Restoration of Electric Footshock-Induced Immunosuppression in Mice by Gynostemma pentaphyllum Components


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
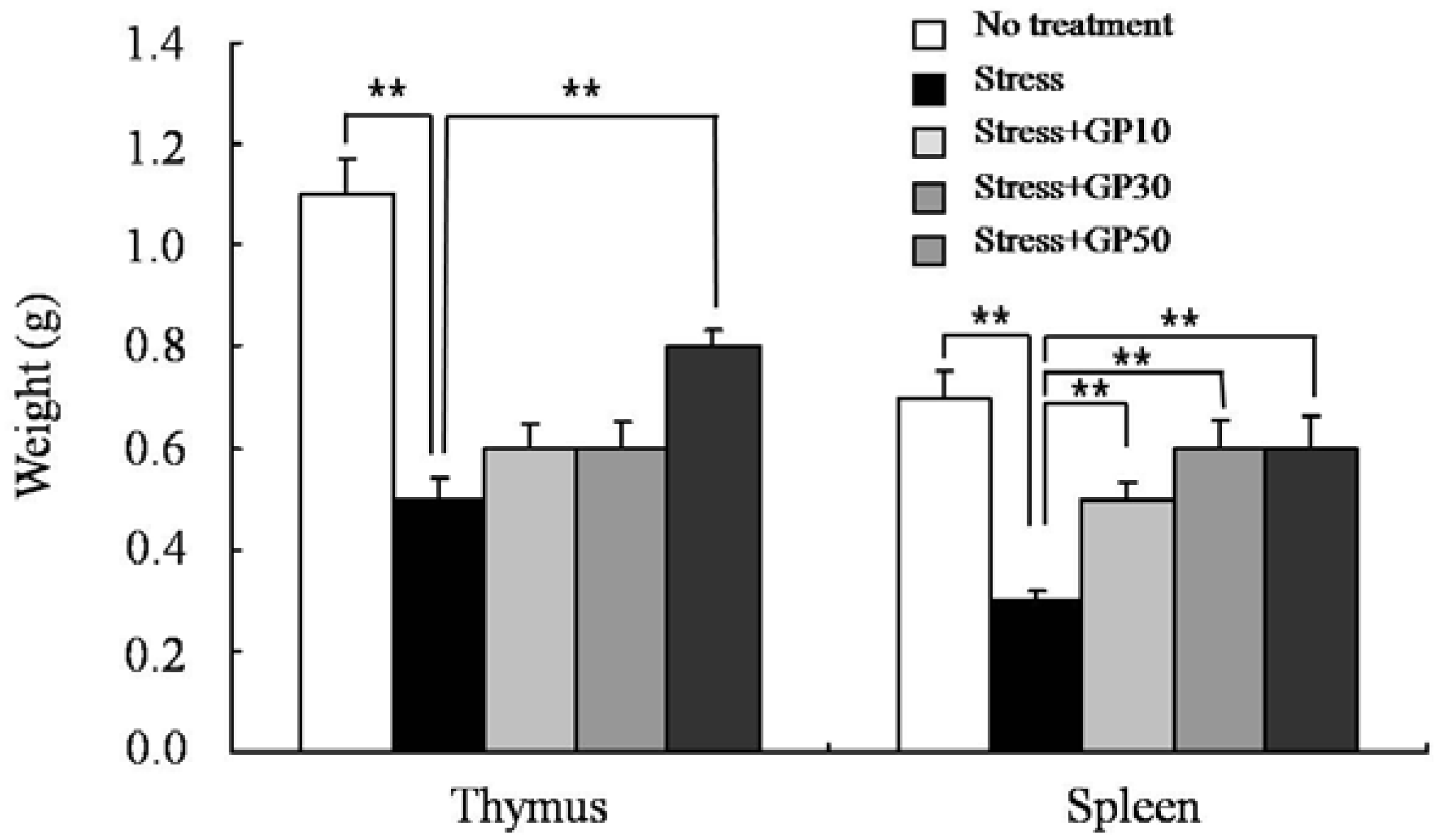
2.1. GP-EX Prevents EFS Stress-Induced Atrophy of Lymphoid Organs

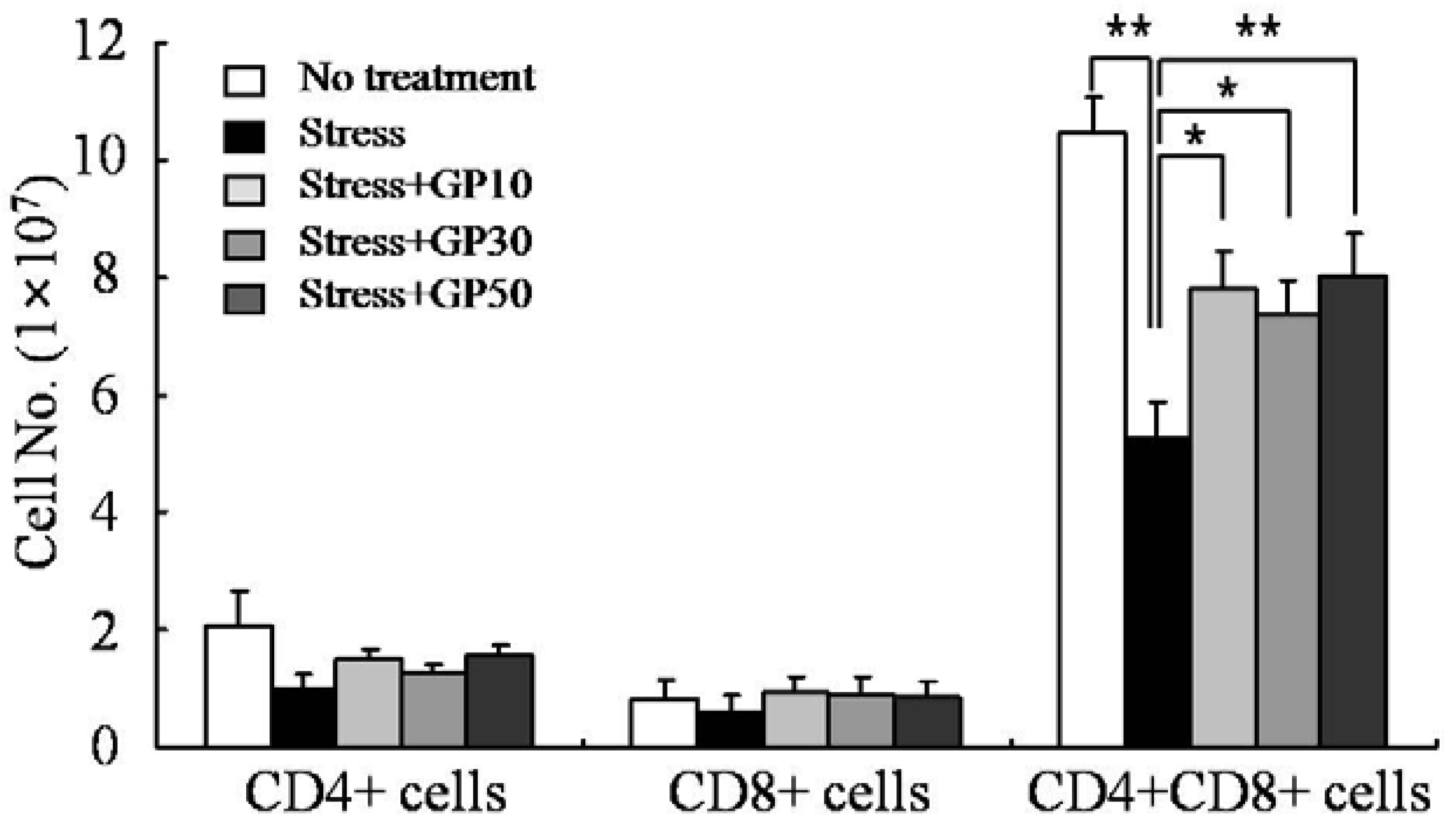
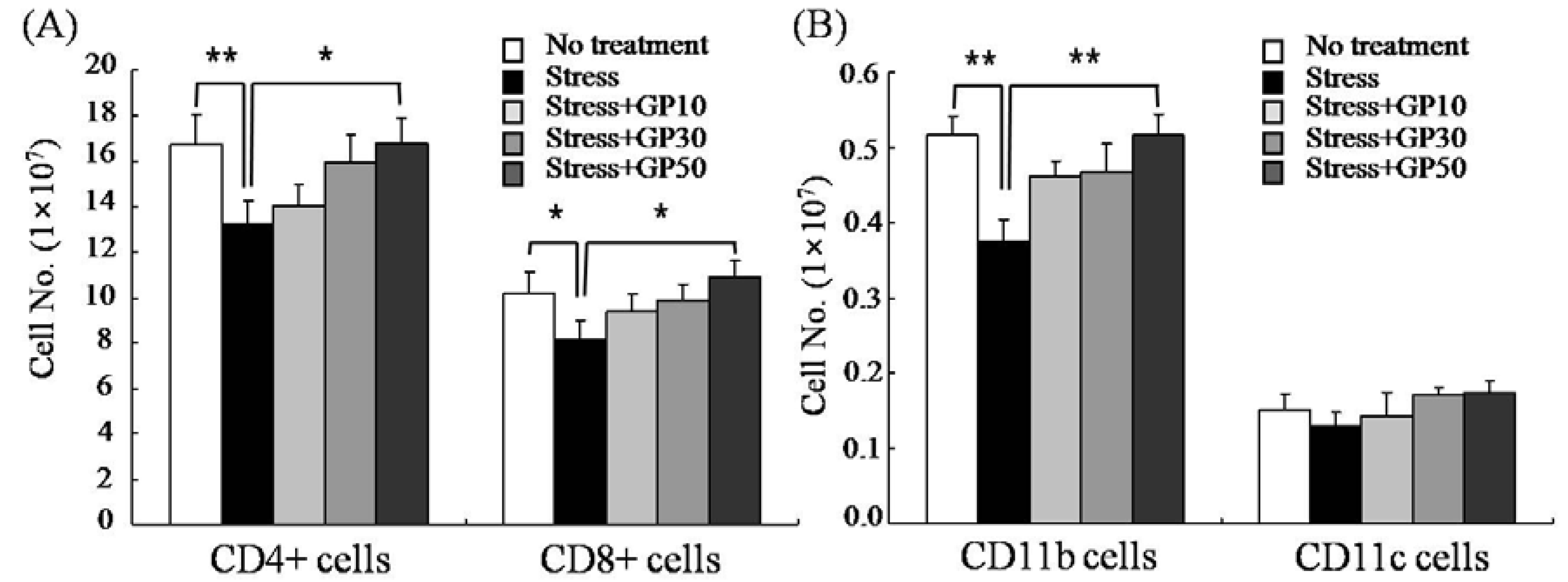
2.2. GP-EX Restores EFS Stress-Induced Disturbances in Lymphocyte Cellularity


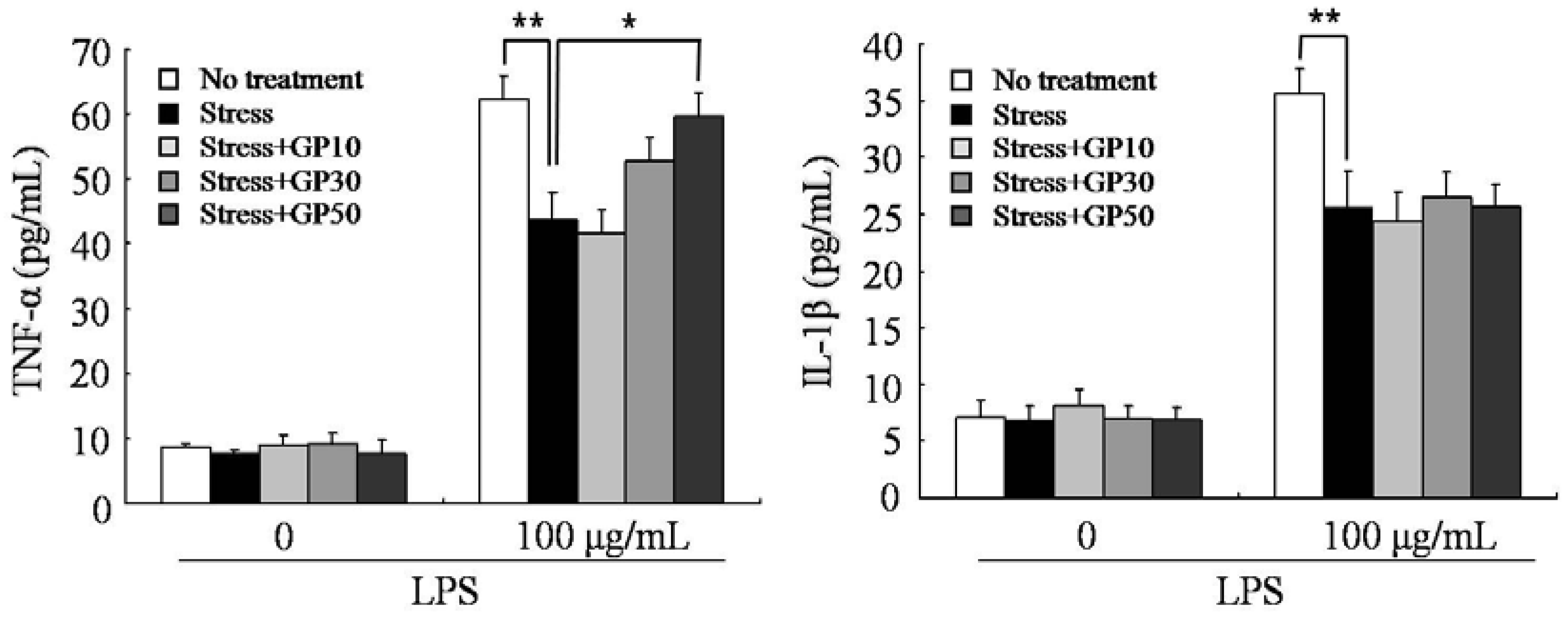
2.3. GP-EX Restores EFS Stress-Induced Decreases in LPS-Induced Cytokine Production

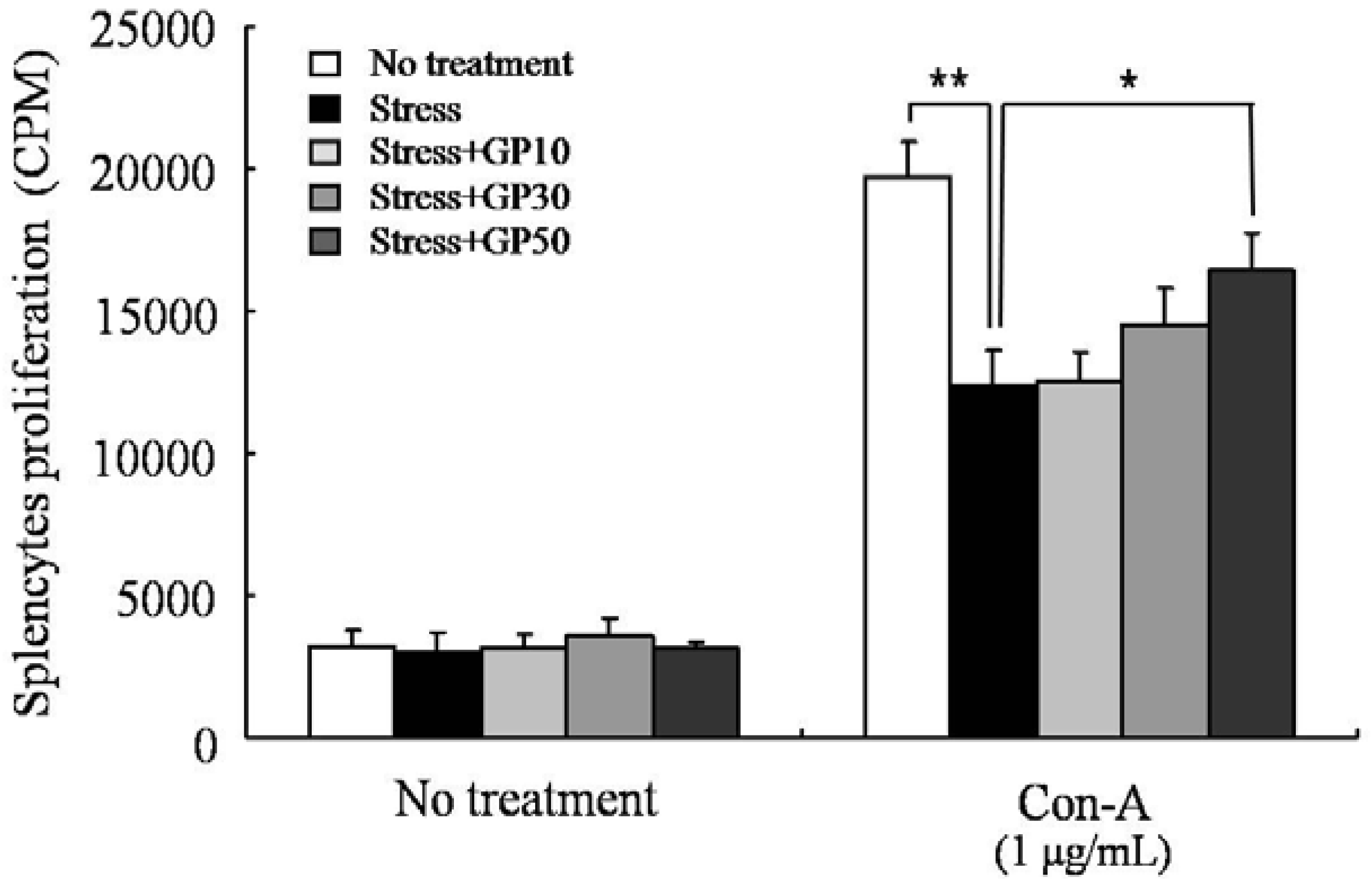
