Toxicity Assessments of Chalcone and Some Synthetic Chalcone Analogues in a Zebrafish Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
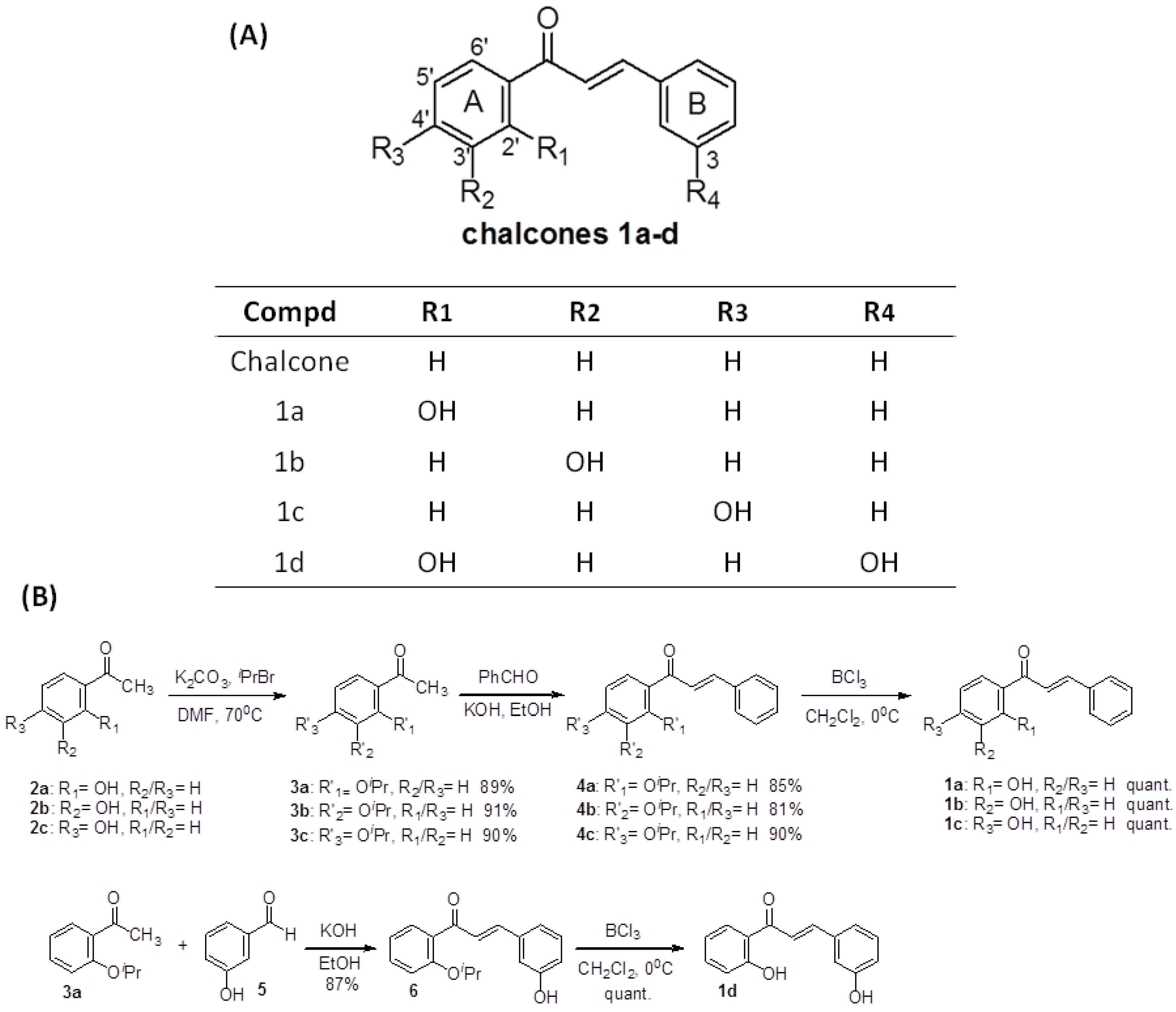
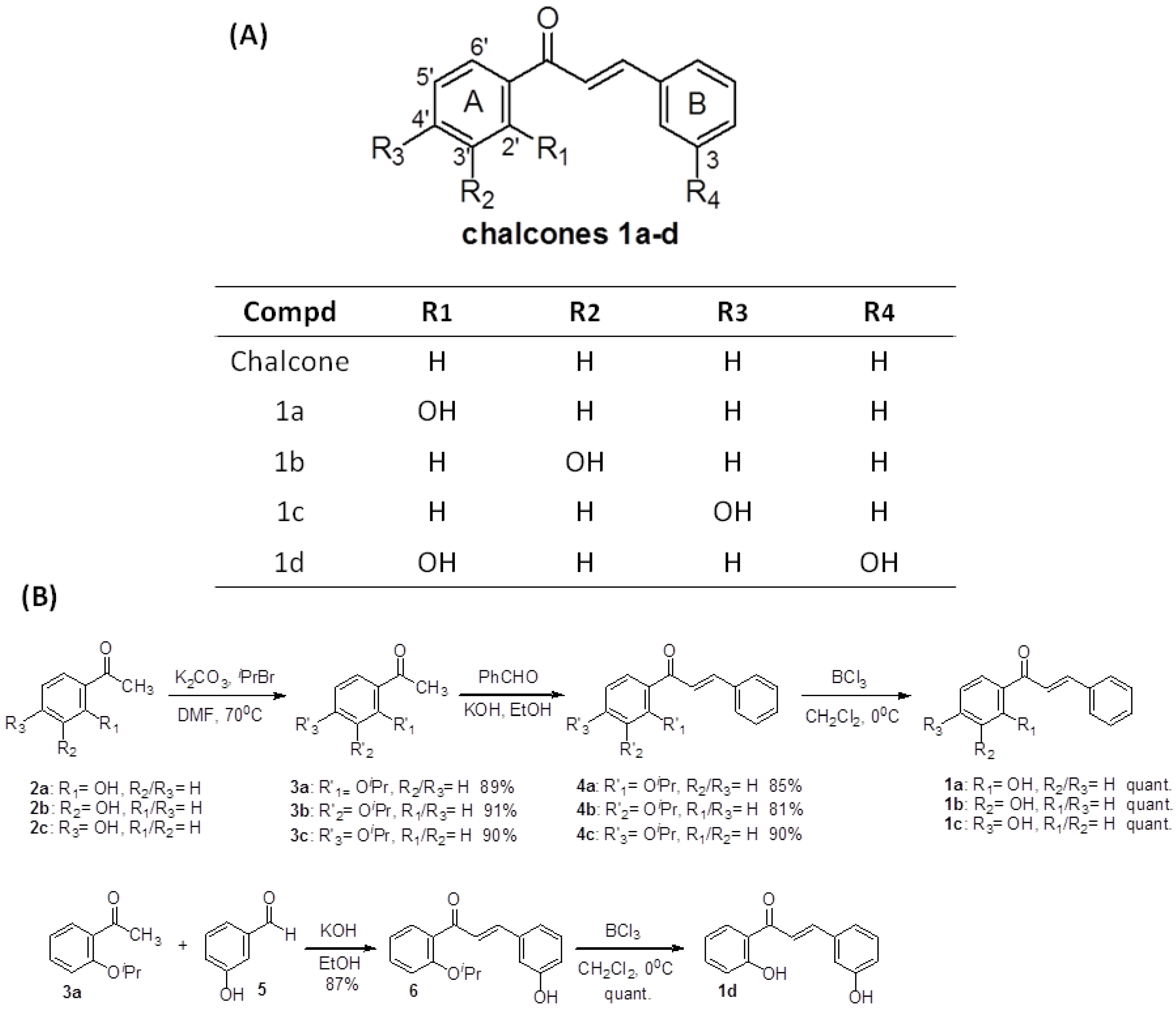
2.1. Chemistry
2.2. Titration and Survival Rates Analysis

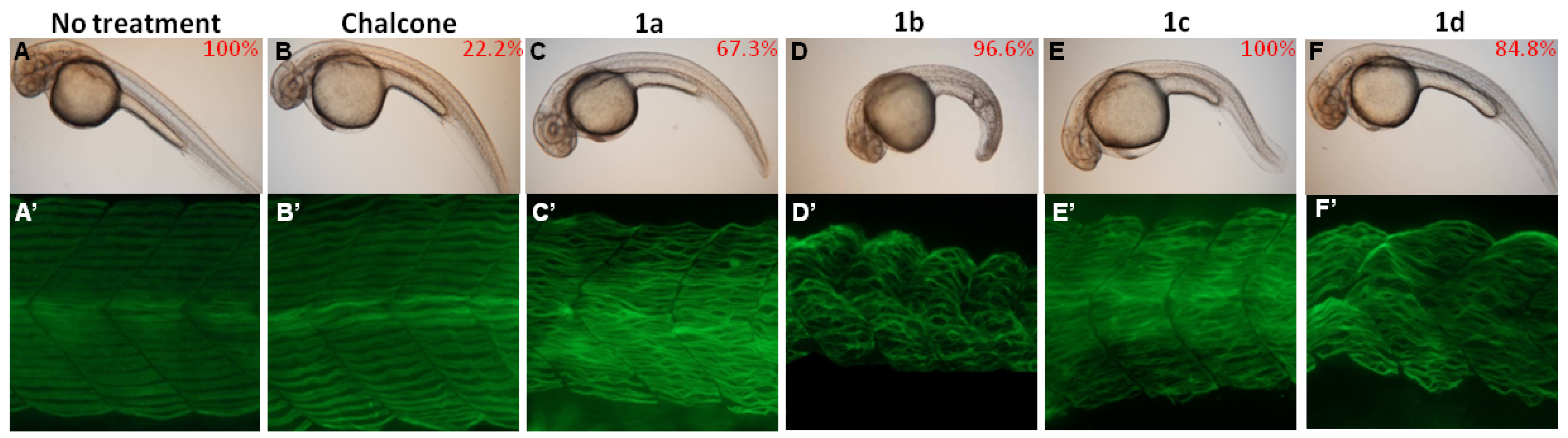
2.3. Phenotypic Changes after Chalcone Treatment
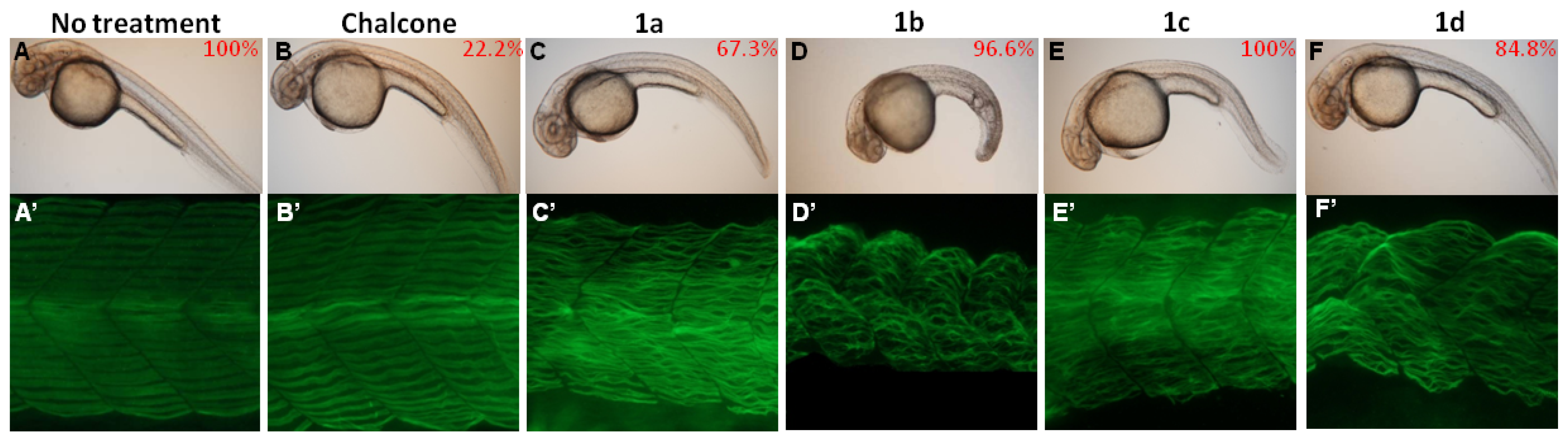
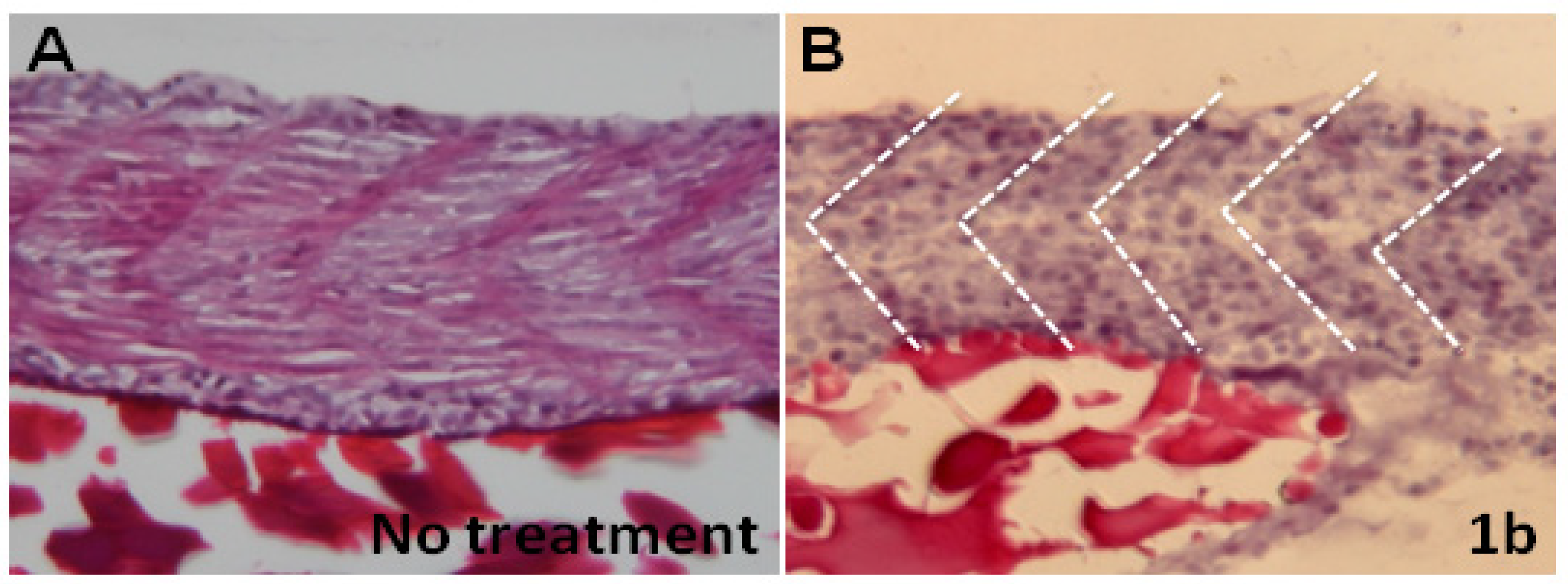
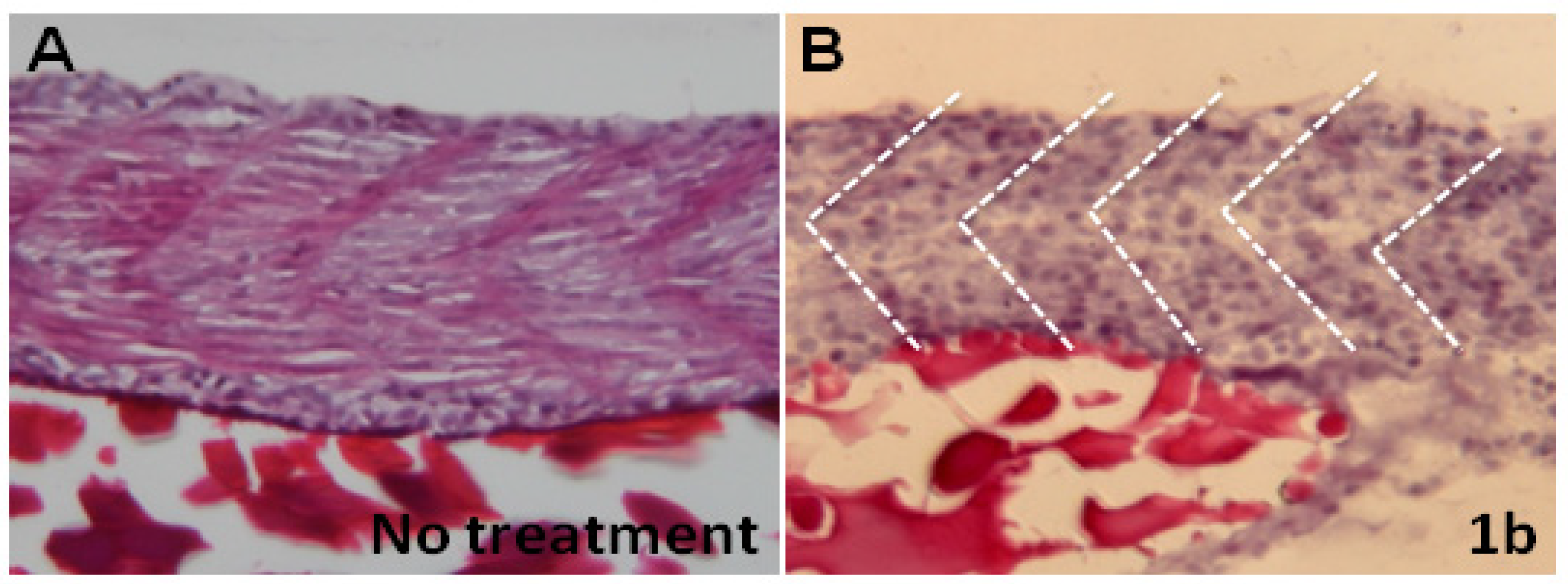
2.4. Chalcones Affect Myofibril Ultrastructures and Alignment

3. Experimental
3.1. General Information
3.2. Synthesis of Chalcone Analogues
3.3. Fish Care and Chemicals Treatment
3.4. Histology, Antibody Labeling and Images
3.5. Electron Microscopy
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Mohamad, A.S.; Akhtar, M.N.; Zakaria, Z.A.; Perimal, E.K.; Khalid, S.; Mohd, P.A.; Khalid, M.H.; Israf, D.A.; Lajis, N.H.; Sulaiman, M.R. Antinociceptive activity of a synthetic chalcone, flavokawin B on chemical and thermal models of nociception in mice. Eur. J. Pharmacol. 2010, 647, 103–109. [Google Scholar] [CrossRef]
- Wu, J.; Lee, J.; Cai, Y.; Pan, Y.; Ye, F.; Zhang, Y.; Zhao, Y.; Yang, S.; Li, X.; Liang, G. Evaluation and discovery of novel synthetic chalcone derivatives as anti-inflammatory agents. J. Med. Chem. 2011, 54, 8110–8123. [Google Scholar] [CrossRef]
