Hormesis of Glyceollin I, an Induced Phytoalexin from Soybean, on Budding Yeast Chronological Lifespan Extension

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
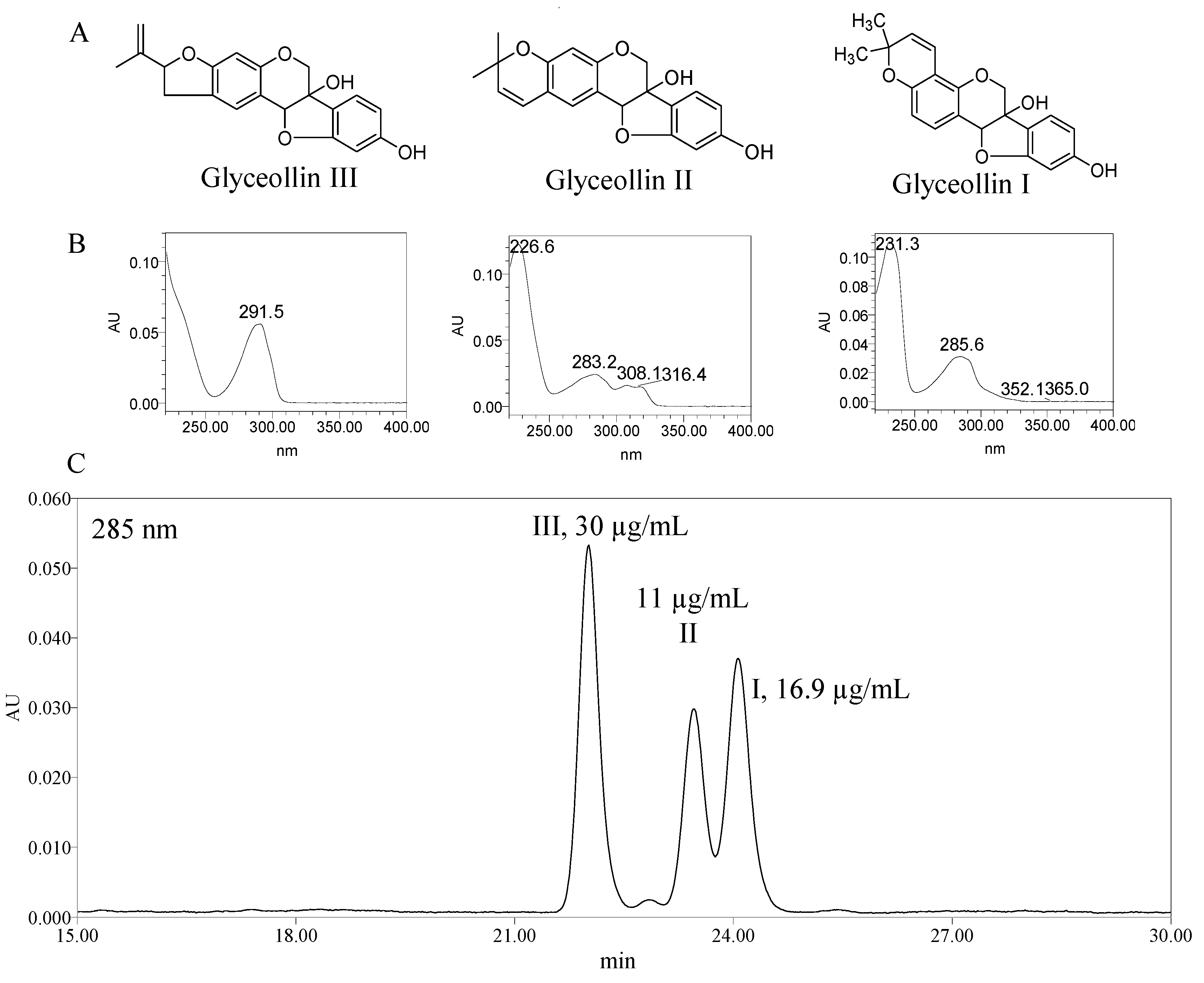
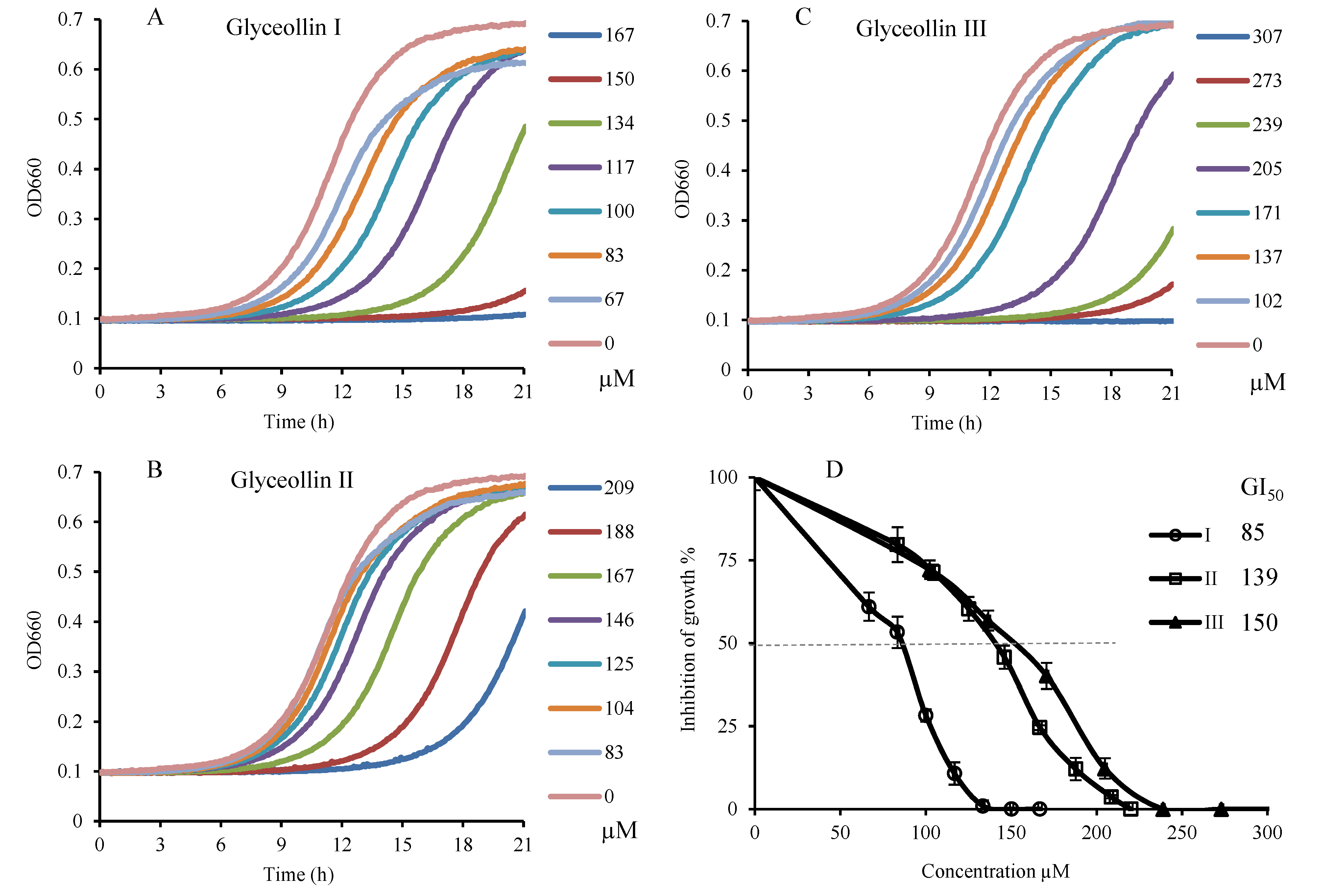
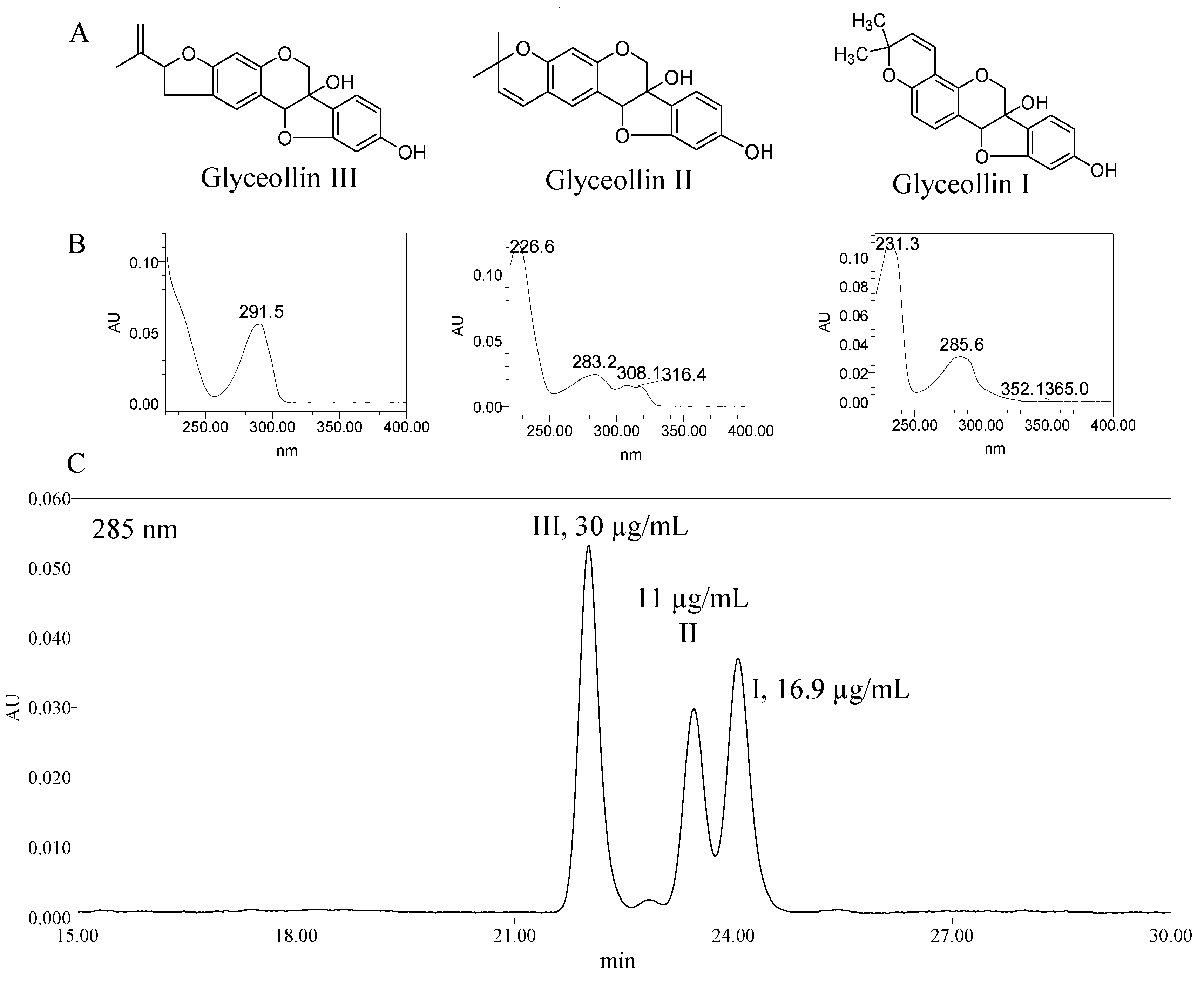
2.1. Antiproliferation Activity of Glyceollins

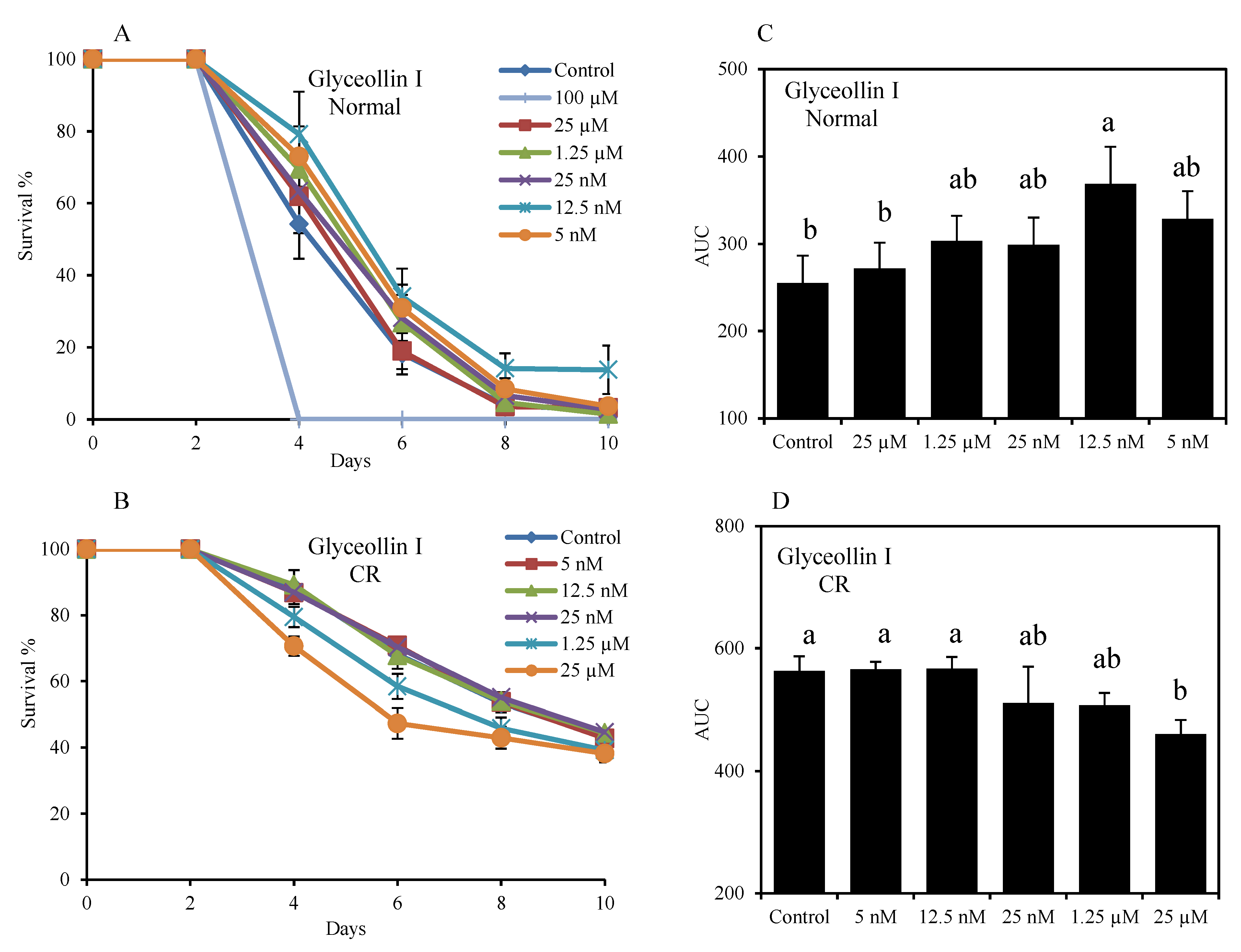
2.2. Glyceollin I Extends Yeast CLS by a CR-Dependent Regime

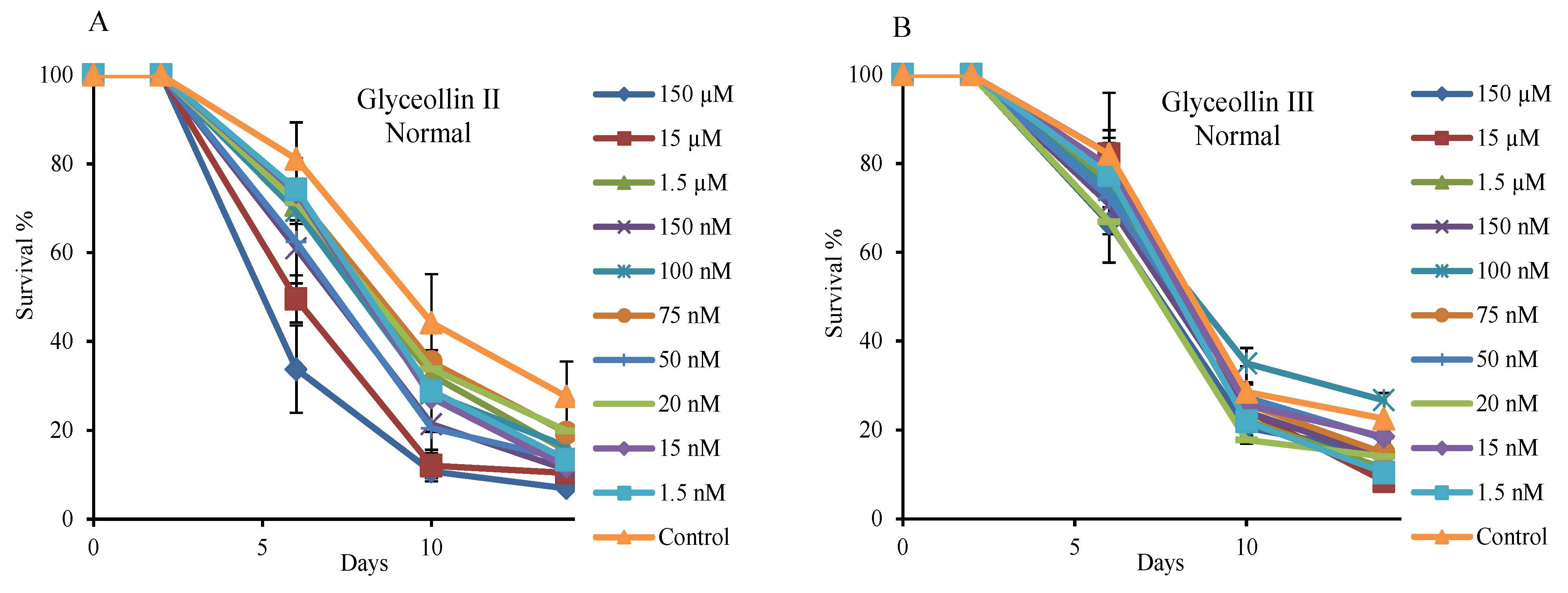
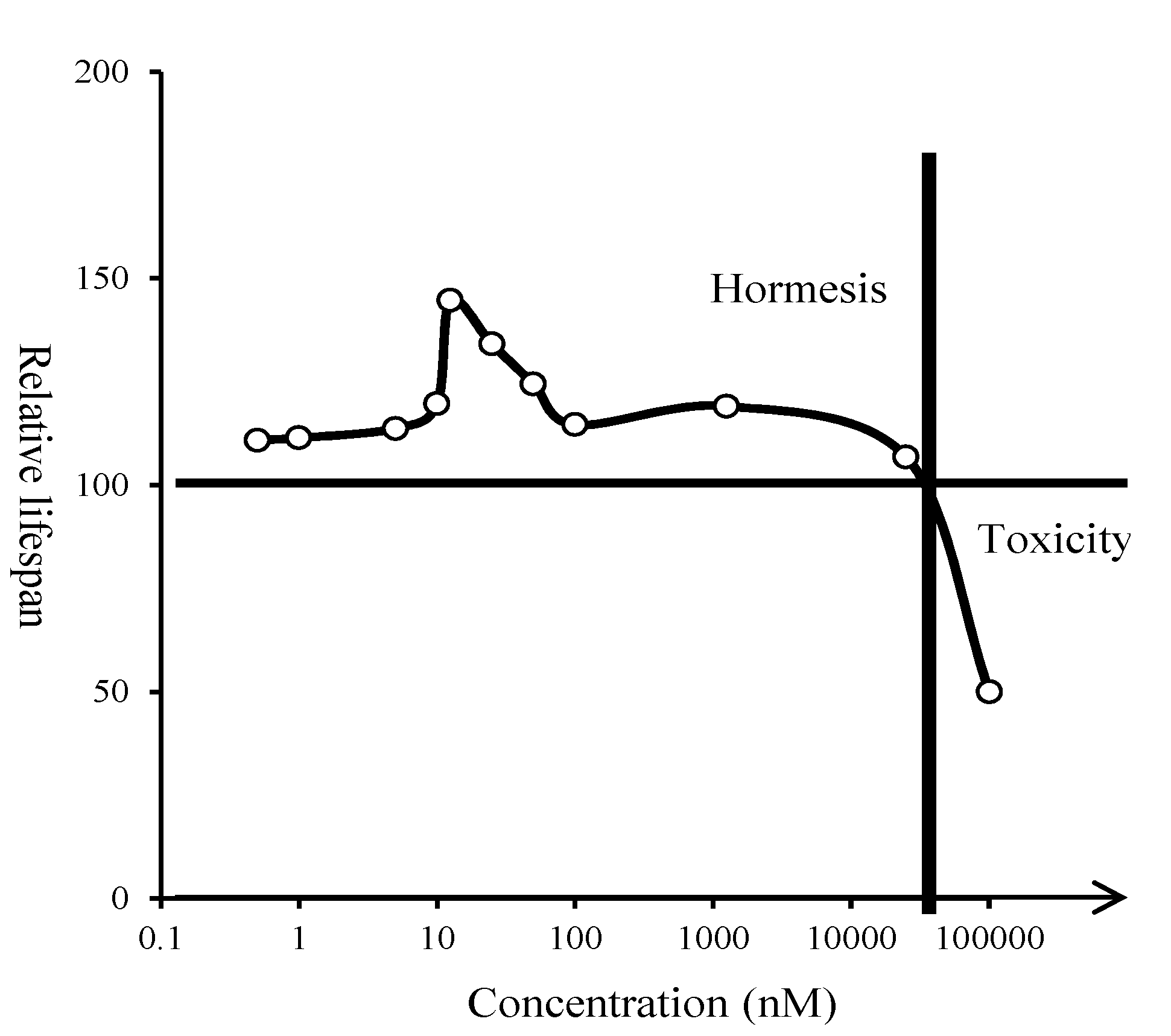
2.3. Hormetic Effect of Glyceollin I on Yeast Life Span


3. Experimental
3.1. Materials
3.2. Isolation of Glyceollins
3.3. Antiproliferation Assay
3.4. Chronological Life Span Assay
3.5. Data Analysis
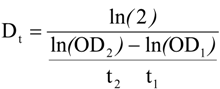
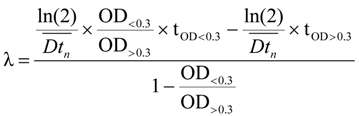
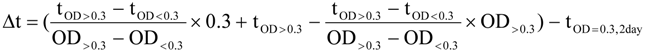
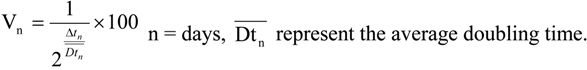
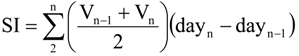
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Francisco, M.L.D.L.; Resurreccion, A.V.A. Functional components in peanuts. Crit. Rev. Food Sci. Nutr. 2008, 48, 715–746. