Evaluating the Anti-Neuroinflammatory Capacity of Raw and Steamed Garlic as Well as Five Organosulfur Compounds

Abstract
:1. Introduction
2. Results and Discussion
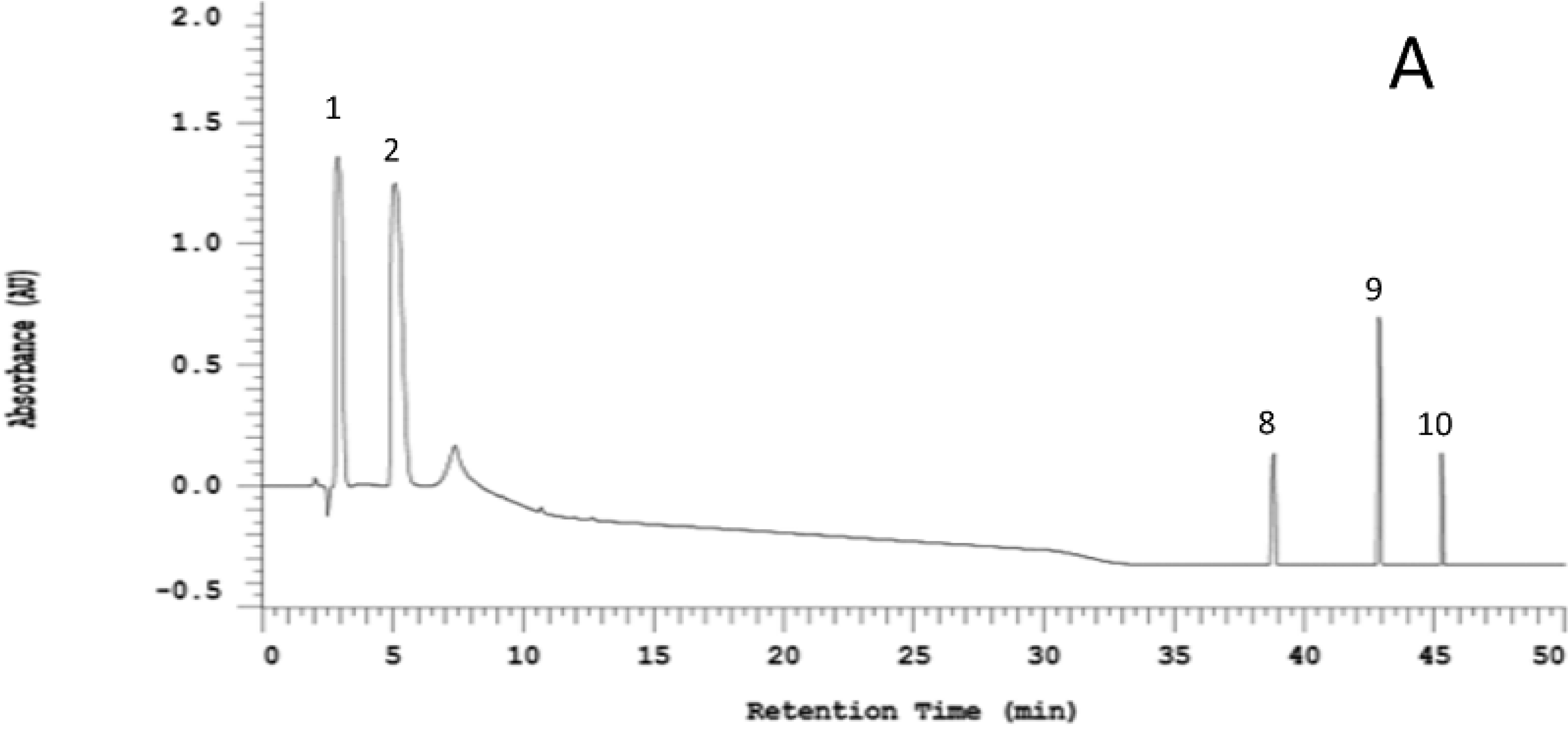
2.1. Organosulfur Compositional Changes after Steaming
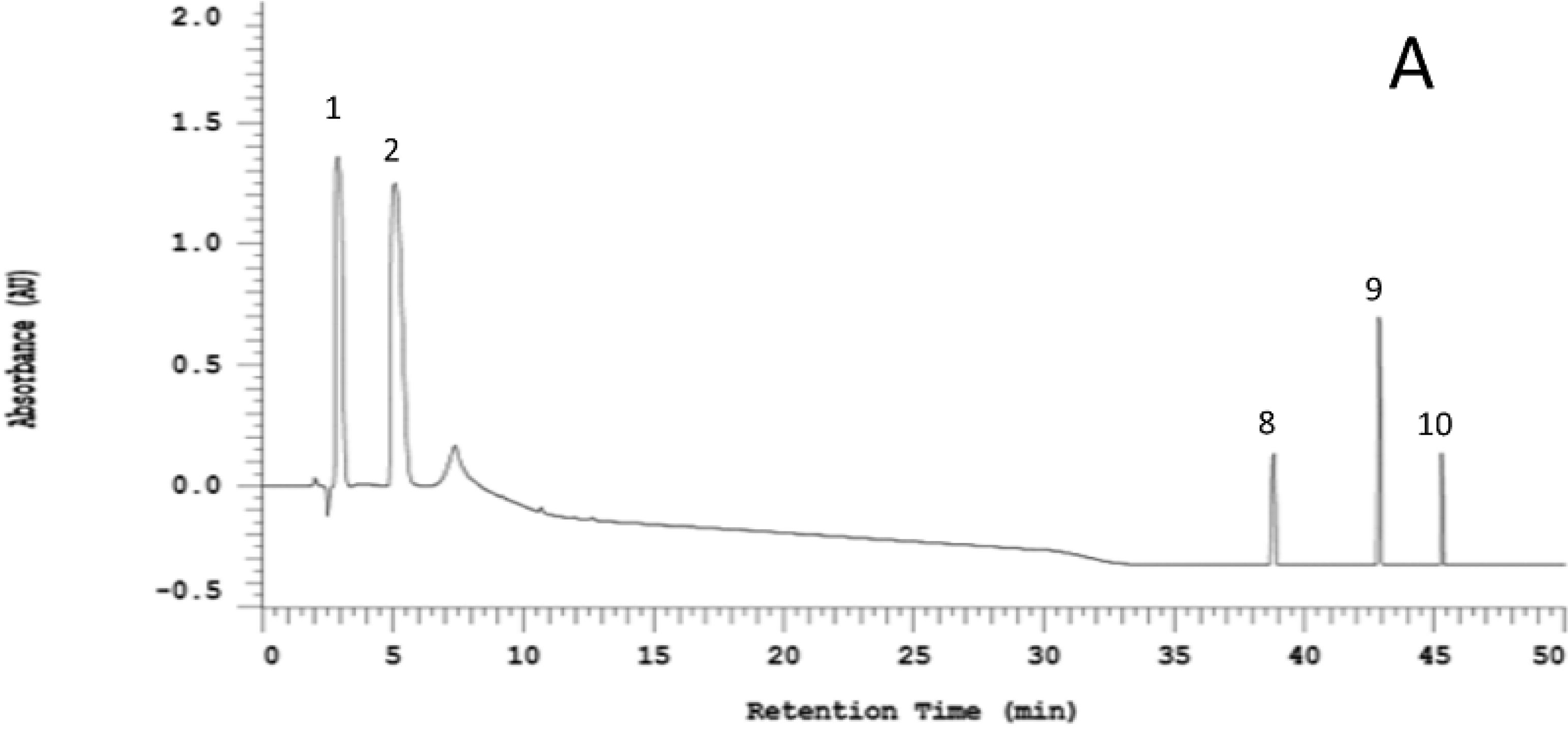
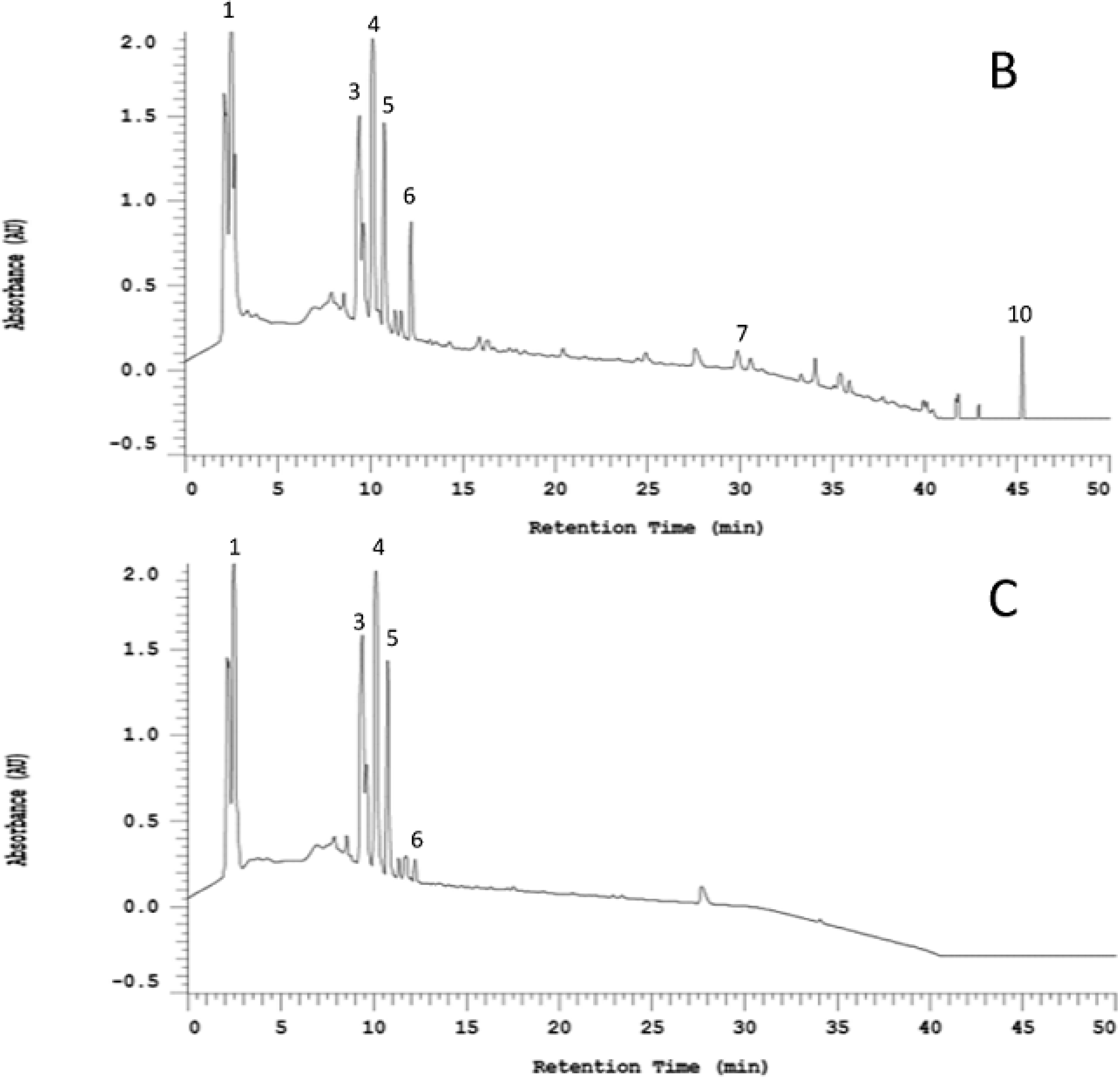
2.2. Anti-Neuroinflammatory Activities of Raw and Steamed Garlic Extracts
2.2.1. Inhibitory Effect of Raw and Steamed Garlic Extracts on NO Production and iNOS Gene Expression
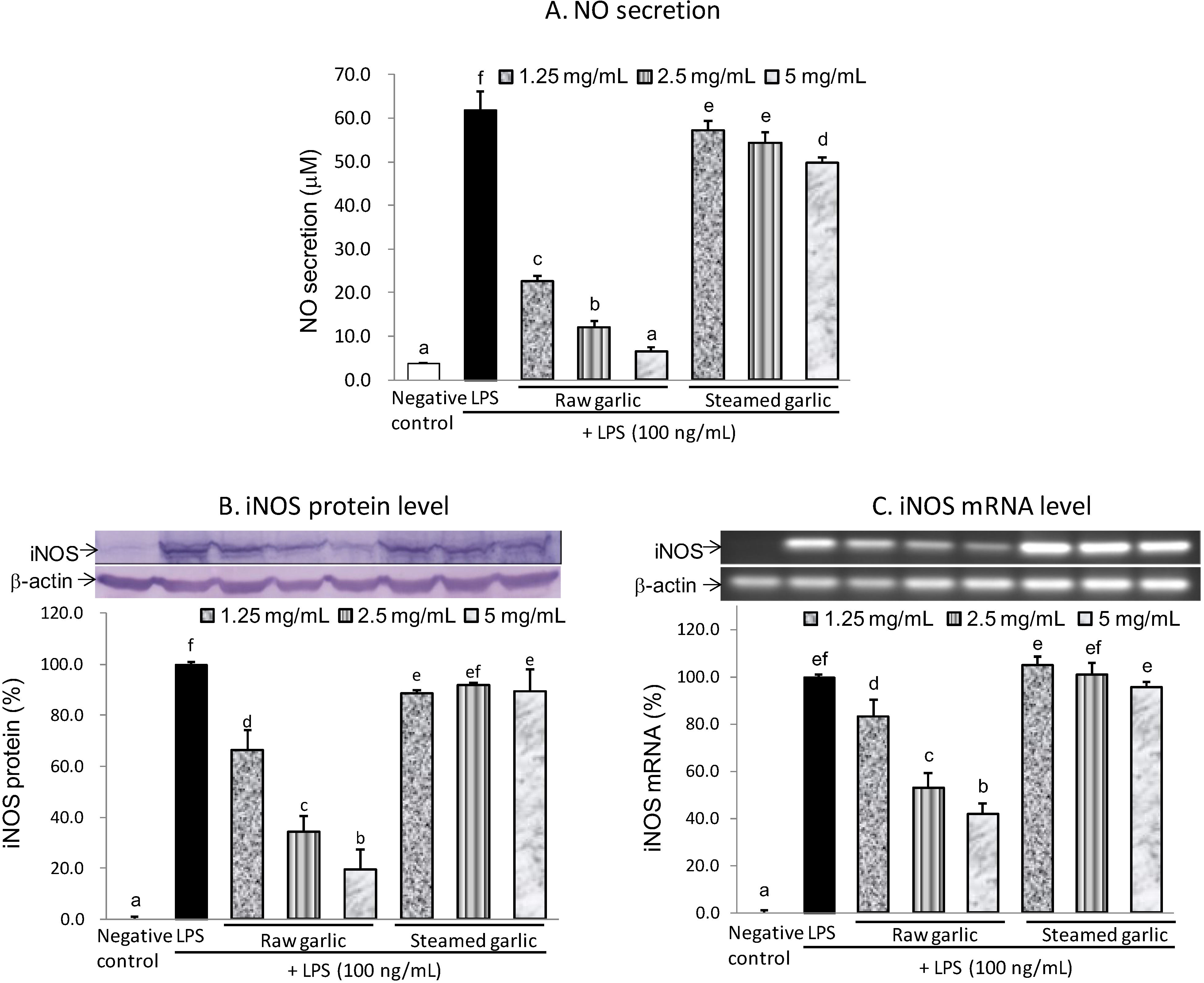

{kind=link}
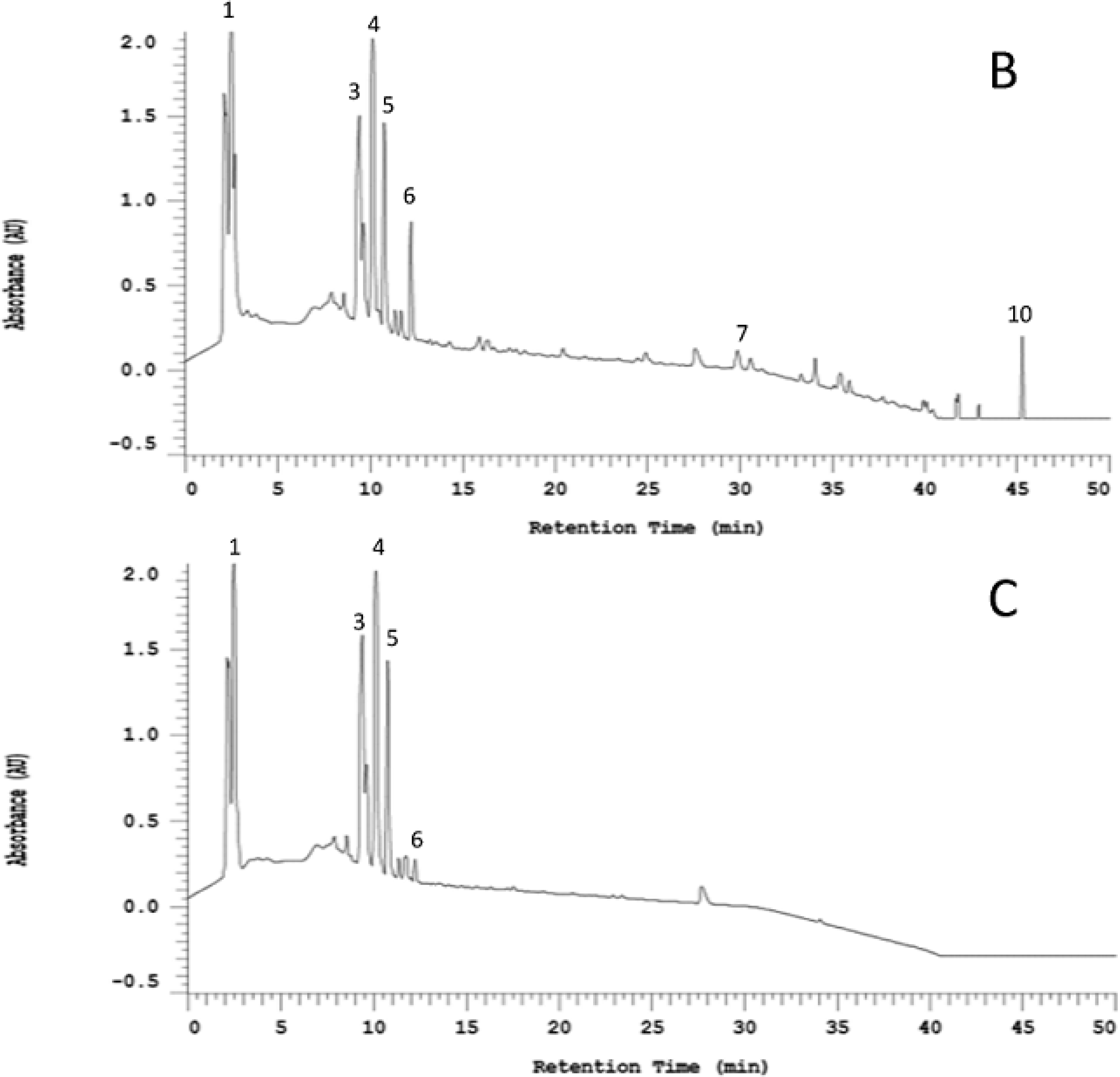{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Negative Control | LPS | Raw Garlic | Steamed Garlic | |||||
|---|---|---|---|---|---|---|---|---|
| 1.25 mg/mL | 2.5 mg/mL | 5.0 mg/mL | 1.25 mg/mL | 2.5 mg/mL | 5.0 mg/mL | |||
| Inhibition of NO secretion (%) | 100.0 | 0.0 | 67.6 | 85.5 | 95.2 | 8.0 | 12.7 | 20.6 |
| Inhibition of iNOS protein (%) | 100.0 | 0.0 | 33.8 | 65.8 | 80.7 | 11.5 | 8.3 | 10.8 |
| Inhibition of iNOS mRNA (%) | 100.0 | 0.0 | 16.9 | 47.3 | 58.2 | −5.1 | −1.1 | 4.3 |
| Inhibition of TNF−α secretion (%) | 100.0 | 0.0 | 10.5 | 36.9 | 66.0 | −2.2 | −0.6 | 9.1 |
| Inhibition of IL−1β secretion (%) | 100.0 | 0.0 | 14.2 | 31.9 | 49.1 | −5.4 | 7.1 | 8.5 |
| Inhibition of MCP−1 secretion (%) | 100.0 | 0.0 | 45.8 | 73.8 | 94.6 | 4.2 | 5.7 | 7.9 |
| Inhibition of TNF−α mRNA (%) | 100.0 | 0.0 | 4.3 | 18.8 | 62.6 | −2.8 | −2.8 | −1.0 |
| Inhibition of IL−1β mRNA (%) | 100.0 | 0.0 | −4.3 | 17.1 | 34.1 | 2.9 | 6.3 | 14.2 |
