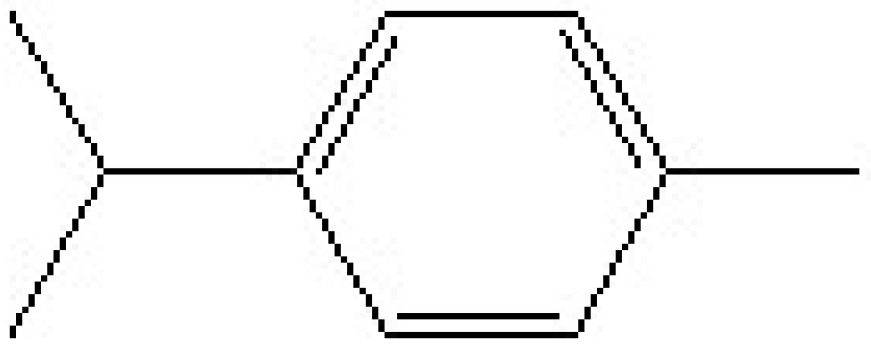
p-Cymene Protects Mice Against Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammatory Cell Activation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
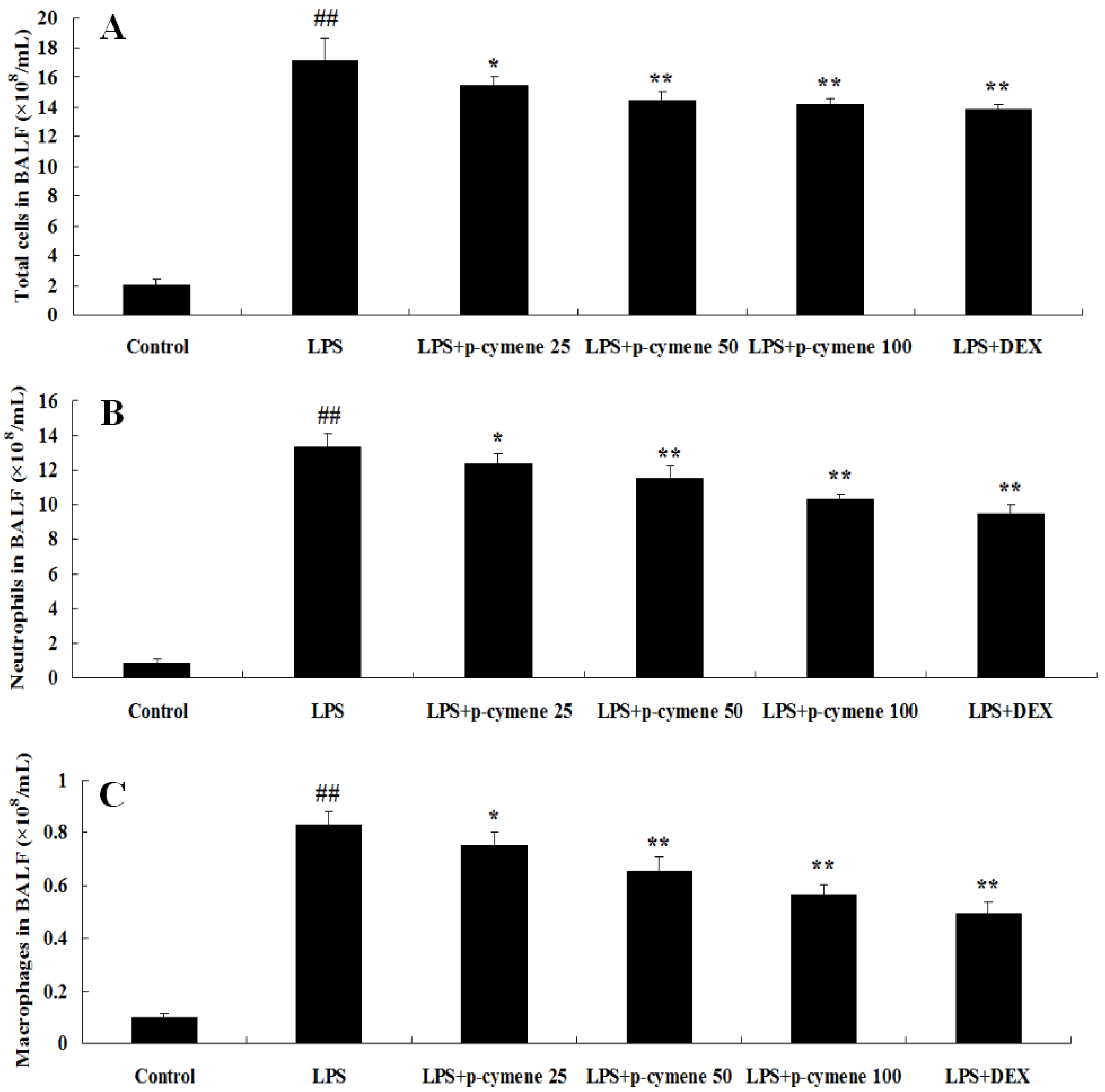
2.1. Effect of p-Cymene on Inflammatory Cell Count in BALF of LPS-Induced Mice

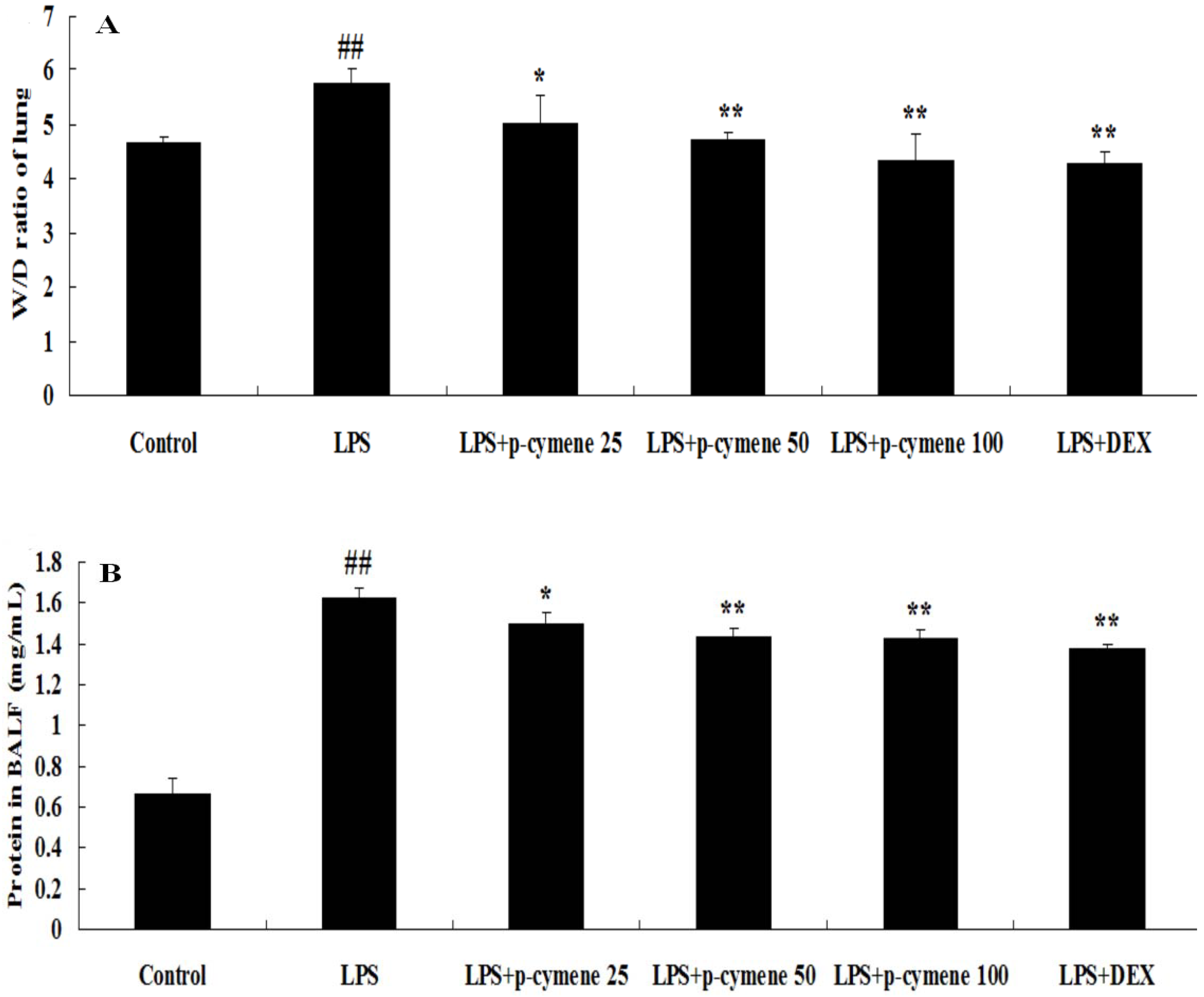
2.2. Effects of p-Cymene on the Lung W/D Ratio and BALF Protein Concentration in LPS-Induced ALI Mice

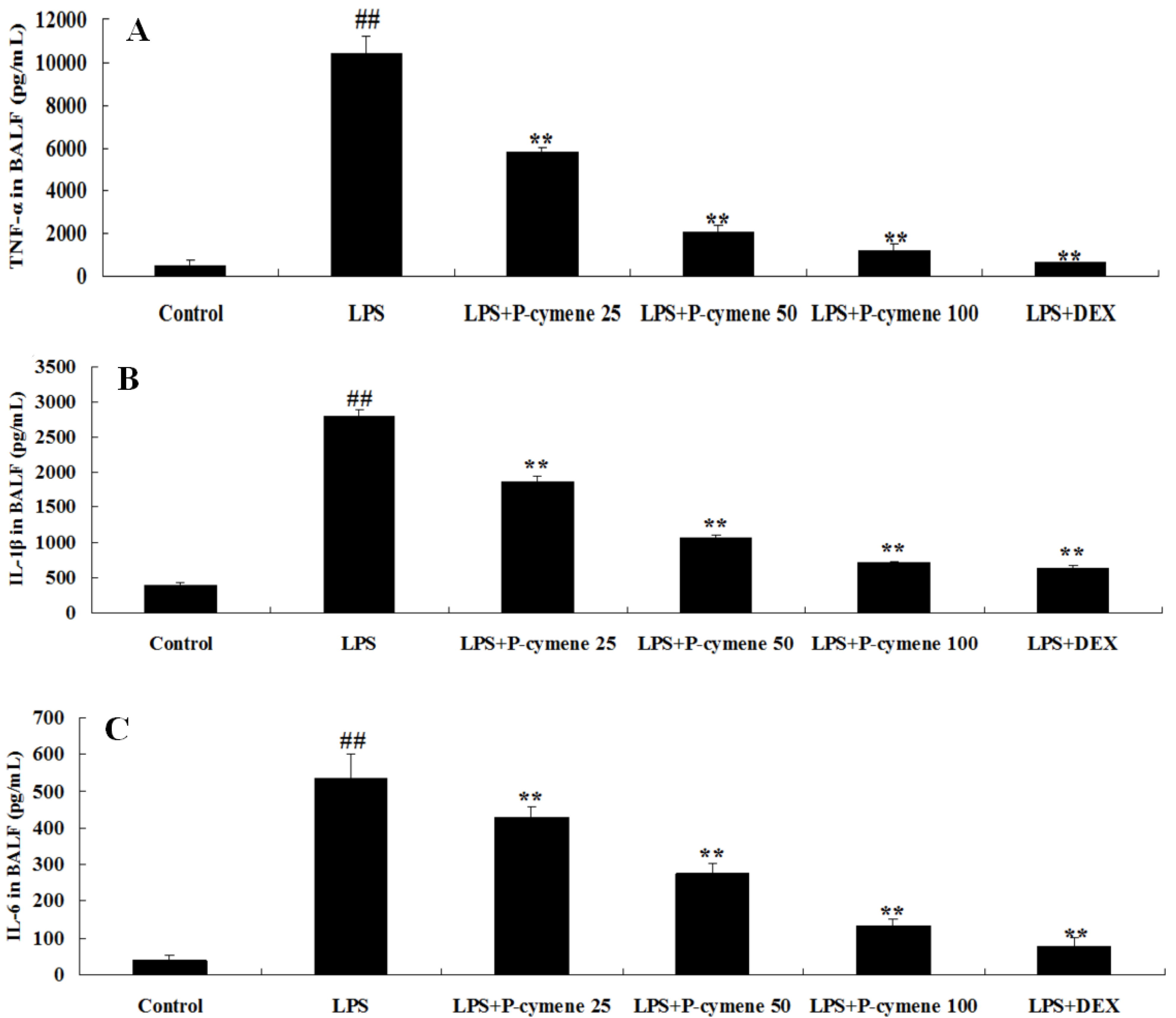
2.3. Effect of p-Cymene on LPS-Induced Release of Cytokines in the BALF of Mice

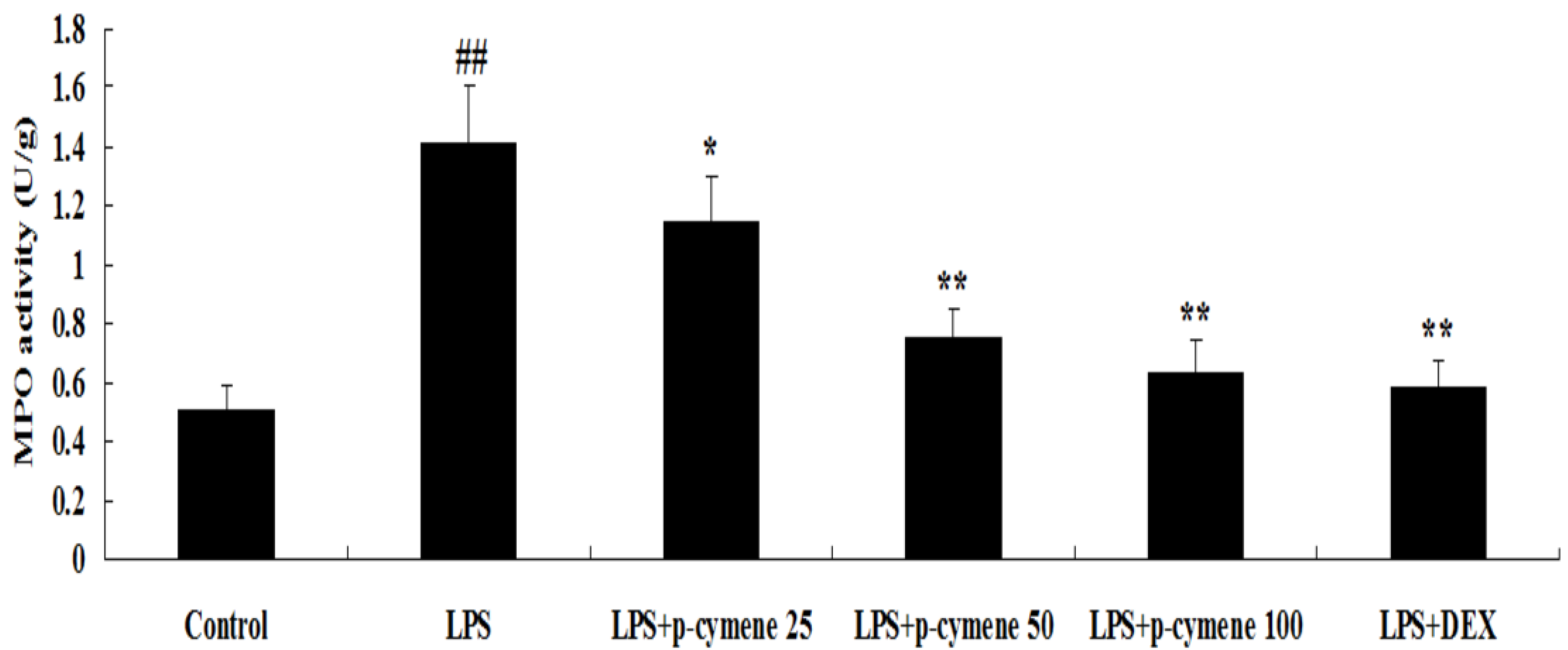
2.4. Effect of p-Cymene on MPO Activity

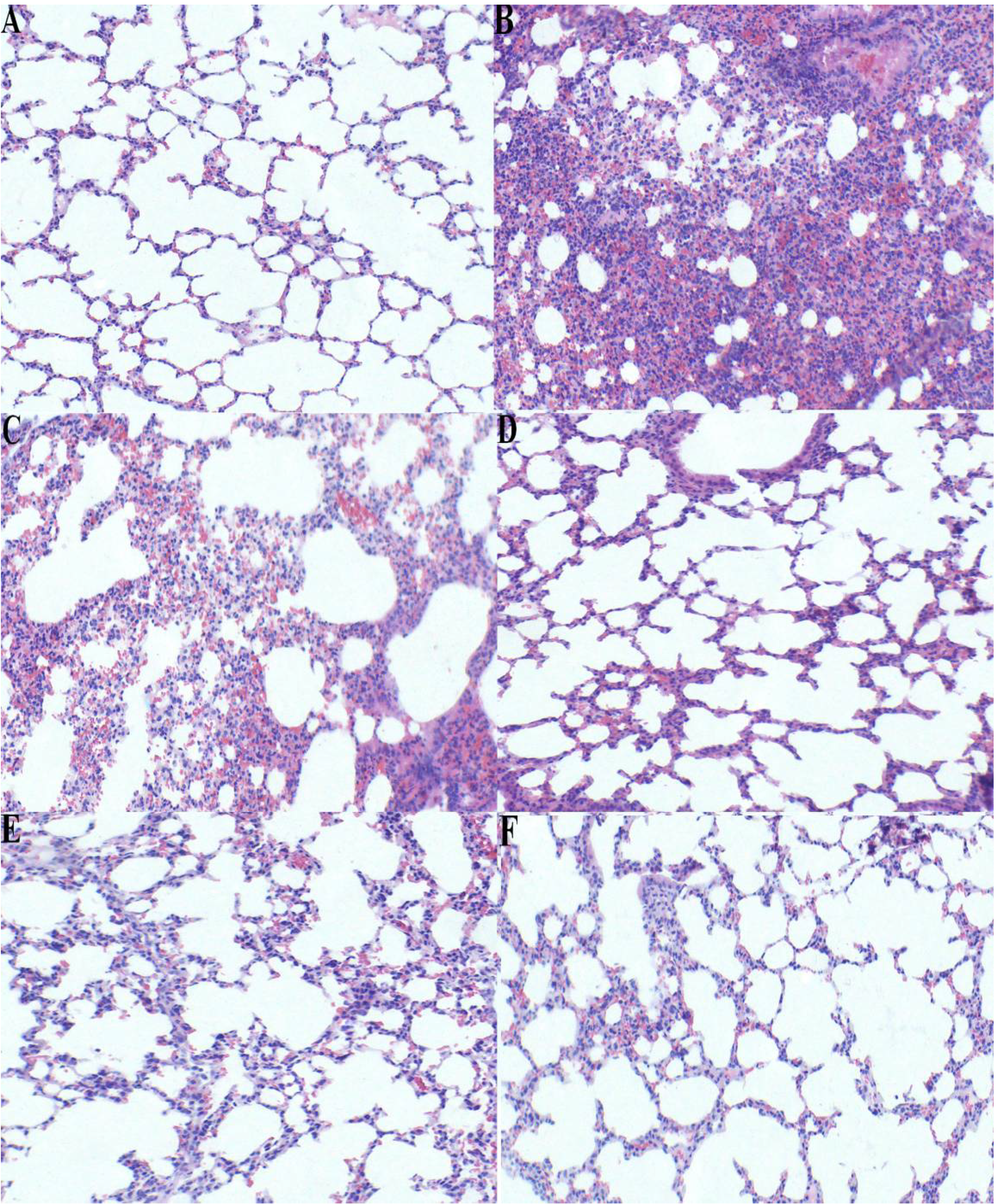
2.5. Effect of p-Cymene on Histopathological Changes in the Lung Tissue of LPS-induced ALI Mice
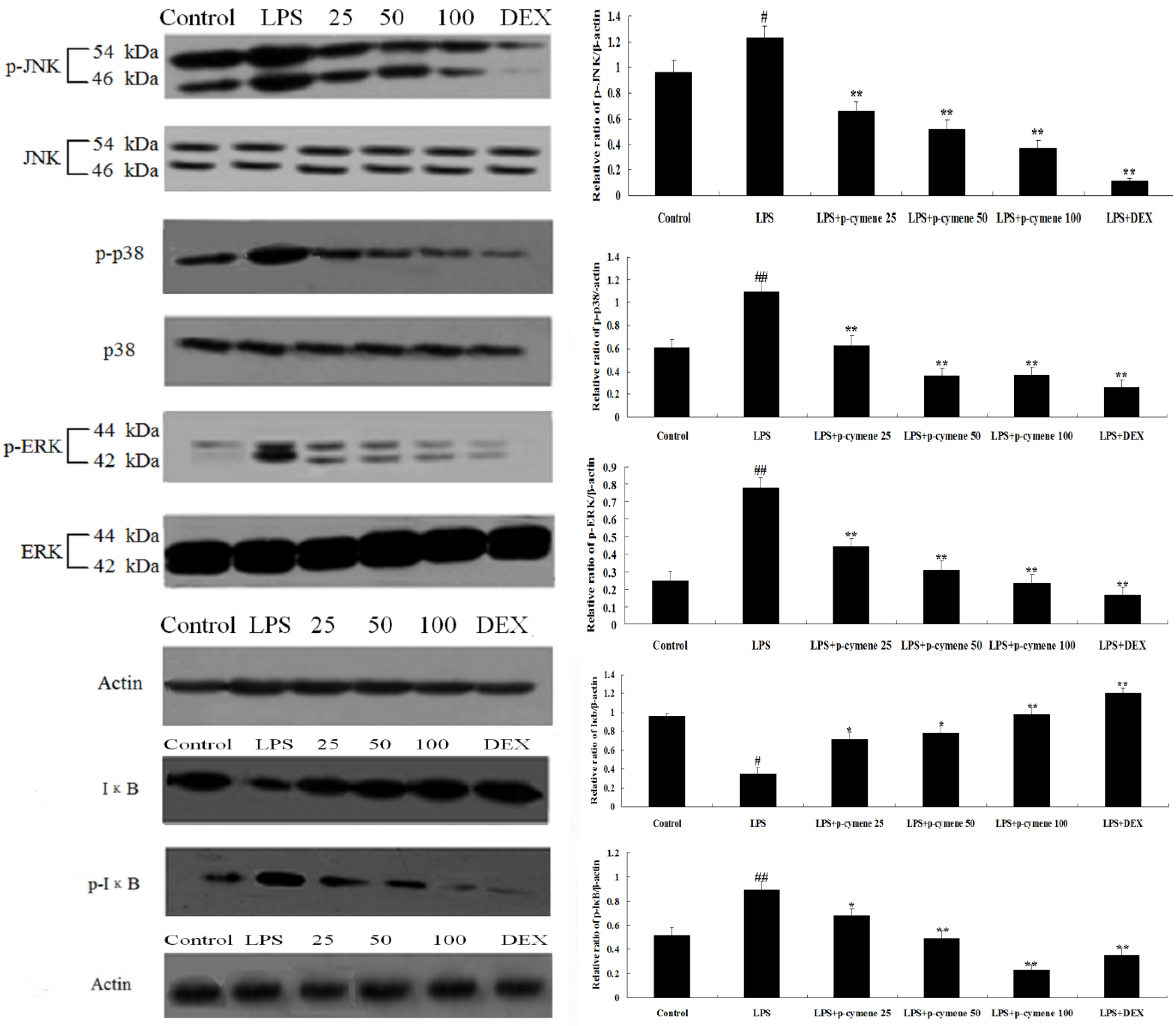
2.6. Effect of p-Cymene on LPS-Induced MAPK and NF-κB Pathways


2.7. Discussion
3. Experimental
3.1. Chemicals and Reagents
3.2. Animals
3.3. LPS-induced ALI Model
3.4. Inflammatory Cell Counts of BALF
3.5. Lung Wet-to-Dry Weight (W/D) Ratio and Protein Analysis
3.6. Cytokine Assays
3.7. Pulmonary Myeloperoxidase Activity in ALI Mice
3.8. Histopathological Evaluation of the Lung Tissue
