3-(Methoxycarbonylmethylene)isobenzofuran-1-imines as a New Class of Potential Herbicides


 ,
, 
 ,
, 

Abstract
:
1. Introduction
2. Results and Discussion
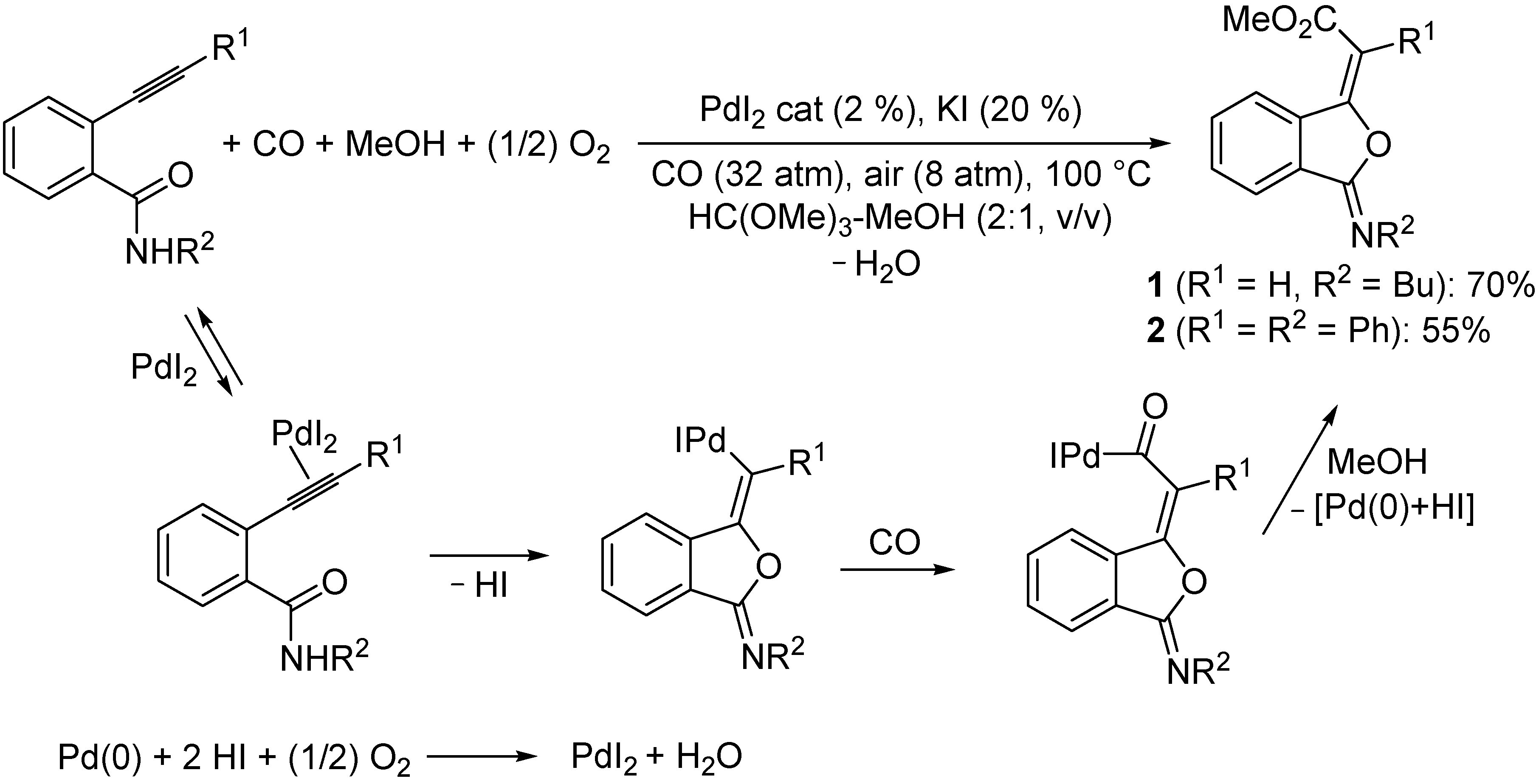
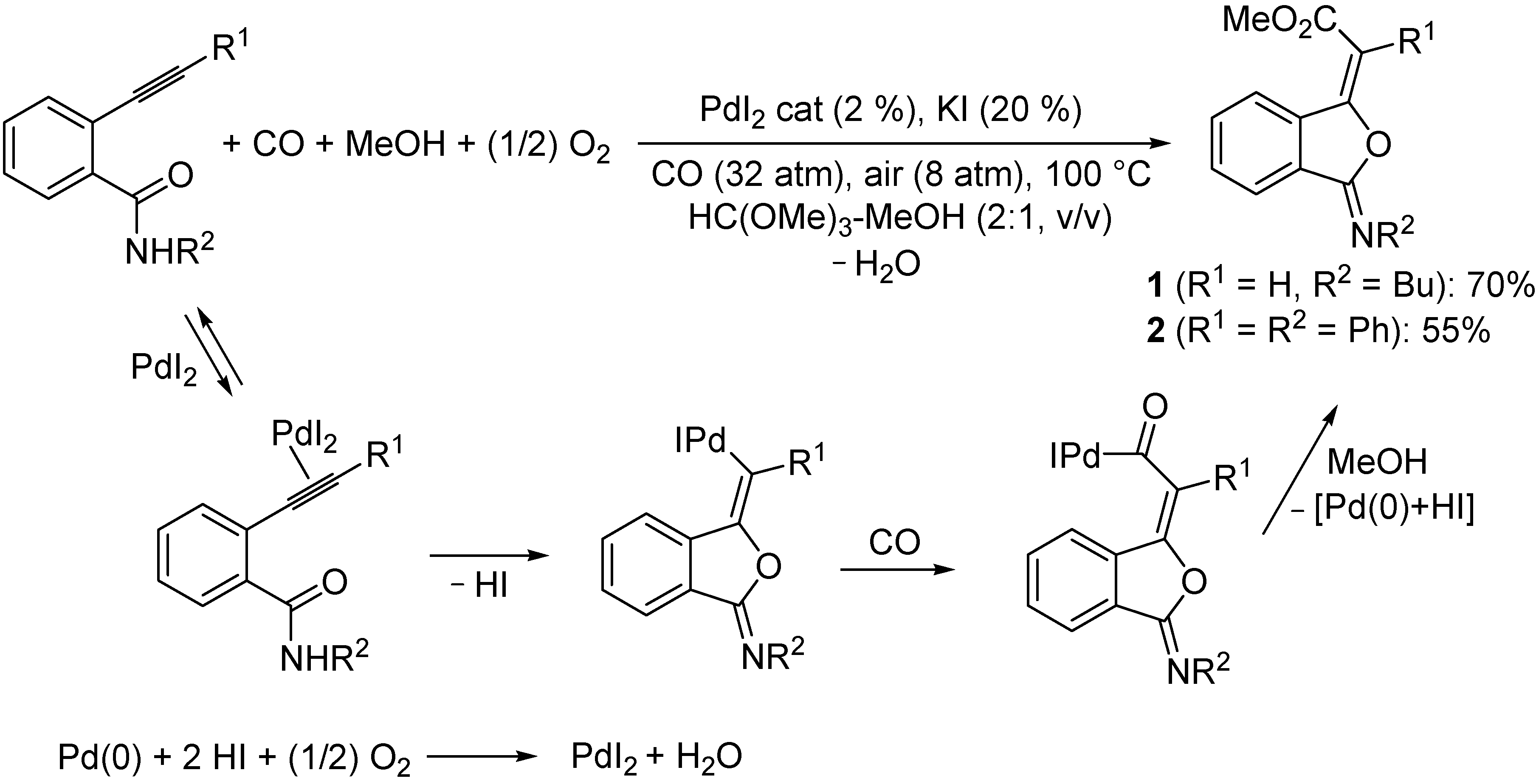
2.1. Synthesis of 3-(Methoxycarbonylmethylene)isobenzofuran-1-imines 1 and 2
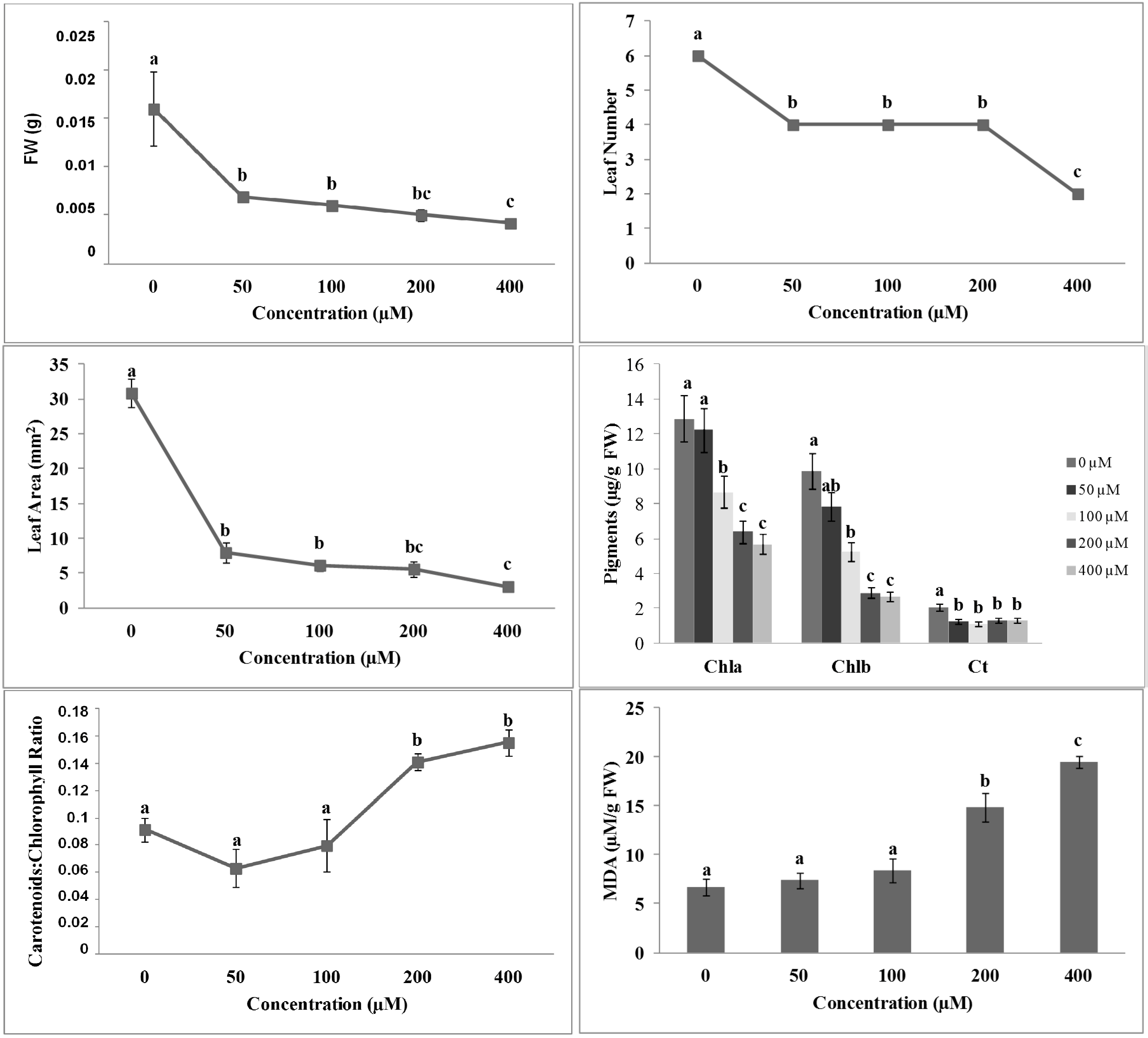
2.2. Effects of 3-(Methoxycarbonylmethylene)isobenzofuran-1-imines 1 and 2 on A. thaliana Shoot Morpho-Physiological Parameters
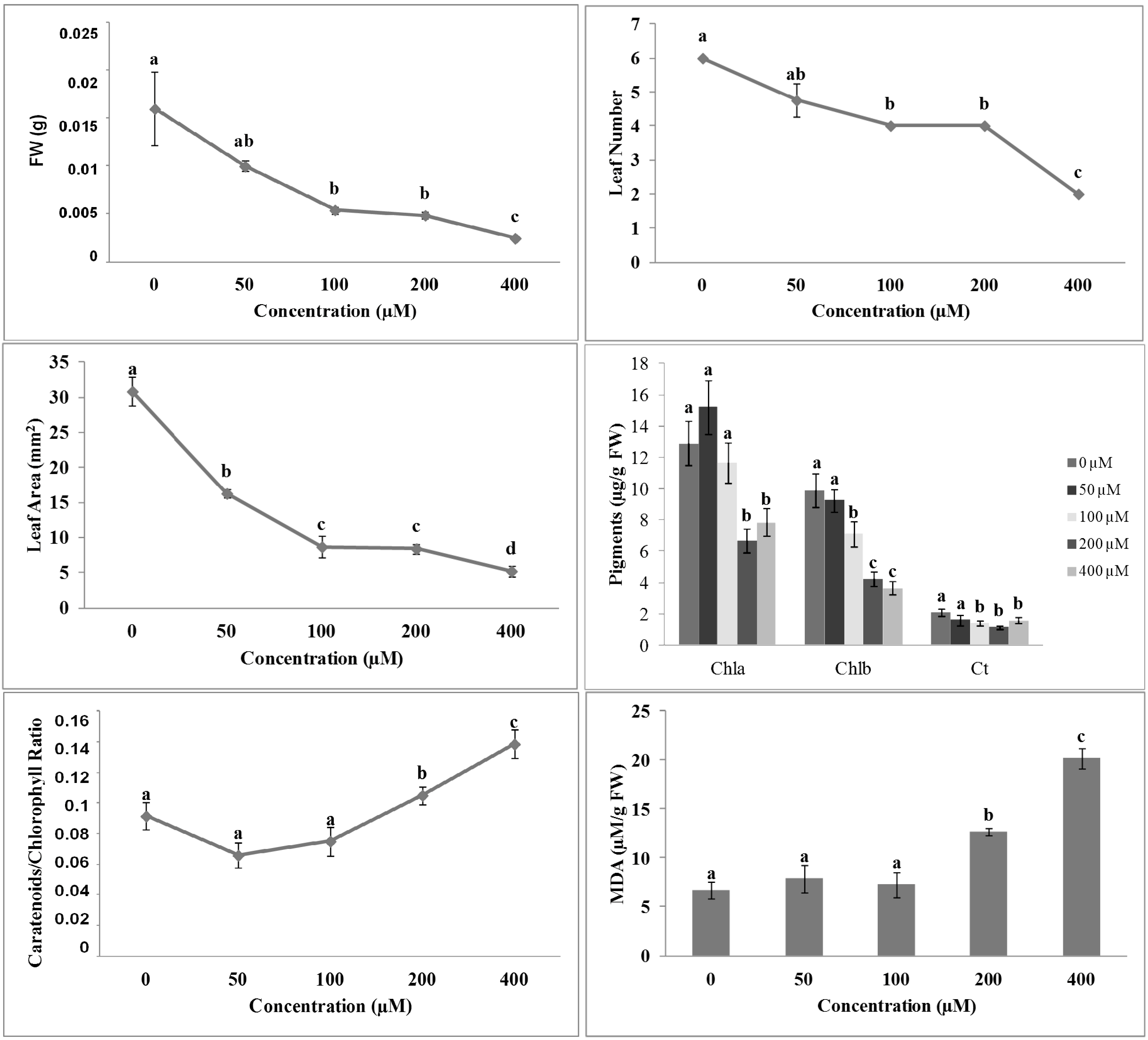
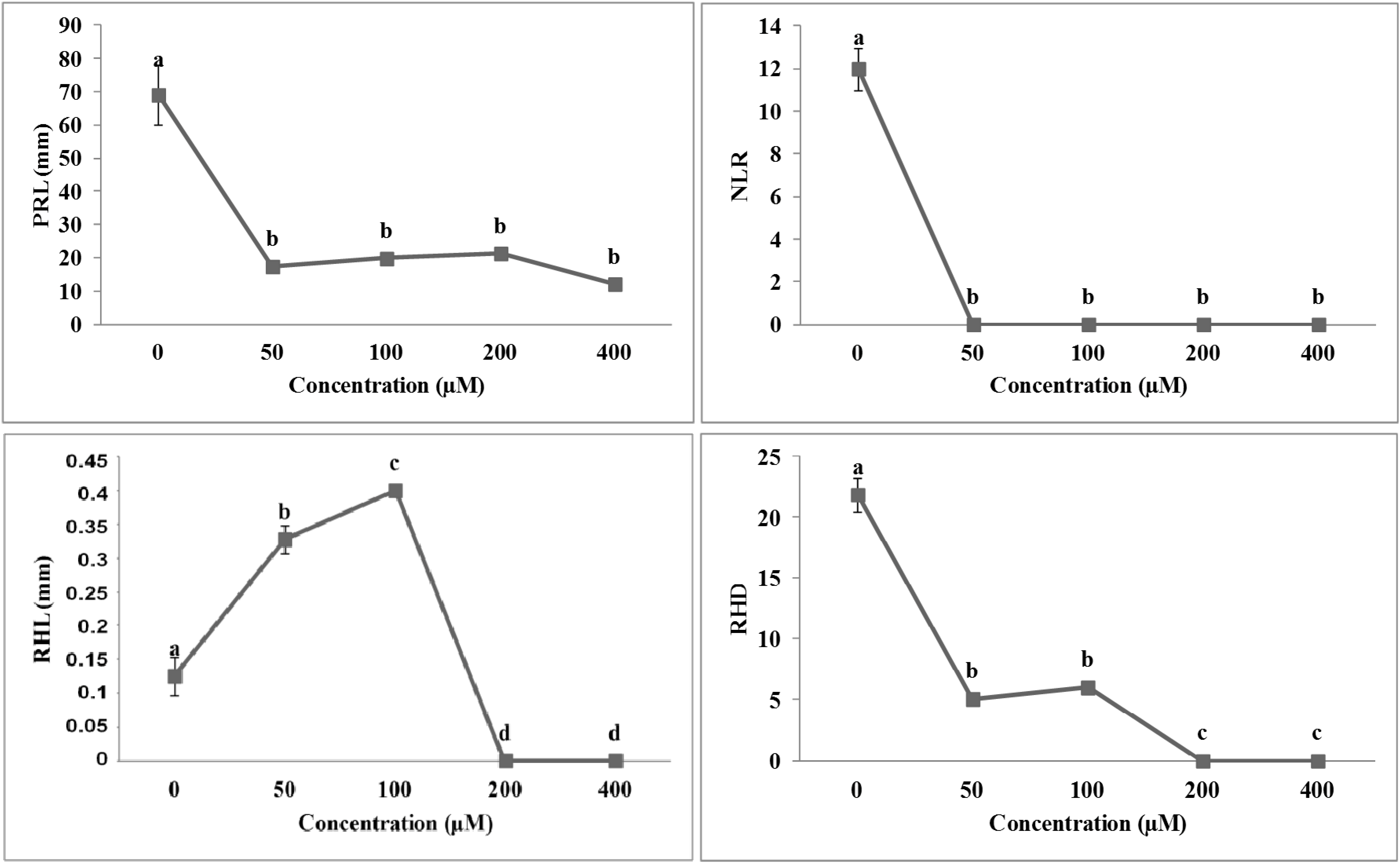
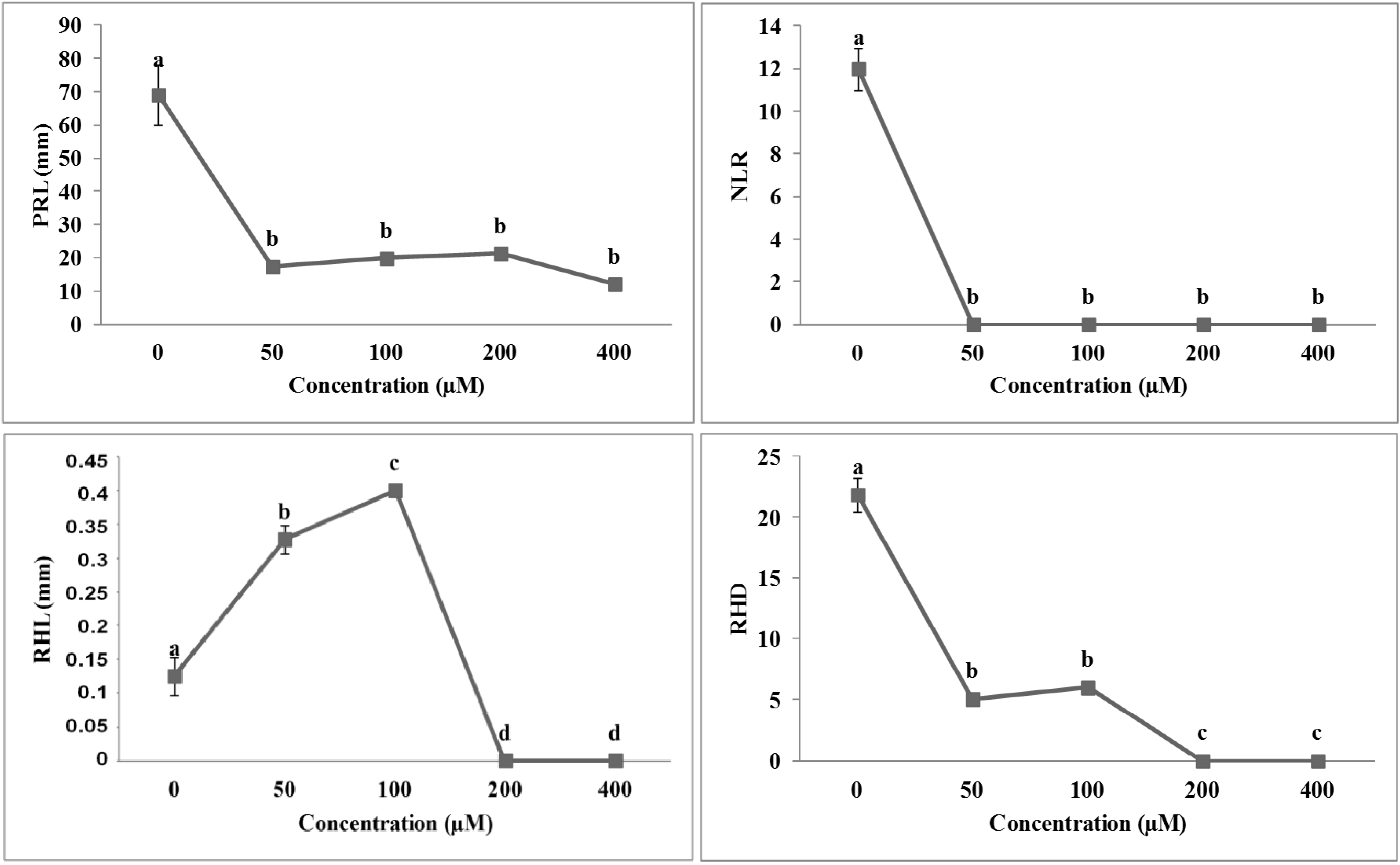
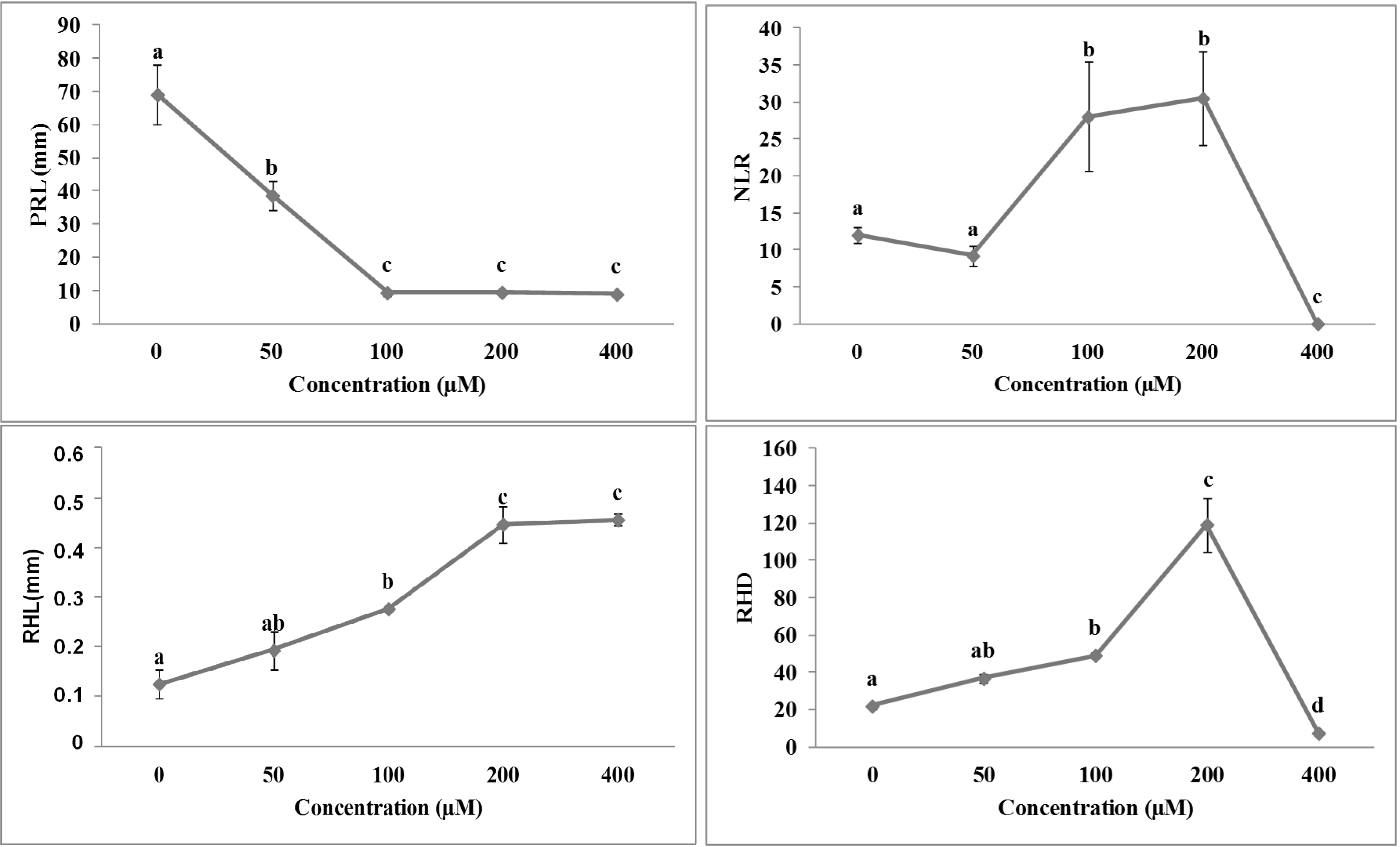
2.3. Effects of Isobenzofuranimines 1 and 2 on Root Morphology of A. thaliana


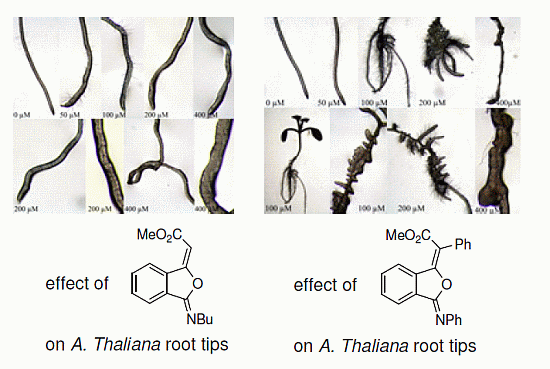{kind=link}
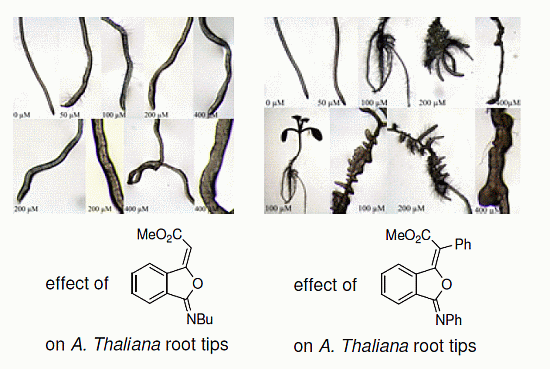{kind=link}
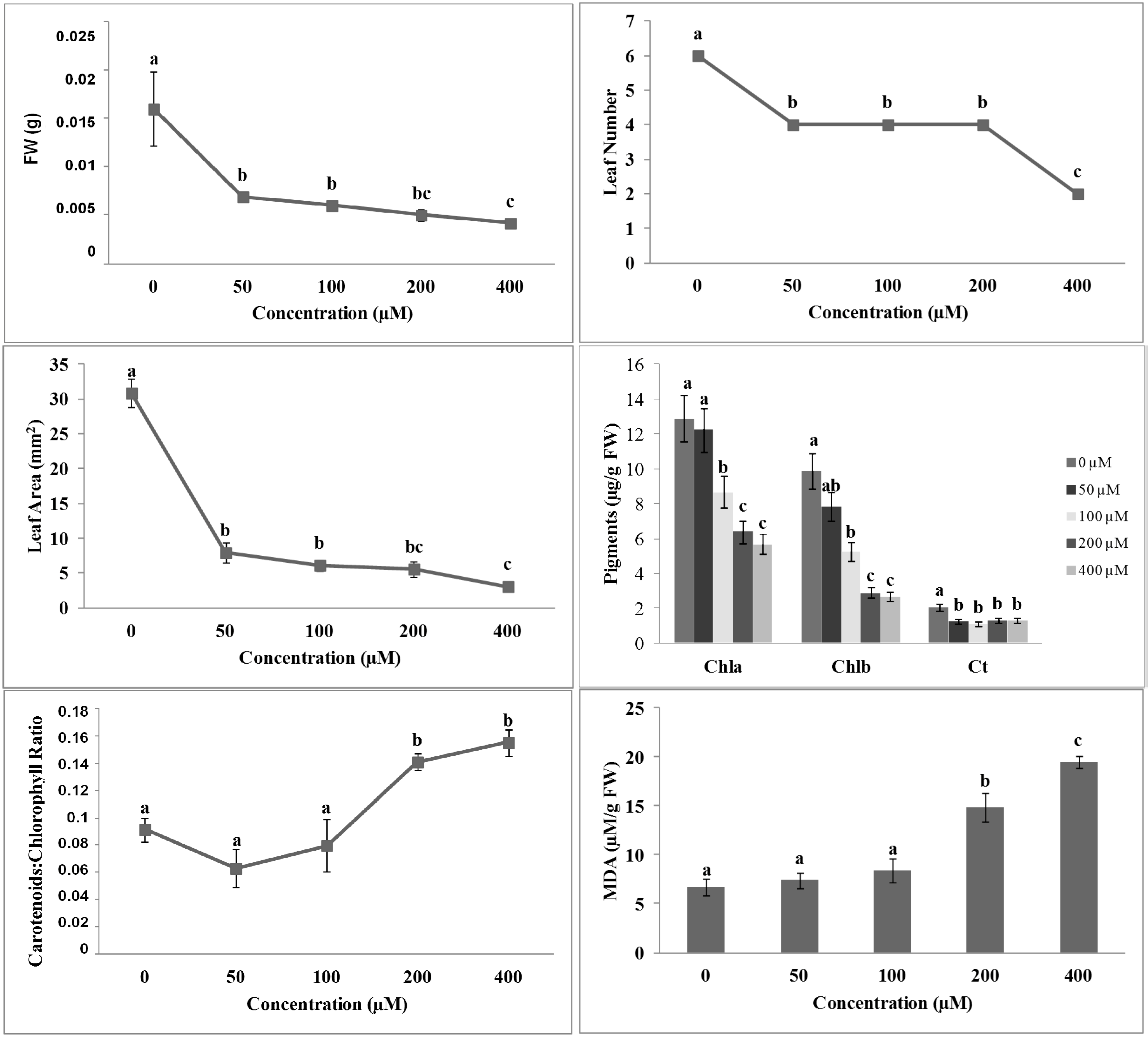{kind=link}
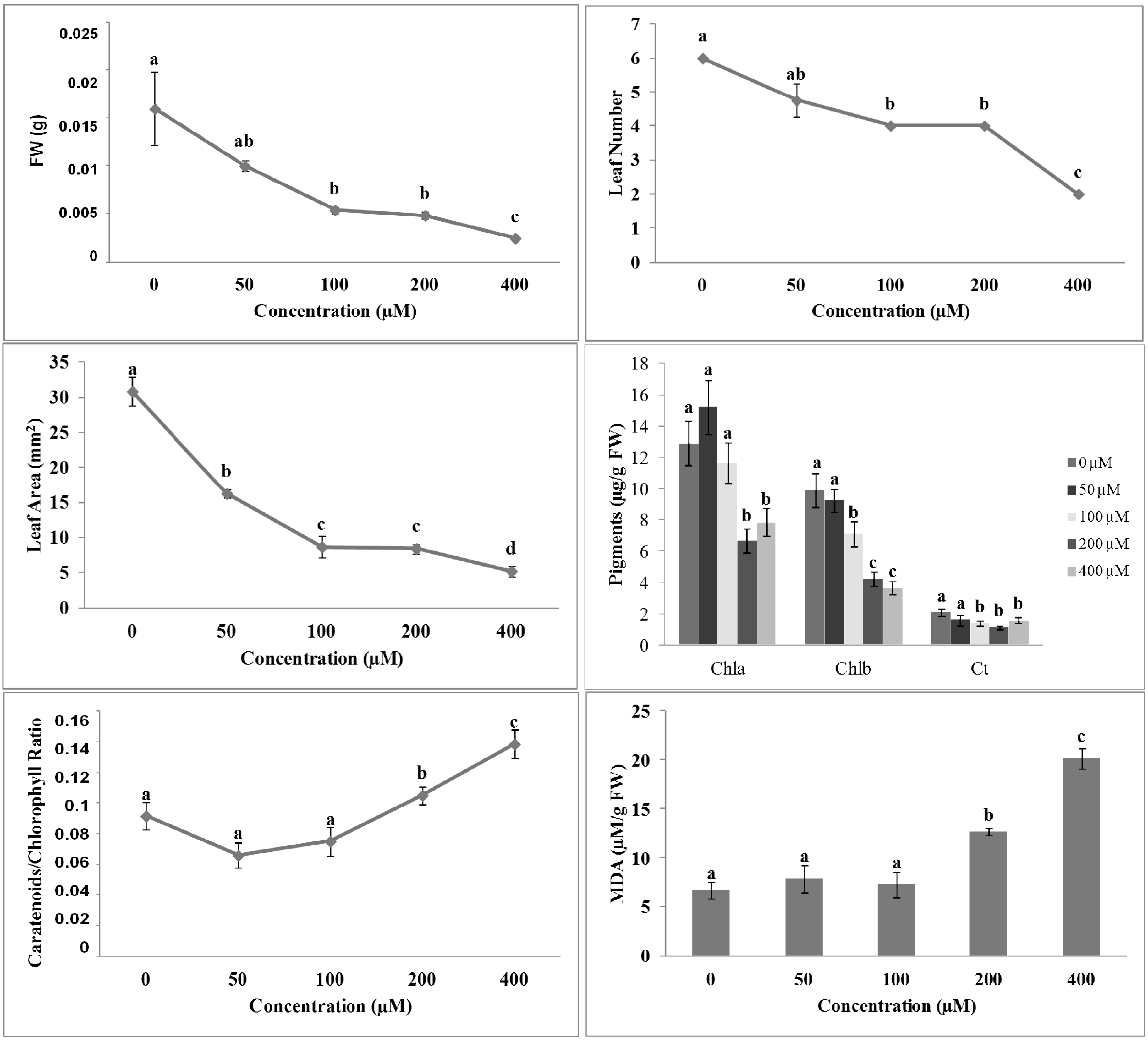{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ED50 (µM) | ||||
|---|---|---|---|---|
| Isobenzofuranimine | PRL | NLR | RHD | RHL |