| Inhibition of MCP−1 mRNA (%) | 100.0 | 0.0 | 29.8 | 57.4 | 72.3 | −4.1 | −1.3 | 12.6 |
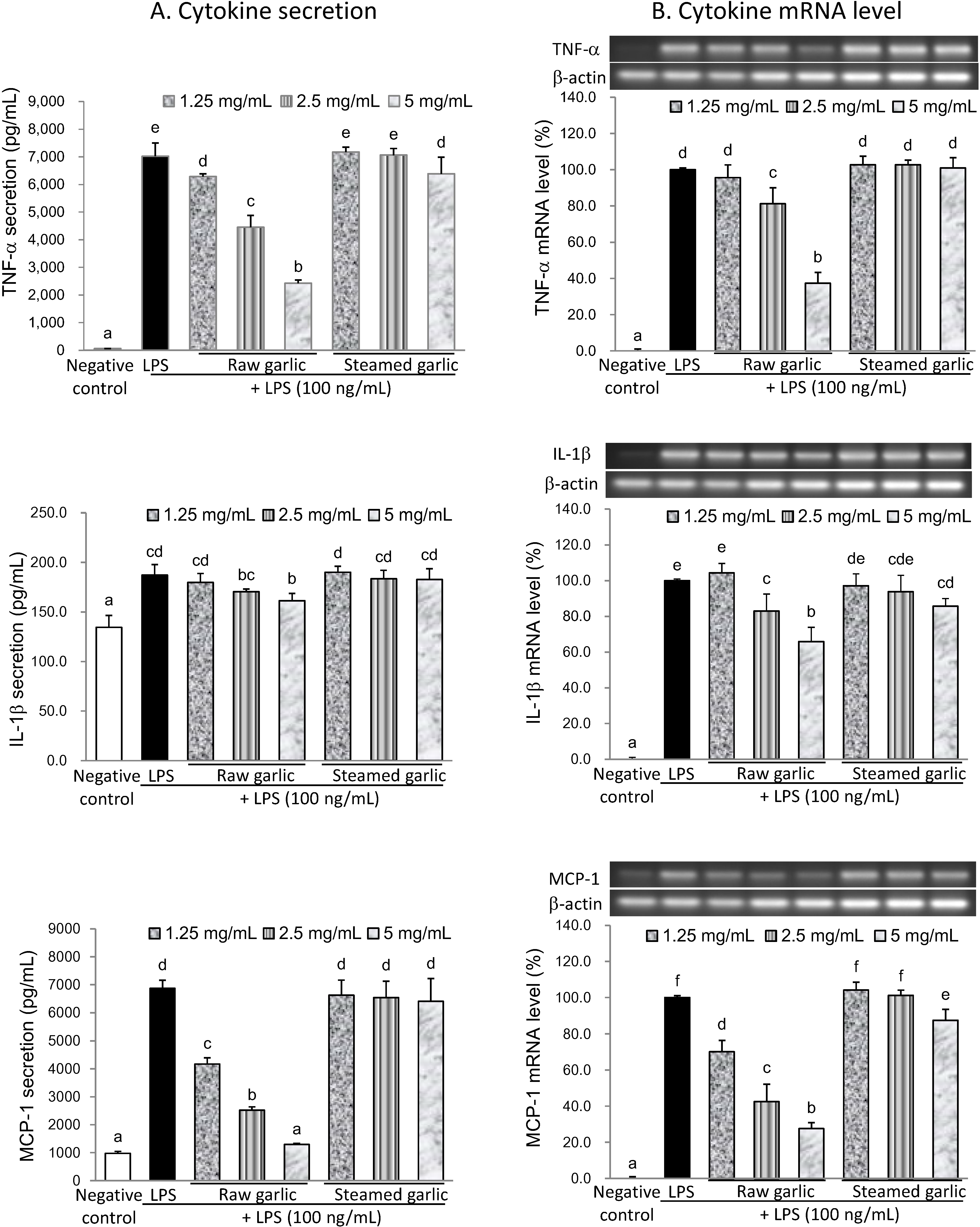
2.2.2. Inhibitory Effect of Raw and Steamed Garlic Extracts on Proinflammatory TNF-α, IL-1β, and MCP-1 Secretion and Gene Expression
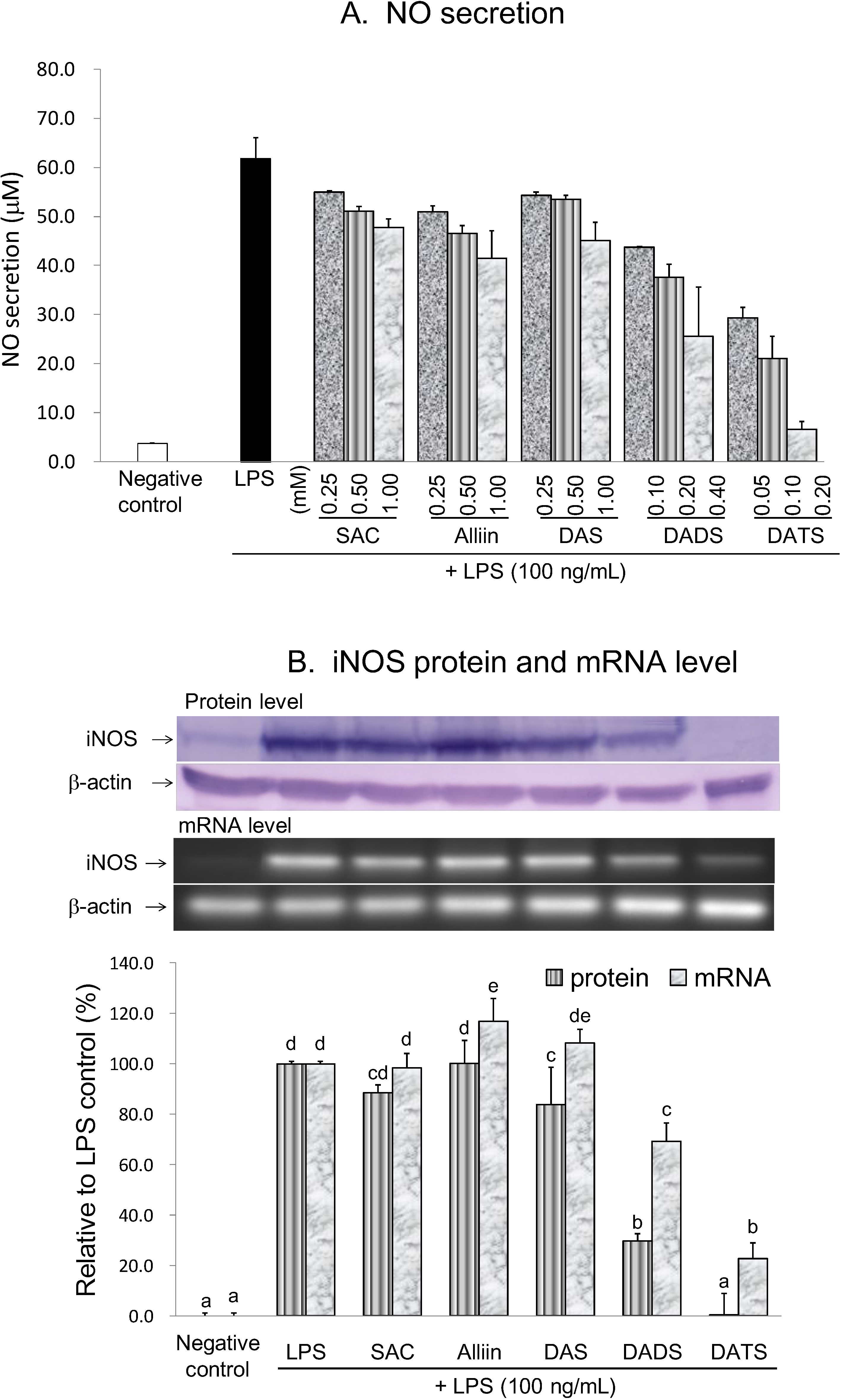
2.3. Anti-Neuroinflammatory Activities of OSCs
2.3.1. Inhibitory Effect of OSCs on NO Production and iNOS Gene Expression
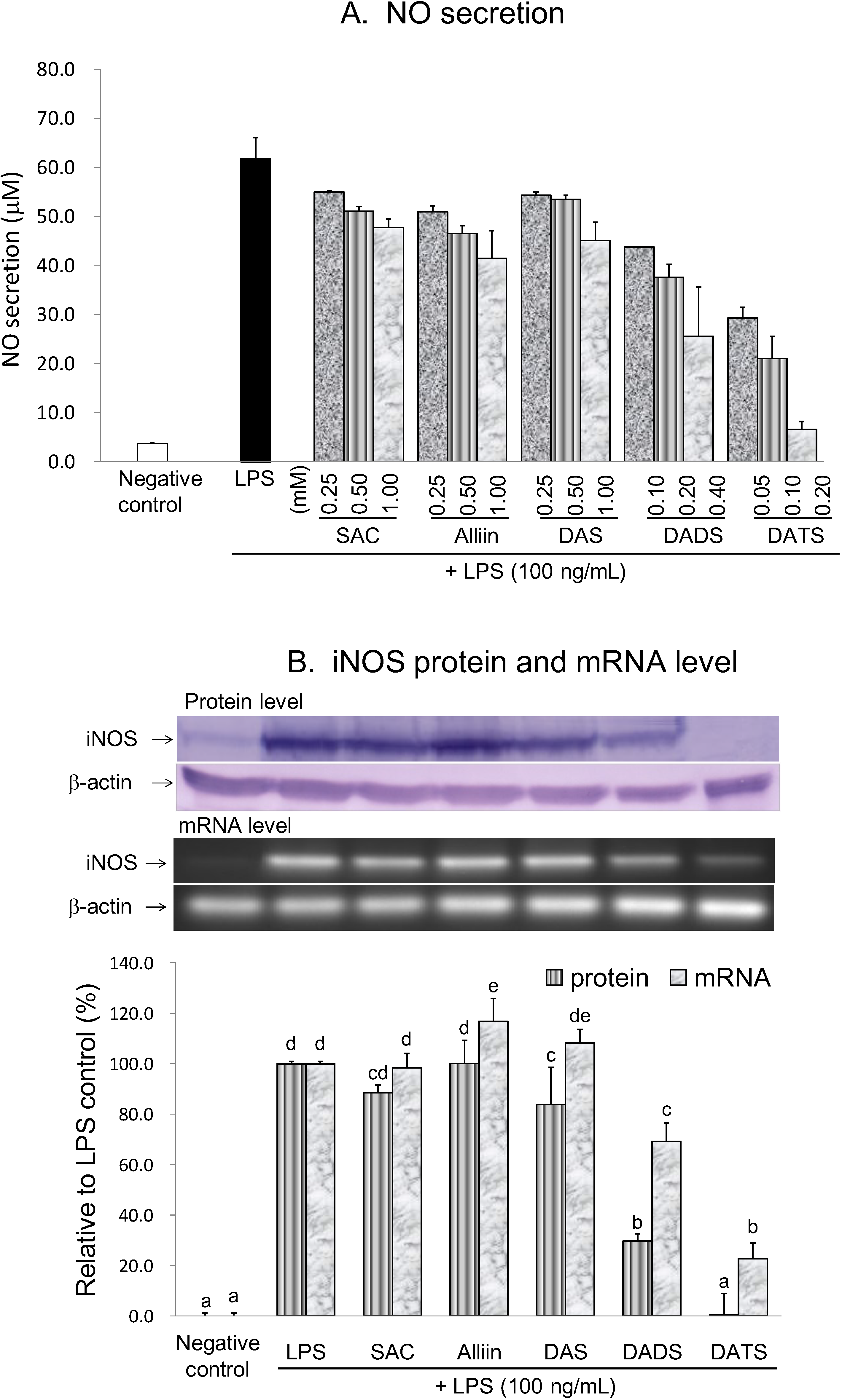
| Negative Control | LPS | SAC | Alliin | DAS | DADS | DATS | |
|---|---|---|---|---|---|---|---|
| 1 mM | 1 mM | 1 mM | 0.40 mM | 0.20 mM | |||
| Inhibition of NO secretion (%) | 100.0 | 0.0 | 24.2 | 35.1 | 28.9 | 62.4 | 94.9 |
| Inhibition of iNOS protein (%) | 100.0 | 0.0 | 11.6 | −0.2 | 16.2 | 70.2 | 99.7 |
| Inhibition of iNOS mRNA (%) | 100.0 | 0.0 | 1.6 | −16.9 | −8.3 | 30.8 | 77.4 |
| Inhibition of TNF-α secretion (%) | 100.0 | 0.0 | 10.1 | 19.9 | 25.6 | 42.4 | 67.4 |
| Inhibition of IL-1β secretion (%) | 100.0 | 0.0 | 30.3 | 27.1 | 19.1 | 44.0 | 69.5 |
| Inhibition of MCP-1 secretion (%) | 100.0 | 0.0 | 18.4 | 10.0 | 14.7 | 51.0 | 85.1 |
| Inhibition of TNF-α mRNA (%) | 100.0 | 0.0 | 16.2 | −11.9 | 9.0 | 43.6 | 79.5 |
| Inhibition of IL-1β mRNA (%) | 100.0 | 0.0 | 5.1 | −13.3 | −1.0 | 37.9 | 53.2 |
| Inhibition of MCP-1 mRNA (%) | 100.0 | 0.0 | 3.5 | −5.7 | 14.5 | 33.5 | 81.6 |
2.3.2. Inhibitory Effect of OSCs on TNF-α, IL-1β, and MCP-1 Secretion and Gene Expression

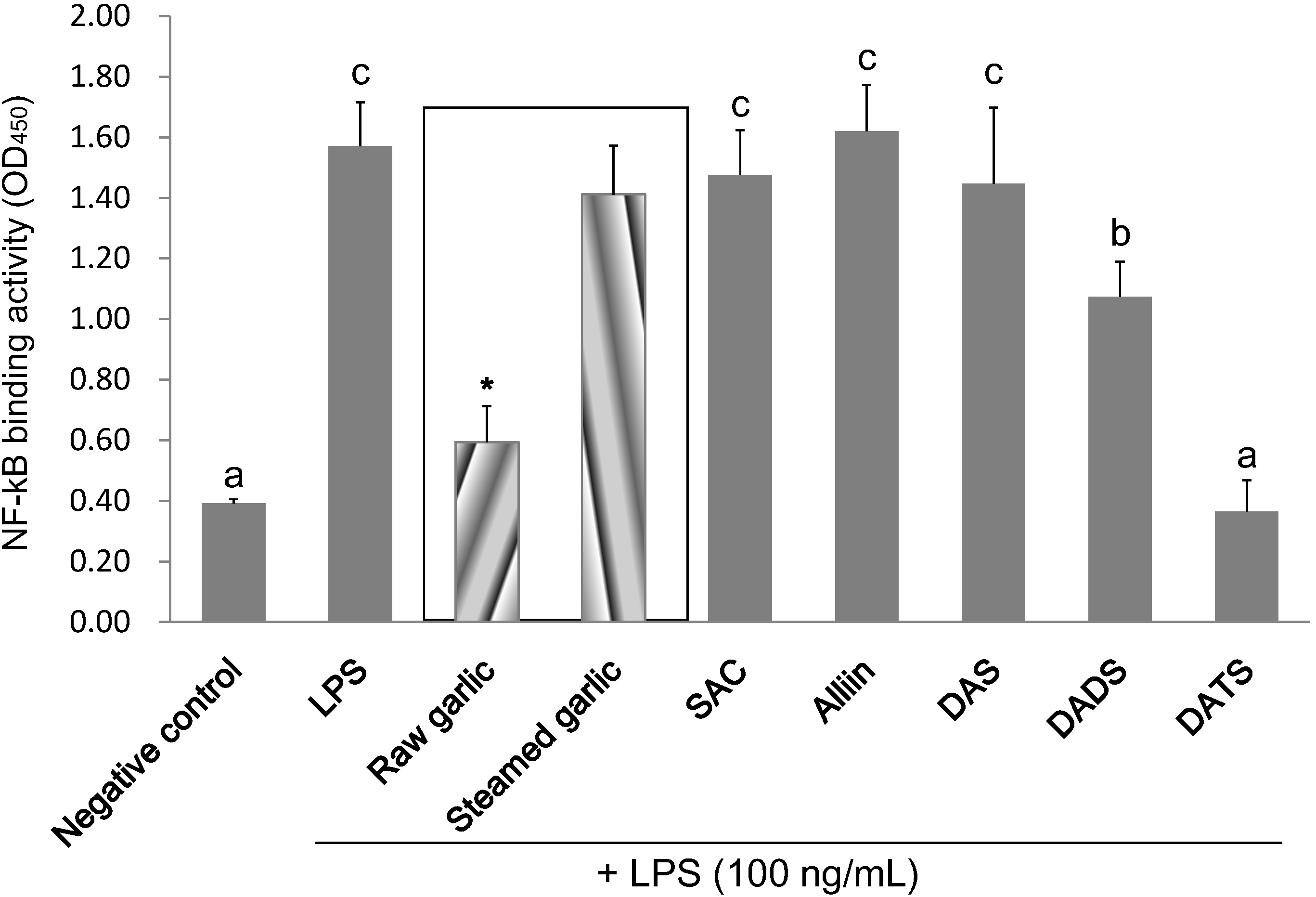