- Ajaiyeoba, E.O.; Ogbole, O.O.; Abiodun, O.O.; Ashidi, J.S.; Houghton, P.J.; Wright, C.W. Cajachalcone: An antimalarial compound from Cajanus cajan leaf extract. J. Parasitol. Res. 2013, 2013, 703781. [Google Scholar]
- Ajiboye, T.O.; Yakubu, M.T.; Oladiji, A.T. Electrophilic and reactive oxygen species detoxification potentials of chalcone dimers is mediated by redox transcription factor Nrf-2. J. Biochem. Mol. Toxicol. 2013. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wang, W.H.; Wang, Y.H.; Lin, Z.Y.; Wen, C.C.; Chern, C.Y. Evaluation of anti-inflammatory effect of chalcone and chalcone analogues in a zebrafish model. Molecules 2013, 18, 2052–2060. [Google Scholar] [CrossRef]
- Pan, Y.; Chen, Y.; Li, Q.; Yu, X.; Wang, J.; Zheng, J. The synthesis and evaluation of novel hydroxyl substituted chalcone analogs with in vitro anti-free radicals pharmacological activity and in vivo anti-oxidation activity in a free radical-injury Alzheimer’s model. Molecules 2013, 18, 1693–1703. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, X.; Wu, G.; Yang, X.; Pan, S.; Wang, Y.; Ruan, J. Chalcone derivatives from the fern Cyclosorus parasiticus and their anti-proliferative activity. Food Chem. Toxicol. 2013, 60, 147–152. [Google Scholar] [CrossRef]