| 1 | 28.5 (±0.46) a | 9.73 * e-13 (±0.0001) a | 20.46 (±1.5) a | 119.87 (±11.78) a |
| 2 | 52.5 (±4.8) b | 225.06 (±5.29) b | 397.1 (±0.78) b | ND |
| Stimulation (µM) | ||||
| 1 | ND | ND | ND | 18.16 (±0.21) a |
| 2 | ND | 28.66 (±5.3) | 60.97 (±0.04) | 108.2 (±2.1) b |


3. Experimental
3.1. General Information
3.2. Preparation of Isobenzofuranimines 1 and 2 [29]
3.3. Seedlings Growth Bioassay
3.4. Measurement of Photosynthetic Pigments
3.5. Lipid Peroxidation
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Böger, P.; Wakabayashi, K.; Hirai, K. (Eds.) Herbicide Classes in Development: Mode of Action, Targets, Genetic Engineering, Chemistry; Springer: Berlin, Germany, 2002.
- Heap, I. International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 21 September 2005).
- Ryan, G.F. Resistance of common groundsel to simazine and atrazine. Weed Sci. 1970, 18, 614–616. [Google Scholar]
- Holt, J.S.; LeBaron, H.M. Significance and distribution of herbicide resistance. Weed Technol. 1990, 4, 141–149. [Google Scholar]
- Beckie, H.J. Herbicide-resistant weeds: Management tactics and practices. Weed Technol. 2006, 20, 793–814. [Google Scholar] [CrossRef]