2.4. Inhibitory Effect of Garlic Extracts and OSCs on NF-κB Binding Activity
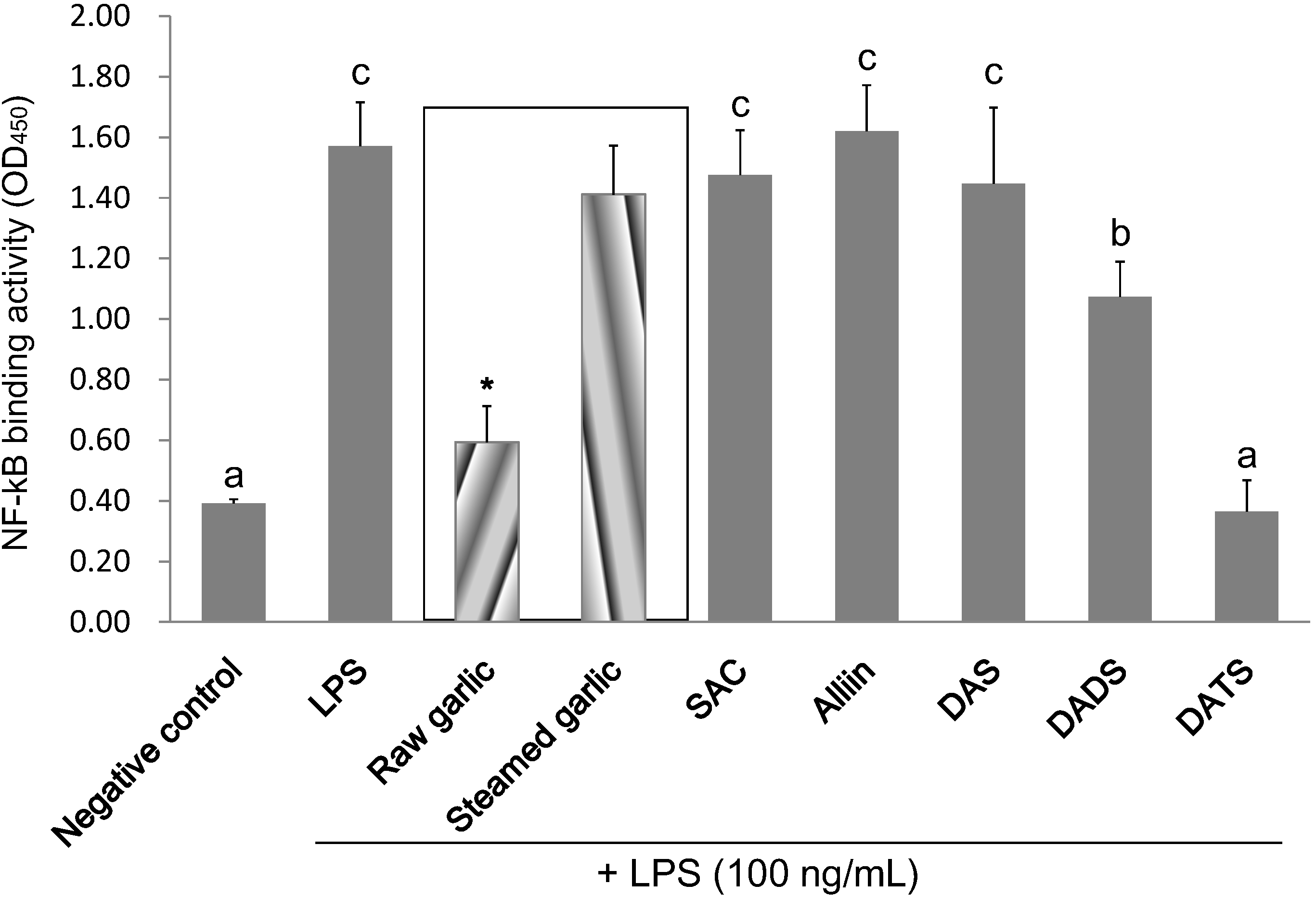
| Negative Control | LPS | Raw Garlic | Steamed Garlic | SAC | Alliin | DAS | DADS | DATS | |
|---|---|---|---|---|---|---|---|---|---|
| 5 mg/mL | 5 mg/mL | 1 mM | 1 mM | 1 mM | 0.4 mM | 0.2 mM | |||
| Inhibition of NF-κB activation (%) | 100.0 | 0.0 | 82.9 | 13.6 | 8.0 | −4.1 | 10.6 | 42.2 | 102.4 |
3. Experimental Section
3.1. Chemicals
3.2. Preparation of Raw- and Steamed-Garlic Ethanolic Extracts
3.3. HPLC and LC-MS Analyses
3.4. Cell Culture Experiment
3.5. Determination of Amounts of Secreted NO, TNF-α, IL-1β, and MCP-1
3.6. Evaluation of iNOS Protein Levels
3.7. Evaluation of iNOS, TNF-α, IL-1β, and MCP-1 mRNA Levels
3.8. Evaluation of NF-κB Binding Activity
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Amor, S.; Puentes, F.; Baker, D.; van der Valk, P. Inflammation in neurodegenerative diseases. Immunology 2010, 129, 154–169. [Google Scholar]
- Glass, C.K.; Saijo, K.; Winner, B.; Marchetto, M.C.; Gage, F.H. Mechanisms underlying inflammation in neurodegeneration. Cell 2010, 140, 918–934. [Google Scholar]
- Lynch, M.A. The multifaceted profile of activated microglia. Mol. Neurobiol. 2009, 40, 139–156. [Google Scholar]
- Rojo, L.E.; Fernández, J.A.; Maccioni, A.A.; Jimenez, J.M.; Maccioni, R.B. Neuroinflammation: Implications for the pathogenesis and molecular diagnosis of Alzheimer’s disease. Arch. Med. Res. 2008, 39, 1–16. [Google Scholar]