2.4. GP-EX Restores EFS Stress-Induced Deceases in Splenocyte Proliferation
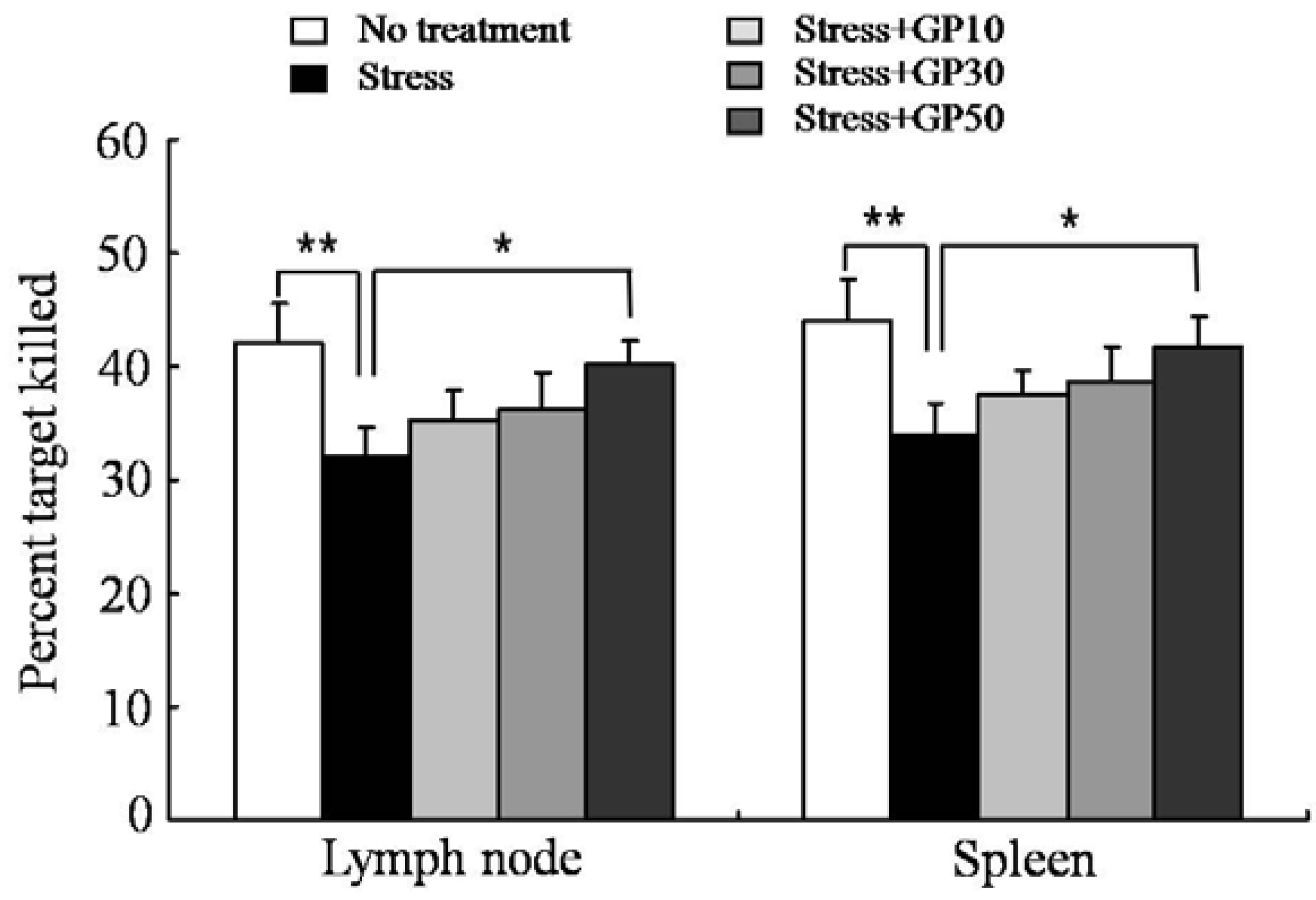
2.5. GP-EX Restores EFS Stress-Induced Deceases in OVA-Specific CTL Activity


3. Discussion
4. Experimental
4.1. Preparation and Treatment of GP-EX
4.2. Animals
4.3. Cell Culture
4.4. EFS Stressor
4.5. Flow Cytometry
4.6. Growth Stimulatory Activity
4.7. Peripheral Blood Cytokine Production
4.7. In Vivo CTL Assay
4.8. Statistical Analysis
5. Conclusions
Acknowledgements
References
- Blumert, M.; Liu, J.L. Jiaogulan, China’s “Immortality” Herb; Torchlight Publishing Inc.: Badger, CA, USA, 1999. [Google Scholar]