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Zhang, Q.G.; Bi, L.W.; Zhao, Z.D.; Chen, Y.P.; Li, D.M.; Gu, Y.; Wang, J.; Chen, Y.X.; Bo, C.Y.; Liu, X.Z. Application of ultrasonic spraying in preparation of p-cymene by industrial dipentene dehydrogenation. Chem. Eng. J. 2010, 159, 190–194. [Google Scholar] [CrossRef]
- Benchaar, C.; Calsamiglia, S.; Chaves, A.V.; Fraser, G.R.; Colombatto, D.; McAllister, T.A.; Beauchemin, K.A. A review of plant-derived essential oils in ruminant nutrition and production. Anim. Feed Sci. Technol. 2008, 145, 209–228. [Google Scholar] [CrossRef]
- Singh, H.P.; Kohli, R.K.; Batish, D.R.; Kaushal, P.S. Allelopathy of gymnospermous trees. J. Forest. Res. 1999, 4, 245–254. [Google Scholar] [CrossRef]
- Selvaraj, M.; Pandurangan, A.; Seshadri, K.S.; Sinha, P.K.; Krishnasamy, V.; Lal, K.B. Comparison of mesorporous A1-MCM-41 molecular sieves in the production of p-cymene for isopropylation of toluene. J. Mol. Catal. A-Chem. 2002, 186, 173–186. [Google Scholar] [CrossRef]
- Siani, A.C.; Garrido, I.S.; Carvalho, E.S.; Ramos, M.F.S. Evaluation of anti-inflammatory-related activity of essential oils from the leaves and resin of species of Protium. J. Ethnopharmacol. 1999, 66, 57–69. [Google Scholar] [CrossRef]
- Oliveira, F.A.; Costa, C.L.S.; Chaves, M.H.; Almeida, F.R.C.; Cavalcante, I.J.M.; Lima, A.F.; Lima-Júnior, R.C.P.; Silva, R.M.; Campos, A.R.; Santos, F.A.; et al. Attenuation of capsaicin-induced acute and visceral nociceptive pain by alpha- and beta-amyrin, a triterpene mixture isolated from Protium heptaphyllum resin in mice. Life Sci. 2005, 77, 2942–2952. [Google Scholar] [CrossRef]