- Liu, B.; Hong, J.S. Role of microglia in inflammation-mediated neurodegenerative diseases: Mechanisms and strategies for therapeutic intervention. J. Pharmacol. Exp. Ther. 2003, 304, 1–7. [Google Scholar]
- Choi, D.K.; Koppula, S.; Suk, K. Inhibitors of microglial neurotoxicity: Focus on natural products. Molecules 2011, 16, 1021–1043. [Google Scholar]
- Kannappan, R.; Gupta, S.C.; Kim, J.H.; Reuter, S.; Aggarwal, B.B. Neuroprotection by spice-derived nutraceuticals: You are what you eat! Mol. Neurobiol. 2011, 44, 142–159. [Google Scholar]
- Agarwal, K.C. Therapeutic actions of garlic constituents. Med. Res. Rev. 1996, 16, 111–124. [Google Scholar]
- Santhosha, S.G.; Jamuna, P.; Prabhavathi, S.N. Bioactive components of garlic and their physiological role in health maintenance: A review. Food Biosci. 2013, 3, 59–74. [Google Scholar]
- Yun, H.M.; Ban, J.O.; Park, K.R.; Lee, C.K.; Jeong, H.S.; Han, S.B.; Hong, J.T. Potential therapeutic effects of functionally active compounds isolated from garlic. Pharmacol. Ther. 2014, 142, 183–195. [Google Scholar]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive compounds. J. Nutr. 2001, 131, 955S–962S. [Google Scholar]
- Geng, Z.; Rong, Y.; Lau, B.H. S-allyl cysteine inhibits activation of nuclear factor kappa B in human T cells. Free Radic. Biol. Med. 1997, 23, 345–350. [Google Scholar]
- Dirsch, V.M.; Kiemer, A.K.; Wagner, H.; Vollmar, A.M. Effect of allicin and ajoene, two compounds of garlic, on inducible nitric oxide synthase. Atherosclerosis 1998, 139, 333–339. [Google Scholar]