- Green, J.M. Current State of Herbicides in Herbicide-Resistant Crops. Pest Manag. Sci. 2014. [Google Scholar] [CrossRef]
- Gabriele, B.; Mancuso, R.; Salerno, G. Oxidative carbonylation as a powerful tool for the direct synthesis of carbonylated heterocycles. Eur. J. Org. Chem. 2012, 2012, 6825–6839. [Google Scholar] [CrossRef]
- Gabriele, B.; Salerno, G.; Costa, M. Oxidative Carbonylations. Top. Organomet. Chem. 2006, 18, 239–272. [Google Scholar]
- Gabriele, B.; Costa, M.; Salerno, G.; Chiusoli, G.P. An efficient and selective palladium-catalysed oxidative dicarbonylation of alkynes to alkyl- or aryl-maleic esters. J. Chem. Soc. Perkin Trans. 1 1994, 1, 83–87. [Google Scholar]
- Veluri, R.; Weir, T.L.; Bais, H.P.; Stermitz, F.R.; Vivanco, J.M. Phytotoxic and antimicrobial activities of catechin derivatives. J. Agric. Food Chem. 2004, 52, 1077–1082. [Google Scholar] [CrossRef]
- Baziramakenga, R.; Simard, R.R.; Leroux, G.D. Effects of benzoic and cinnamic acids on growth, mineral composition, and chlorophyll content of soybean. J. Chem. Ecol. 1994, 20, 2821–2833. [Google Scholar] [CrossRef]