- Razmovski-Naumovski, V.; Huang, T.-H.; Tran, V.H.; Li, G.Q.; Duke, C.C.; Roufogalis, B.D. Chemistry and pharmacology of Gynostemma pentaphyllum. Phytochem. Rev. 2005, 4, 197–219. [Google Scholar] [CrossRef]
- Choi, H.S.; Park, M.S.; Kim, S.H.; Hwang, B.Y.; Lee, C.K.; Lee, M.K. Neuroprotective Effects of Herbal Ethanol Extracts from Gynostemma pentaphyllum in the 6-Hydroxydopamine-Lesioned Rat Model of Parkinson’s Disease. Molecules 2010, 15, 2814–2824. [Google Scholar] [CrossRef]
- Li, L.; Jiao, L.; Lau, B.H. Protective effect of gypenosides against oxidative stress in phagocytes, vascular endothelial cells and liver microsomes. Cancer Biother. 1993, 8, 263–272. [Google Scholar] [CrossRef]
- Huang, T.H.; Tran, V.H.; Roufogalis, B.D.; Li, Y. Gypenoside XLIX, a naturally occurring PPAR-alpha activator, inhibits cytokine-induced vascular cell adhesion molecule-1 expression and activity in human endothelial cells. Eur. J. Pharmacol. 2007, 565, 158–165. [Google Scholar] [CrossRef]
- Circosta, C.; De Pasquale, R.; Occhiuto, F. Cardiovascular effects of the aqueous extract of Gynostemma pentaphyllum Makino. Phytomedicine 2005, 12, 638–643. [Google Scholar] [CrossRef]