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Feng, S.; Liu, Y.; He, G.; Yioe, Y.; Liu, S.; Huang, D. Germination dramatically increases isoflavonoid content and diversity in chickpea (Cicer arietinum L.) seeds. J. Agric. Food Chem. 2012, 60, 8606–8615. [Google Scholar] [CrossRef]
- Van Loon, L.; Rep, M.; Pieterse, C. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef]
- Hammerschmidt, R. Phytoalexins: What have we learned after 60 years? Annu. Rev. Phytopathol. 1999, 37, 285–306. [Google Scholar] [CrossRef]
- Boue, S.; Cleveland, T.; Carter-Wientjes, C.; Shih, B.; Bhatnagar, D.; McLachlan, J.; Burow, M. Phytoalexin-enriched functional foods. J. Agric. Food Chem. 2009, 57, 2614–2622. [Google Scholar] [CrossRef]
- Feng, S.; Saw, C.; Lee, Y.; Huang, D. Novel process of fermenting black soybean [Glycine max (L.) Merrill] yogurt with dramatically reduced flatulence-causing oligosaccharides but enriched soy phytoalexins. J. Agric. Food Chem. 2008, 56, 10078–10084. [Google Scholar] [CrossRef]
- Feng, S.; Song, L.; Lee, Y.; Huang, D. The effects of fungal stress on the antioxidant contents of black soybeans under Germination. J. Agric. Food Chem. 2010, 58, 12491–12496. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Huang, D. Food grade fungal stress on germinating peanut seeds induced phytoalexins and enhanced polyphenolic antioxidants. J. Agric. Food Chem. 2011, 59, 5993–6003. [Google Scholar] [CrossRef]