- Einhellig, F.A.; Reigosa, M.J.; Pedrol, N. The physiology of allelochemical action: Clues and views. In Allelopathy: From Molecules to Ecosystems; Reigosa, M.J., Pedrol, N., Eds.; Science Publishers: Enfield, NH, USA, 2002; pp. 1–23. [Google Scholar]
- Graña, E.; Sotelo, T.; Díaz-Tielas, C.; Reigosa, M.J.; Sánchez-Moreiras, A.M. The phytotoxic potential of the terpenoid citral on seedlings and adult plants. Weed Sci. 2013, 61, 469–481. [Google Scholar] [CrossRef]
- Djordjevic, D.; Cercaci, L.; Alamed, J.; McClements, D.J.; Decker, E.A. Stability of citral in protein-and gum arabic-stabilized oil-in-water emulsions. Food Chem. 2008, 106, 698–705. [Google Scholar] [CrossRef]
- Filella, I.; Serrano, L.; Serra, J.; Penuelas, J. Evaluating wheat nitrogen status with canopy reflectance indices and discriminant analysis. Crop Sci. 1995, 35, 1400–1405. [Google Scholar] [CrossRef]
- Sánchez-Moreiras, A.M.; Martínez-Peñalver, A.; Reigosa, M.J. Early senescence induced by 2–3H-benzoxazolinone (BOA) in Arabidopsis thaliana. J. Plant Physiol. 2011, 168, 863–870. [Google Scholar] [CrossRef]