- Liu, K.L.; Chen, H.W.; Wang, R.Y.; Lei, Y.P.; Sheen, L.Y.; Lii, C.K. DATS reduces LPS-induced iNOS expression, NO production, oxidative stress, and NF-κB activation in RAW 264.7 macrophages. J. Agric. Food Chem. 2006, 54, 3472–3478. [Google Scholar]
- Shin, J.-H.; Ryu, J.H.; Kang, M.J.; Hwang, C.R.; Han, J.; Kang, D. Short-term heating reduces the anti-inflammatory effects of fresh raw garlic extracts on the LPS-induced production of NO and pro-inflammatory cytokines by downregulating allicin activity in RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 58, 545–551. [Google Scholar]
- Song, K.; Milner, J.A. The influence of heating on the anticancer properties of garlic. J. Nutr. 2001, 131, 1054S–1057S. [Google Scholar]
- Arnault, I.; Christides, J.P.; Mandon, N.; Haffner, T.; Kahane, R.; Auger, J. High-performance ion-pair chromatography method for simultaneous analysis of alliin, deoxyalliin, allicin and dipeptide precursors in garlic products using multiple mass spectrometry and UV detection. J. Chromatogr. A 2003, 991, 69–75. [Google Scholar]
- Gupta, V.B.; Indi, S.S.; Rao, K.S. Garlic extract exhibits antiamyloidogenic activity on amyloid-β fibrillogenesis: Relevance to Alzheimer’s disease. Phytother. Res. 2009, 23, 111–115. [Google Scholar]
- Zhang, G.; Parkin, K.L. S-Alk(en)ylmercaptocysteine: Chemical synthesis, biological activities, and redox-related mechanism. J. Agric. Food Chem. 2013, 61, 1896–1903. [Google Scholar]