- Otuki, M.F.; Lima, F.V.; Malheiros, A.; Filho, V.C.; Monache, F.D.M.; Yunes, R.A.; Calixto, J. Evaluation of the antinociceptive action caused by ether fraction and a triterpene isolated from resin of Protium kleinii. Life Sci. 2001, 69, 2225–2236. [Google Scholar] [CrossRef]
- Bispo, M.D.; Mourão, R.H.V.; Franzotti, E.M.; Bomfim, K.B.R.; Arrigoni-Blank, M.F.; Moreno, M.P.N.; Marchioro, M.; Antoniolli, A.R. Antinociceptive and antiedematogenic effects of the aqueous extract of Hyptis pectinata leaves in experimental animals. J. Etnopharmacol. 2001, 76, 81–86. [Google Scholar] [CrossRef]
- Ramezani, M.; Hosseinzadeh, H.; Samizadeh, S. Antinociceptive effects of Zataria multiflora Boiss fractions in mice. J. Ethonopharmacol. 2004, 91, 167–170. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Synergistic antimicrobial effect of nisin and p-cymene on Salmonella enterica serovar Typhi in vitro and on ready-to-eat food. Biotechnol. Biochem. 2010, 74, 520–524. [Google Scholar] [CrossRef]
- Santana, M.F.; Quintans-Júnior, L.J.; Sócrates Cavalcanti, C.H.; Makson Oliveira, G.B.; Guimarães, A.G.; Cunha, E.S.; Melo, M.S.; Márcio Santos, R.V.; Adriano Araújo, A.S.; Bonjardim, L.R. p-Cymene reduces orofacial nociceptive response in mice. Rev. Bras. Farmacogn. 2011, 21, 1138–1143. [Google Scholar] [CrossRef]