- Liu, J.; Wu, Y.H.; Yang, J.J.; Liu, Y.D.; Shen, F.F. Protein degradation and nitrogen remobilization during leaf senescence. J. Plant Biol. 2008, 51, 11–19. [Google Scholar] [CrossRef]
- Smith, R.D.; Wilson, J.E.; Walker, J.C.; Baskin, T.I. Protein-phosphatase inhibitors block root hair growth and alter cortical cell shape of Arabidopsis roots. Planta 1994, 194, 516–524. [Google Scholar] [CrossRef]
- Ishihara, H.; Martin, B.L.; Brautigan, D.L.; Karaki, H.; Ozaki, Y.; Kato, Y.; Fusetani, N.; Watabe, S.; Hashimoto, K.; Uemura, D.; et al. Calyculin A and okadaic acid, inhibitors of protein phosphatases activity. Biochem. Biophys. Res. Commun. 1989, 159, 871–877. [Google Scholar] [CrossRef]
- Cohen, P.; Holmes, C.F.B.; Tsukitani, Y. Okadaic acid: A new probe for the study of cellular regulation. Trends Biochem. Sci. 1990, 15, 98–102. [Google Scholar] [CrossRef]
- Eriksson, J.E.; Brautigan, D.L.; Vallee, R.; Olmsted, J.; Fujiki, H.; Goldman, R.D. Cytoskeletal integrity in interphase cells requires protein phosphatase activity. Proc. Natl. Acad. Sci. USA 1992, 89, 11093–11097. [Google Scholar] [CrossRef]
