPMI-1640 complete culture solution. The fraction of living cells was determined to be >95% by Trypan Blue staining, and the concentration of PMФ cells was adjusted to 5 × 106/mL.
The PMФ cells were cultured in 96-well flat cell culture plates (100 µL per well) in a humidified 5% CO2 atmosphere at 37 °C for 2 h. The wells were then washed with RPMI-1640 culture solution, and 100 µL of RPMI-1640 was added. The resulting cells were purified mice PMФ cells.
3.7. Index Detection Method
3.7.1. Anti-Tumor Rate, Spleen Index, and Thymus Index
The tumor, spleen, and thymus were removed from the mice. The anti-tumor rate, spleen index, and thymus index were calculated as follows:
where,
wc is the tumor weight of the model control and
wd is the tumor weight of the drug-treated animal.
3.7.2. Determination of Peritoneal Macrophages Phagocytosis Function
For the neutral red phagocytosis experimental method, purified PMФ cells were centrifuged at 152 g for 10 min after being cultured for 24 h. The suspended cells were then seeded in a 96-well flat cell culture plate (100 µL per well). Added 100 µL of neutral red (1 g/L saline solution) to each well and placed the culture plate inside a 5% CO
2 incubator for cultivation at 37 °C for 30 min. After that, centrifugation was applied at 152 g for 10 min and the supernatant discarded. The plate was washed with PBS (200 µL per well) three times before adding the lysis agent (100 µL per well) and incubating overnight. The lysis agent was concocted with acetic acid: anhydrous ethanol 1:1 (v/v). 12 h later, an ELISA reader was used to measure the OD value at 540 nm and the strength of PMФ phagocytosis was indicated by the OD
540 value [
33].
3.7.3. Lymphocyte Proliferation Assay
Preparation of cell suspension of spleen lymphocytes from mice:
The aseptically treated spleen of the mice was weighed in RPMI-1640 medium without serum and then rinsed twice. Approximately 5 mL of serum-free RPMI-1640 medium was drawn into a 5 mL syringe, which was then injected into the spleen. The cells in the spleen were then gently washed out. This operation was repeated several times until the outer membrane of the spleen was transparent. The remaining spleen was ground using a syringe and forced through a 100-mesh sieve. All the spleen cell was collected into RPMI-1640 medium and then the cell suspension was prepared.
The cell suspension was then centrifuged at 152 g for 5 min, and the supernatant was discarded. Approximately 3 mL of tris-NH4Cl was added to remove red blood cells, followed by centrifugation at 152 g for 5 min. This operation was repeated twice. The cell pellet was then resuspended with serum-free RPMI-1640 medium and centrifuged at 152 g for 5 min.
Resulting cell pellet was then resuspended in complete RPMI-1640 medium and Trypan Blue staining was used to count living cells to confirm >95% viability, and the cell density was adjusted to 5 × 106/mL.
3.7.4. Detection of Spleen Lymphocyte Activity (MTT)
Spleen lymphocytes were added to 96-well cell flat culture plates (200 µL per well). The blank control group (RPMI-1640 complete culture fluid) was set up at the same time, and there were four wells for each group.
The 96-well cell culture plate was cultured for 44 h in a 5% CO
2 incubator at 37 °C, and then 20 µL of MTT solution was added to each well and cultured for an additional 4 h. The supernatant was discarded after centrifuging at 152 g for 10 min, and 150 µL DMSO was then added to each well. The plate was shaken for 10 min until the purple crystals completely dissolved, and an ELISA reader was used to measure the OD value at 570 nm. The lymphocyte proliferation and conversion rate were reflected by the OD
570 values [
34].
3.7.5. Determination of NK Cell Activity (LDH)
The target cells (Yac-1) were washed with RPMI-1640 medium, viability >95% was confirmed by Trypan Blue staining, and then the cell concentration was adjusted to 1 × 105/mL. Suspended spleen lymphocytes were used as effector cells (E) suspension, and their concentration was adjusted to 5 × 106/mL.
E and T cells (0.1 mL; E/T = 50:1) were added separately into a cell culture plate with U-bottom wells. Each specimen was allocated to four wells. Target cells were included in a natural release well [(Yac-1) and RPMI1640 medium, 0.1 mL each] and a maximum release well [(Yac-1) and 2.5% Triton X, 0.1 mL each], centrifuged at 152 g for 2 min, then incubated in a 5% CO
2 incubator at 37 °C. After 2 h, the 0.1 mL supernatant from each well was transferred to another culture plate that had been pre-warmed (37 °C), and 0.1 mL of recently prepared LDH substrate solution was added. The reaction was allowed to proceed for 15 min. To stop the enzymatic reaction, 30 µL of 1 mol/L citric acid stop buffer was added. An ELISA reader was used to read the OD
490 value of each well at 490 nm to calculate the viability of NK cells [
35].
3.7.6. Detection of IFN-γ, TNF-α, and IL-2
The serum samples and medium supernatants from spleen lymphocytes cultured for 48 h were applied to 3 wells each. The content of IFN-γ, TNF-α, and interleukin IL-2 were examined by ELISA kit. Summary procedures: Preparing reagents, samples and standard → Add prepared sample and standard, incubated 30 min at 37 °C → Plate washed five times, adding HRP-Conjugate Reagent incubated 30 min at 37 °C → Plate washed five times, adding Chromogen Solution A and B incubated 30 min at 37 °C → Add stop solution → Measure within 15 min → Calculation.
3.8. Statistical Analysis
All experiments were repeated at least three times. Analysis of variance and t-test were used. Data are reported as the mean ± SD (standard deviation) and average. Statistical calculations were performed using SPSS software (version 12.0, SPSS Inc., Chicago, IL, USA).
4. Conclusions
This study, to our knowledge, is the first report on anti-cancer and immune adjustment of a polysaccharide (BBP3-1) isolated from blueberry. It is found that the BBP3-1 has the strongly inhibited the growth of S180 tumor and the best dose is 100 mg/kg body mass in this experiment. In addition, the BBP3-1 can up-regulate the immunity of S180 tumor-bearing mice by increasing the thymus index and spleen index, enhancing lymphocyte conversion and multiplication capacity, promoting macrophage phagocytosis, promoting immune factor secretion (TNF-α, IFN-γ, and IL-2), and enhancing NK cell activity. However, elucidation of the detailed mechanism requires further experiments.



{kind=link}