- Peng, X.Q.; Damarla, M.; Skirball, J.; Nonas, S.; Wang, X.Y.; Han, E.J.; Hasan, E.J.; Cao, X.; Boueiz, A.; Damico, R.; et al. Protective role of PI3-kinase/Akt/eNOS signaling in mechanical stress through inhibition of p38 mitogen-activated protein kinase in mouse lung. Acta Pharmacol. Sin. 2010, 31, 175–183. [Google Scholar] [CrossRef]
- The Acute Respiratory Distress Syndrome Network. Ventilation with lower tidal volumes as compared with traditional tidal volumes for acute lung injury and the acute respiratory distress syndrome. N. Engl. J. Med. 2000, 342, 1301–1308. [CrossRef]
- Zhang, X.; Huang, H.; Yang, T.; Ye, Y.; Shan, J.; Yin, Z.; Luo, L. Chlorogenic acid protects mice against lipopolysaccharide-induced acute lung injury. Injury 2010, 41, 746–752. [Google Scholar] [CrossRef]
- Severgnini, M.; Takahashi, S.; Rozo, L.M.; Homer, R.J.; Kuhn, C.; Jhung, J.W.; Perides, G.; Steer, M.; Hassoun, P.M.; Fanburg, B.L.; et al. Activation of the STATpathway in acute lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2004, 286, 1282–1292. [Google Scholar] [CrossRef]
- Windsor, A.C.; Mullen, P.G.; Fowler, A.A. Acute lung injury: What have we learned from animal models? Am. J. Med. Sci. 1993, 306, 111–116. [Google Scholar] [CrossRef]