- Sashidhara, K.V.; Kumar, A.; Kumar, M.; Sarkar, J.; Sinha, S. Synthesis and in vitro evaluation of novel coumarin-chalcone hybrids as potential anticancer agents. Bioorg. Med. Chem. Lett. 2010, 20, 7205–7211. [Google Scholar] [CrossRef]
- Fang, X.; Yang, B.; Cheng, Z.; Yang, M.; Su, N.; Zhou, L.; Zhou, J. Synthesis and antitumor activity of novel mustard-linked chalcones. Arch. Pharm. 2013, 346, 292–299. [Google Scholar] [CrossRef]
- Kamal, A.; Kashi Reddy, M.; Viswanath, A. The design and development of imidazothiazole-chalcone derivatives as potential anticancer drugs. Expert Opin. Drug Discov. 2013, 8, 289–304. [Google Scholar] [CrossRef]
- Neves, M.P.; Lima, R.T.; Choosang, K.; Pakkong, P.; de São José Nascimento, M.; Vasconcelos, M.H.; Pinto, M.; Silva, A.M.; Cidade, H. Synthesis of a natural chalcone and its prenyl analogs—Evaluation of tumor cell growth-inhibitory activities, and effects on cell cycle and apoptosis. Chem. Biodivers. 2012, 9, 1133–1143. [Google Scholar] [CrossRef]
- Shin, S.Y.; Yoon, H.; Ahn, S.; Kim, D.W.; Kim, S.H.; Koh, D.; Lee, Y.H.; Lim, Y. Chromenylchalcones showing cytotoxicity on human colon cancer cell lines and in siloco docking with aurora kinases. Bioorg. Med. Chem. 2013, 21, 4250–4258. [Google Scholar] [CrossRef]