- Pervaiz, S.; Holme, A. Resveratrol: Its biologic targets and functional activity. Antioxid. Redox Signal. 2009, 11, 2851–2897. [Google Scholar] [CrossRef]
- McCay, C.M.; Crowell, M.F.; Maynard, L.A. The effect of retarded growth upon the length of life span and upon the ultimate body size: One figure. J. Nutr. 1935, 10, 63. [Google Scholar]
- Mair, W.; Dillin, A. Aging and survival: The genetics of life span extension by dietary restriction. Annu. Rev. Biochem. 2008, 77, 727–754. [Google Scholar] [CrossRef]
- Baur, J.A.; Sinclair, D.A. Therapeutic potential of resveratrol: The in vivo evidence. Nat. Rev. Drug Discov. 2006, 5, 493–506. [Google Scholar] [CrossRef]
- Howitz, K.T.; Bitterman, K.J.; Cohen, H.Y.; Lamming, D.W.; Lavu, S.; Wood, J.G.; Zipkin, R.E.; Chung, P.; Kisielewski, A.; Zhang, L.L.; et al. Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 2003, 425, 191–196. [Google Scholar] [CrossRef]
- Harrison, D.E.; Strong, R.; Sharp, Z.D.; Nelson, J.F.; Astle, C.M.; Flurkey, K.; Nadon, N.L.; Wilkinson, J.E.; Frenkel, K.; Carter, C.S.; et al. Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 2009, 460, 392–395. [Google Scholar]
- Steinkraus, K.A.; Kaeberlein, M.; Kennedy, B.K. Replicative aging in yeast: The means to the end. Annu. Rev. Cell Dev. Biol. 2008, 24, 29–54. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar] [CrossRef]
- Mattson, M. Hormesis defined. Age Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef]
- Saul, N.; Pietsch, K.; Stürzenbaumb, S.R.; Menzel, R.; Steinberg, C.E.W. Hormesis and longevity with tannins: Free of charge or cost-intensive? Chemosphere 2013, 93, 1005–1008. [Google Scholar] [CrossRef]
- Howitz, K.; Sinclair, D. Xenohormesis: Sensing the chemical cues of other species. Cell 2008, 133, 387–391. [Google Scholar] [CrossRef]
- Gems, D.; Partridge, L. Stress-response hormesis and aging. Cell Metab. 2008, 7, 200–203. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. A high throughput screening assay for determination of chronological lifespan of yeast. Exp. Gerontol. 2011, 46, 915–922. [Google Scholar] [CrossRef]