- Gliksman, N.R.; Parsons, S.F.; Salmon, E.D. Okadaic acid induces interphase to mitotic-like microtubule dynamic instability by inactivating rescue. J. Cell Biol. 1992, 119, 1271–1276. [Google Scholar] [CrossRef]
- Vandré, D.D.; Wills, V.L. Inhibition of mitosis by okadaic acid: Possible involvement of a protein phosphatase 2A in the transition from metaphase to anaphase. J. Cell Sci. 1992, 101, 79–91. [Google Scholar]
- Furutani, I.; Watanabe, Y.; Prieto, R.; Masukawa, M.; Suzuki, K.; Naoi, K.; Thitamadee, S.; Shikanai, T.; Hashimoto, T. The SPIRAL genes are required for directional control of cell elongation in Arabidopsis thaliana. Development 2000, 127, 4443–4453. [Google Scholar]
- Sugimoto, K.; Himmelspach, R.; Williamson, R.E.; Wasteneys, G.O. Mutation or drug-dependent microtubule disruption causes radial swelling without altering parallel cellulose microfibril deposition in Arabidopsis root cells. Plant Cell Online 2003, 15, 1414–1429. [Google Scholar] [CrossRef]
- Ivanchenko, M.G.; Muday, G.K.; Dubrovsky, J.G. Ethylene-auxin interactions regulate lateral root initiation and emergence in Arabidopsis thaliana. Plant J. 2008, 55, 335–347. [Google Scholar] [CrossRef]
- Li, X.; Gruber, M.Y.; Hegedus, D.D.; Lydiate, D.J.; Gao, M.J. Effects of a coumarin derivative, 4-methylumbelliferone, on seed germination and seedling establishment in Arabidopsis. J. Chem. Ecol. 2011, 37, 880–890. [Google Scholar] [CrossRef]
- Laskowski, M.; Grieneisen, V.A.; Hofhuis, H.; Colette, A.; Hogeweg, P.; Marée, A.F.; Scheres, B. Root system architecture from coupling cell shape to auxin transport. PLoS Biol. 2008, 6, e307. [Google Scholar] [CrossRef]
- Mancuso, R.; Ziccarelli, I.; Armentano, D.; Marino, N.; Giofrè, S.V.; Gabriele, B. Divergent palladium-catalyzed multicomponent carbonylative approaches to functionalized isoidolinone and isobenzofuranimine derivatives. J. Org. Chem. 2014, 79, 3506–3518. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Streibig, J.C.; Kudsk, P.; Jensen, J.E. A general joint action model for herbicide mixtures. Pestic. Sci. 1998, 53, 21–28. [Google Scholar] [CrossRef]