- De Vasconcelos, A.; Campos, V.F.; Nedel, F.; Seixas, F.K.; Dellagostin, O.A.; Smith, K.R.; de Pereira, C.M.; Stefanello, F.M.; Collares, T.; Barschak, A.G. Cytotoxic and apoptotic effects of chalcone derivatives of 2-acetylthiophene on human colon adenocarcinoma cells. Cell Biochem. Funct. 2013, 31, 289–297. [Google Scholar]
- Forejtníková, H.; Lunerová, K.; Kubínová, R.; Jankovská, D.; Marek, R.; Suchŷ, V.; Vondrácek, J. Chemoprotective and toxic potentials of synthetic and natural chalcones and dihydrochalcones in vitro. Toxicology 2005, 208, 81–93. [Google Scholar] [CrossRef]
- Waalkens-Berendsen, D.H.; Kuilman-Wahls, M.E.; Bär, A. Embryotoxicity and teratogenicity study with neohesperidin dihydrochalcone in rats. Regul. Toxicol. Pharmacol. 2004, 40, 74–79. [Google Scholar] [CrossRef]
- Ohkatsu, Y.; Satoh, T. Antioxidant and photo-antioxidant activities of chalcone derivatives. J. Jpn. Pet. Inst. 2008, 51, 298–308. [Google Scholar] [CrossRef]
- Karki, R.; Thapa, P.; Kang, M.J.; Jeong, T.C.; Nam, J.M.; Kim, H.L.; Na, Y.; Cho, W.J.; Kwon, Y.; Lee, E.S. Synthesis, topoisomerase I and II inhibitory activity, cytotoxicity, and structure-activity relationship study of hydroxylated 2,4-diphenyl-6-aryl pyridines. Bioorg. Med. Chem. 2010, 18, 3066–3077. [Google Scholar] [CrossRef]
- Kurniadewi, F.; Juliawaty, L.D.; Syah, Y.M.; Achmad, S.A.; Hakim, E.H.; Koyama, K.; Kinoshita, K.; Takahashi, K. Phenolic compounds from Cryptocarya konishii: Their cytotoxic and tyrosine kinase inhibitory properties. J. Nat. Med. 2010, 64, 121–125. [Google Scholar] [CrossRef]