- Feng, S.; Saw, C.L.; Lee, Y.K.; Huang, D. Fungal-stressed germination of black soybeans leads to generation of oxooctadecadienoic acids in addition to glyceollins. J. Agric. Food Chem. 2007, 55, 8589–8595. [Google Scholar] [CrossRef]
- Ng, T.B.; Ye, X.J.; Wong, J.H.; Fang, E.F.; Chan, Y.S.; Pan, W.; Ye, X.Y.; Sze, S.C.; Zhang, K.Y.; Liu, F.; et al. Glyceollin, a soybean phytoalexin with medicinal properties. Appl. Microbiol. Biotechnol. 2011, 90, 59–68. [Google Scholar] [CrossRef]
- Khupse, R.S.; Sarver, J.G.; Trendel, J.A.; Bearss, N.R.; Reese, M.D.; Wiese, T.E.; Boue, S.M.; Burow, M.E.; Cleveland, T.E.; Bhatnagar, D.; et al. Biomimetic syntheses and antiproliferative activities of racemic, natural (−), and unnnatural (+) glyceollin I. J. Med. Chem. 2011, 54, 3506–3523. [Google Scholar] [CrossRef]
- Zimmermann, M.C.; Tilghman, S.L.; Boue, S.M.; Salvo, V.A.; Elliott, S.; Williams, K.Y.; Skripnikova, E.V.; Ashe, H.; Payton-Stewart, F.; Vanhoy-Rhodes, L.; et al. Glyceollin I, a novel antiestrogenic phytoalexin isolated from activated soy. J. Pharmacol. Exp. Ther. 2010, 332, 35–45. [Google Scholar] [CrossRef]
- Payton-Stewart, F.; Khupse, R.S.; Boue, S.M.; Elliott, S.; Zimmermann, M.C.; Skripnikova, E.V.; Ashe, H.; Tilghman, S.L.; Beckman, B.S.; Cleveland, T.E.; et al. Glyceollin I enantiomers distinctly regulate ER-mediated gene expression. Steroids 2010, 75, 870–878. [Google Scholar] [CrossRef]