- Fukao, T.; Hosono, T.; Misawa, S.; Seki, T.; Ariga, T. The effects of allyl sulfides on the induction of phase II detoxification enzymes and liver injury by carbon tetrachloride. Food Chem. Toxicol. 2004, 42, 743–749. [Google Scholar]
- Lai, K.C.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Ma, C.Y.; Lu, H.F.; Tang, N.Y.; Hsia, T.C.; Ho, H.C.; Chung, J.G. Diallyl sulfide, diallyl disulfide, and diallyl trisulfide inhibit migration and invasion in human colon cancer colo 205 cells through the inhibition of matrix metalloproteinase-2, -7, and -9 expressions. Environ. Toxicol. 2013, 28, 479–488. [Google Scholar]
- Lei, Y.P.; Chen, H.W.; Sheen, L.Y.; Lii, C.K. Diallyl disulfide and diallyl trisulfide suppress oxidized LDL-induced vascular cell adhesion molecule and E-selectin expression through protein kinase A- and B-dependent signaling pathways. J. Nutr. 2008, 138, 996–1003. [Google Scholar]
- Xiao, D.; Choi, S.; Johnson, D.E.; Vogel, V.G.; Johnson, C.S.; Trump, D.L.; Lee, Y.J.; Singh, S.V. Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 2004, 23, 5594–5606. [Google Scholar]
- Youn, H.-S.; Lim, H.J.; Lee, H.J.; Hwang, D.; Yang, M.; Jeon, R.; Ryu, J.-H. Garlic (Allium sativum) extract inhibits lipopolysaccharide-induced toll-like receptor 4 dimerization. Biosci. Biotechnol. Biochem. 2008, 72, 368–375. [Google Scholar]
- Park, H.Y.; Kim, N.D.; Kim, G.Y.; Hwang, H.J.; Kim, B.W.; Kim, W.J.; Choi, Y.H. Inhibitory effects of diallyl disulfide on the production of inflammatory mediators and cytokines in lipopolysaccharide-activated BV2 microglia. Toxicol. Appl. Pharmacol. 2012, 262, 177–184. [Google Scholar]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Scott Isbell, T.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar]