- Qian, Y.P.; Shang, Y.J.; Teng, Q.F.; Chang, J.; Fan, G.J.; Wei, X.; Li, R.R.; Li, H.P.; Yao, X.J.; Dai, F.; et al. Hydroxychalcones as potent antioxidants: Structure-activity relationship analysis and mechanism considerations. Food Chem. 2011, 126, 241–248. [Google Scholar] [CrossRef]
- Moorthy, N.S.H.N.; Singh, R.J.; Singh, H.P.; Gupta, S.D. Synthesis, biological evaluation and in silico metabolic and toxicity prediction of some flavanone derivatives. Chem. Pharm. Bull. 2006, 54, 1384–1390. [Google Scholar] [CrossRef]
- Karki, R.; Thapa, P.; Yoo, H.Y.; Kadayat, T.M.; Park, P.H.; Na, Y.; Lee, E.; Jeon, K.H.; Cho, W.J.; Choi, H.; et al. Dihydroxylated 2,4,6-triphenyl pyridines: Synthesis, topoisomerase I and II inhibitory activity, cytotoxicity, and structure-activity relationship study. Eur. J. Med. Chem. 2012, 49, 219–228. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book, 3rd ed.; University of Oregon Press: Eugene, OR, USA, 1995. [Google Scholar]
- Chen, Y.H.; Wang, Y.H.; Yu, T.H.; Wu, H.J.; Pai, C.W. Transgenic zebrafish line with over-expression of Hedgehog on the skin: A useful tool to screen Hedgehog-inhibiting compounds. Transgenic Res. 2009, 18, 855–864. [Google Scholar] [CrossRef]