- Kim, H.J.; Cha, B.Y.; Choi, B.; Lim, J.S.; Woo, J.T.; Kim, J.S. Glyceollins inhibit platelet-derived growth factor-mediated human arterial smooth muscle cell proliferation and migration. Br. J. Nutr. 2011, 107, 1–12. [Google Scholar]
- Payton-Stewart, F.; Schoene, N.W.; Kim, Y.S.; Burow, M.E.; Cleveland, T.E.; Boue, S.M.; Wang, T.T. Molecular effects of soy phytoalexin glyceollins in human prostate cancer cells LNCaP. Mol. Carcinog. 2009, 48, 862–871. [Google Scholar] [CrossRef]
- Simon, J.A.; Bedalov, A. Opinion—Yeast as a model system for anticancer drug discovery. Nat. Rev. Cancer 2004, 4, 481–488. [Google Scholar] [CrossRef]
- Kaeberlein, M. Resveratrol and rapamycin: Are they anti-aging drugs? Bioessays 2010, 32, 96–99. [Google Scholar] [CrossRef]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Buttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Ingram, D.K.; Zhu, M.; Mamczarz, J.; Zou, S.; Lane, M.A.; Roth, G.S.; deCabo, R. Calorie restriction mimetics: An emerging research field. Aging Cell 2006, 5, 97–108. [Google Scholar] [CrossRef]