- Munday, R.; Munday, J.S.; Munday, C.M. Comparative effects of mono-, di-, tri-, and tetrasulfides derived from plants of the Allium family: Redox cycling in vitro and hemolytic activity and phase 2 enzyme induction in vivo. Free Radic. Biol. Med. 2003, 34, 1200–1211. [Google Scholar]
- Hosono, T.; Fukao, T.; Ogihara, J.; Ito, Y.; Shiba, H.; Seki, T.; Ariga, T. Diallyl trisulfide suppresses the proliferation and induces apoptosis of human colon cancer cells through oxidative modification of β-tubulin. J. Biol. Chem. 2005, 280, 41487–41493. [Google Scholar]
- Kim, S.; Lee, H.G.; Park, S.A.; Kundu, J.K.; Keum, Y.S.; Cha, Y.N.; Na, H.K.; Surh, Y.J. Keap1 cysteine 288 as a potential target for diallyl trisulfide-induced Nrf2 activation. PLoS One 2014, 9, e85984. [Google Scholar]
- Matsunaga, N.; Tsuchimori, N.; Matsumoto, T.; Ii, M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol. Pharmacol. 2011, 79, 34–41. [Google Scholar]
- Mukherjee, D.; Banerjee, S. Learning and memory promoting effects of crude garlic extract. Indian J. Exp. Biol. 2013, 51, 1094–1100. [Google Scholar]
- Chauhan, N.B. Effect of aged garlic extract on APP processing and tau phosphorylation in Alzheimer’s transgenic model Tg2576. J. Ethnopharmacol. 2006, 108, 385–394. [Google Scholar]