- Wang, Y.H.; Cheng, C.C.; Lee, W.J.; Chiou, M.L.; Pai, C.W.; Wen, C.C.; Chen, W.L.; Chen, Y.H. A novel phenotype-based approach for systematically screening antiproliferation metallodrugs. Chem. Biol. Interact. 2009, 182, 84–91. [Google Scholar] [CrossRef]
- Kimmel, C.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development in the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Chen, Y.H.; Lin, Y.T.; Lee, G.H. Novel and unexpected functions of zebrafish CCAAT box binding transcription factor (NF-Y) B subunit during cartilages development. Bone 2009, 44, 777–784. [Google Scholar] [CrossRef]
- Lee, G.H.; Chang, M.Y.; Hsu, C.H.; Chen, Y.H. Essential roles of basic helix-loop-helix transcription factors, Capsulin and Musculin, during craniofacial myogenesis of zebrafish. Cell Mol. Life Sci. 2011, 68, 4065–4078. [Google Scholar] [CrossRef]
- Chen, Y.H.; Chang, C.Y.; Wang, Y.H.; Wen, C.C.; Chen, Y.C.; Hu, S.C.; Yu, D.S.; Chen, Y.H. Embryonic exposure to diclofenac disturbs actin organization and leads to myofibril misalignment. Birth Defects Res. B Dev. Reprod. Toxicol. 2011, 92, 139–147. [Google Scholar] [CrossRef]
- Ding, Y.J.; Chen, Y.H. Developmental nephrotoxicity of aristolochic acid in a zebrafish model. Toxicol. Appl. Pharmacol. 2012, 261, 59–65. [Google Scholar] [CrossRef]