- Sutcliffe, A.J. The future of ARDS. Injury 1994, 25, 587–593. [Google Scholar] [CrossRef]
- Stockley, R.A. The role of proteinases in the pathogenesis of chronic bronchitis. Am. J. Respir. Crit. Care Med. 1994, 150, 109–113. [Google Scholar]
- Venaille, T.J.; Ryan, G.; Robinson, B.W. Epithelial cell damage is induced by neutrophil-derived, not pseudomonas-derived, proteases in cystic fibrosis sputum. Respir. Med. 1998, 92, 233–240. [Google Scholar] [CrossRef]
- Chu, P.Y.; Chien, S.P.; Hsu, D.Z.; Liu, M.Y. Protective effect of sesamol on the pulmonary inflammatory response and lung injury in endotoxemic rats. Food Chem. Toxicol. 2010, 48, 1821–1826. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, H.; Yang, T.; Ye, Y.; Shan, J.; Yin, Z.; Luo, L. Chlorogenic acid protects mice against lipopolysaccharide-induced acute lung injury. Injury 2010, 41, 746–752. [Google Scholar] [CrossRef]
- Brigham, K.L. Mechanisms of lung injury. Clin. Chest Med. 1982, 3, 9–24. [Google Scholar]
- Sibille, Y.; Reynolds, H.Y. Macrophages and polymorphonuclear neutrophils in lung defence and injury. Am. Rev. Respir. Dis. 1990, 141, 471–501. [Google Scholar]
- Bradley, P.P.; Priebat, D.A.; Christensen, R.D.; Rothestein, G. Measurement of cutaneous inflammation: Estimation of neutrophil content with an enzyme marker. J. Invest. Dermatol. 1982, 78, 206–209. [Google Scholar] [CrossRef]