- Guo, Y.; Zhang, K.; Wang, Q.; Li, Z.; Yin, Y.; Xu, Q.; Duan, W.; Li, C. Neuroprotective effects of diallyl trisulfide in SOD1-G93A transgenic mouse model of amyotrophic lateral sclerosis. Brain Res. 2010, 1374, 110–115. [Google Scholar]
- Amagase, H. Clarifying the real bioactive constituents of garlic. J. Nutr. 2006, 136, 716S–725S. [Google Scholar]
- Jeong, J.H.; Jeong, H.R.; Jo, Y.N.; Kim, H.J.; Shin, J.H.; Heo, H.J. Ameliorating effects of aged garlic extracts against Aβ-induced neurotoxicity and cognitive impairment. BMC Complement. Altern. Med. 2013, 13, 268–278. [Google Scholar]
- Sample Availability: Samples of the compounds, alliin, SAC, DAS, DADS and DATS are not availabele from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ho, S.-C.; Su, M.-S. Evaluating the Anti-Neuroinflammatory Capacity of Raw and Steamed Garlic as Well as Five Organosulfur Compounds. Molecules 2014, 19, 17697-17714. https://doi.org/10.3390/molecules191117697
Ho S-C, Su M-S. Evaluating the Anti-Neuroinflammatory Capacity of Raw and Steamed Garlic as Well as Five Organosulfur Compounds. Molecules. 2014; 19(11):17697-17714. https://doi.org/10.3390/molecules191117697
Chicago/Turabian StyleHo, Su-Chen, and Min-Sheng Su. 2014. "Evaluating the Anti-Neuroinflammatory Capacity of Raw and Steamed Garlic as Well as Five Organosulfur Compounds" Molecules 19, no. 11: 17697-17714. https://doi.org/10.3390/molecules191117697
APA StyleHo, S.-C., & Su, M.-S. (2014). Evaluating the Anti-Neuroinflammatory Capacity of Raw and Steamed Garlic as Well as Five Organosulfur Compounds. Molecules, 19(11), 17697-17714. https://doi.org/10.3390/molecules191117697