- Goldberg, A.A.; Richard, V.R.; Kyryakov, P.; Bourque, S.D.; Beach, A.; Burstein, M.T.; Glebov, A.; Koupaki, O.; Boukh-Viner, T.; Gregg, C.; et al. Chemical genetic screen identifies lithocholic acid as an anti-aging compound that extends yeast chronological life span in a TOR-independent manner, by modulating housekeeping longevity assurance processes. Aging (Albany NY) 2010, 2, 393–414. [Google Scholar]
- Goldberg, A.A.; Kyryakov, P.; Bourque, S.D.; Titorenko, V.I. Xenohormetic, hormetic and cytostatic selective forces driving longevity at the ecosystemic level. Aging (Albany NY) 2010, 2, 361–370. [Google Scholar]
- Calabrese, E.J.; Baldwin, L.A. Hormesis: The dose-response revolution. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 175–197. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. Independent and additive effects of glutamic acid and methionine on yeast longevity. PLoS One 2013, 8, e79319. [Google Scholar]
- Wu, Z.; Liu, S.Q.; Huang, D. Dietary restriction depends on nutrient composition to extend chronological lifespan in budding yeast Saccharomyces cerevisiae. PLoS One 2013, 8, e64448. [Google Scholar]
- Toussaint, M.; Conconi, A. High-throughput and sensitive assay tomeasure yeast cell growth: A bench protocol for testing genotoxic agents. Nat. Protoc. 2006, 1, 1922–1928. [Google Scholar] [CrossRef]
- Murakami, C.J.; Burtner, C.R.; Kennedy, B.K.; Kaeberlein, M. A method for high throughput quantitative analysis of yeast chronological life span. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 113–121. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, Y.; Wu, Z.; Feng, S.; Yang, X.; Huang, D. Hormesis of Glyceollin I, an Induced Phytoalexin from Soybean, on Budding Yeast Chronological Lifespan Extension. Molecules 2014, 19, 568-580. https://doi.org/10.3390/molecules19010568
Liu Y, Wu Z, Feng S, Yang X, Huang D. Hormesis of Glyceollin I, an Induced Phytoalexin from Soybean, on Budding Yeast Chronological Lifespan Extension. Molecules. 2014; 19(1):568-580. https://doi.org/10.3390/molecules19010568
Chicago/Turabian StyleLiu, Yuancai, Ziyun Wu, Shengbao Feng, Xuena Yang, and Dejian Huang. 2014. "Hormesis of Glyceollin I, an Induced Phytoalexin from Soybean, on Budding Yeast Chronological Lifespan Extension" Molecules 19, no. 1: 568-580. https://doi.org/10.3390/molecules19010568