- Maniatis, N.A.; Kotanidou, A.; Catravas, J.D.; Orfanos, S.E. Pathomechanisms in acute lung injury. Vascul. Pharmacol. 2008, 49, 119–133. [Google Scholar] [CrossRef]
- Goodman, R.B.; Strieter, R.M.; Martin, D.P.; Steinberg, K.P.; Milberg, J.A.; Maunder, R.J.; Kunkel, S.L.; Walz, A.; Hudson, L.D.; Martin, T.R. Inflammatory cytokines in patients with persistence of the acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 1996, 154, 602–611. [Google Scholar]
- Zhu, Y.B.; Wang, Q.; Liu, Y.L.; Li, X.F.; Li, J.A.; Lü, X.D.; Ling, F.; Liu, A.J.; Fan, X.M. Effect of partial liquid ventilation on oleic acid-induced inflammatory responses in piglets. Chin. Med. J. 2010, 123, 2088–2093. [Google Scholar]
- Abraham, E. NF-kappaB activation. Crit. Care Med. 2000, 28, 100–104. [Google Scholar] [CrossRef]
- Blackwell, T.S.; Christman, J.W. The role of nuclear factor-kappa B in cytokine gene regulation. Am. J. Respir. Cell Mol. Biol. 1997, 17, 3–9. [Google Scholar]
- Molnar, A.; Theodoras, A.M.; Zon, L.I.; Kyriakis, J.M. Cdc42Hs, but not Rac 1, inhibits serum-stimulated cell cycle progressin at G1/S through a mechanism requiring p38/RK. J. Biol. Chem. 1997, 272, 13229–13235. [Google Scholar]