- Belz, R.G.; Hurle, K.; Duke, S.O. Dose-response a challenge for allelopathy? Nonlinearity Biol. Toxicol. Med. 2005, 3, 173–211. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds 1 and 2 are available from the authors.
© 2014 by the authors. licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Araniti, F.; Mancuso, R.; Ziccarelli, I.; Sunseri, F.; Abenavoli, M.R.; Gabriele, B. 3-(Methoxycarbonylmethylene)isobenzofuran-1-imines as a New Class of Potential Herbicides. Molecules 2014, 19, 8261-8275. https://doi.org/10.3390/molecules19068261
Araniti F, Mancuso R, Ziccarelli I, Sunseri F, Abenavoli MR, Gabriele B. 3-(Methoxycarbonylmethylene)isobenzofuran-1-imines as a New Class of Potential Herbicides. Molecules. 2014; 19(6):8261-8275. https://doi.org/10.3390/molecules19068261
Chicago/Turabian StyleAraniti, Fabrizio, Raffaella Mancuso, Ida Ziccarelli, Francesco Sunseri, Maria Rosa Abenavoli, and Bartolo Gabriele. 2014. "3-(Methoxycarbonylmethylene)isobenzofuran-1-imines as a New Class of Potential Herbicides" Molecules 19, no. 6: 8261-8275. https://doi.org/10.3390/molecules19068261