- Chen, Y.H.; Tsai, H.J. Treatment with myf5-morpholino results in somite patterning and brain formation defects in zebrafish. Differentiation 2002, 70, 447–456. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the all compounds in this study are available from Prof. Ching-Yuh Chern.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, Y.-T.; Fong, T.-H.; Chen, H.-M.; Chang, C.-Y.; Wang, Y.-H.; Chern, C.-Y.; Chen, Y.-H. Toxicity Assessments of Chalcone and Some Synthetic Chalcone Analogues in a Zebrafish Model. Molecules 2014, 19, 641-650. https://doi.org/10.3390/molecules19010641
Lee Y-T, Fong T-H, Chen H-M, Chang C-Y, Wang Y-H, Chern C-Y, Chen Y-H. Toxicity Assessments of Chalcone and Some Synthetic Chalcone Analogues in a Zebrafish Model. Molecules. 2014; 19(1):641-650. https://doi.org/10.3390/molecules19010641
Chicago/Turabian StyleLee, Ya-Ting, Tsorng-Harn Fong, Hui-Min Chen, Chao-Yuan Chang, Yun-Hsin Wang, Ching-Yuh Chern, and Yau-Hung Chen. 2014. "Toxicity Assessments of Chalcone and Some Synthetic Chalcone Analogues in a Zebrafish Model" Molecules 19, no. 1: 641-650. https://doi.org/10.3390/molecules19010641
APA StyleLee, Y.-T., Fong, T.-H., Chen, H.-M., Chang, C.-Y., Wang, Y.-H., Chern, C.-Y., & Chen, Y.-H. (2014). Toxicity Assessments of Chalcone and Some Synthetic Chalcone Analogues in a Zebrafish Model. Molecules, 19(1), 641-650. https://doi.org/10.3390/molecules19010641