- Takenaka, K.; Mcriguchi, T.; Nishida, E. Activation of the protein kinase p38 in the spindle assembly checkpoint and mitotic arrest. Science 1998, 280, 599–602. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H.Z.; Gong, X.; Luo, F.L.; Wang, B.; Hu, N.; Wang, C.D.; Zhang, Z.; Wan, J.Y. Protective effects of Asiaticoside on acute liver injury induced by lipopolysaccharide/D-galactosamine in mice. Phytomedicine 2010, 17, 811–819. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, H.J.; Moon, C.S.; Chong, Y.H.; Kang, J.L. Inhibition of c-Jun NH2-terminal kinase or extracellular signal-regulated kinase improves lung injury. Respir. Res. 2004, 5. [Google Scholar]
- Van den Blink, B.; Juffermans, N.P.; ten Hove, T.; Schultz, M.J.; van Deventer, S.J.; van der Poll, T.; Peppelenbosch, M.P. p38 Mitogen-activated protein kinase inhibition increases cytokine release by macrophages in vitro and during infection in vivo. J. Immunol. 2001, 166, 582–587. [Google Scholar]
- Scherle, P.A.; Jones, E.A.; Favata, M.F.; Daulerio, A.J.; Covington, M.B.; Nurnberg, S.A.; Magolda, R.L.; Trzaskos, J.M. Inhibition of MAP kinase prevents cytokine and prostaglandin E2 production in lipopolysaccharide-stimulated monocytes. J. Immunol. 1998, 161, 5681–5686. [Google Scholar]
- Ysebaert, D.K.; de Greef, K.E.; Vercauteren, S.R.; Ghielli, M.; Verpooten, G.A.; Eyskens, E.J. Identification and kinetics of leukocytes after severe ischaemia/reperfusion renal injury. Nephrol. Dial. Transplant. 2000, 15, 1562–1574. [Google Scholar] [CrossRef]
- Peng, X.; Hassoun, P.M.; Sammani, S.; McVerry, B.J.; Burne, M.J.; Rabb, H. Protective effects of sphingosine1-phosphate in murine endotoxin-induced inflammatory lung injury. Am. J. Respir. Crit. Care Med. 2004, 169, 1245–1251. [Google Scholar] [CrossRef]
- Sample Availability: Samples of p-cymene are available from the authors.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xie, G.; Chen, N.; Soromou, L.W.; Liu, F.; Xiong, Y.; Wu, Q.; Li, H.; Feng, H.; Liu, G. p-Cymene Protects Mice Against Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammatory Cell Activation. Molecules 2012, 17, 8159-8173. https://doi.org/10.3390/molecules17078159
Xie G, Chen N, Soromou LW, Liu F, Xiong Y, Wu Q, Li H, Feng H, Liu G. p-Cymene Protects Mice Against Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammatory Cell Activation. Molecules. 2012; 17(7):8159-8173. https://doi.org/10.3390/molecules17078159
Chicago/Turabian StyleXie, Guanghong, Na Chen, Lanan Wassy Soromou, Fang Liu, Ying Xiong, Qianchao Wu, Hongyu Li, Haihua Feng, and Guowen Liu. 2012. "p-Cymene Protects Mice Against Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting Inflammatory Cell Activation" Molecules 17, no. 7: 8159-8173. https://doi.org/10.3390/molecules17078159