- Megallia, S.; Aktanb, F.; Daviesc, N.M.; Roufogalisa, B.D. Phytopreventative anti-hyperlipidemic effects of Gynostemma Pentaphyllum in rats. J. Pharm. Pharmaceut. Sci. 2005, 8, 507–515. [Google Scholar]
- Hou, J.; Liu, S.; Ma, Z.; Lang, X.; Wang, J.; Wang, J.; Liang, Z. Effects of Gynostemma pentaphyllum Makino on the immunological functions of cancer patients. J. Trad. Chin. Med. 1991, 11, 47–52. [Google Scholar]
- Norberg, A.; Hoa, N.K.; Liepinsh, E.; Van Phan, D.; Thuan, N.D.; Jornvall, H.; Sillard, R.; Ostenson, C.G. A novel insulin-releasing substance, phanoside, from the plant Gynostemma pentaphyllu. J. Biol. Chem. 2004, 279, 41361–41367. [Google Scholar]
- Lin, J.M.; Lin, C.C.; Chiu, H.F.; Yang, J.J.; Lee, S.G. Evaluation of the anti-inflammatory and liver-protective effects of Anoectochilus formosanus, Ganoderma lucidum and Gynostemma pentaphyllum in rats. Am. J. Chin. Med. 1993, 21, 59–69. [Google Scholar] [CrossRef]
- Yin, F.; Zhang, Y.N.; Yang, Z.Y.; Hu, L.H. Nine new dammarane saponins from Gynostemma pentaphyllum. Chem. Biodiver. 2006, 3, 771–782. [Google Scholar] [CrossRef]
- Kim, J.H.; Han, Y.N. Dammarane-type saponins from Gynostemma pentaphyllum. Phytochemistry 2011, 72, 1453–1459. [Google Scholar]
- Hu, Y.; Ip, F.C.; Fu, G.; Pang, H.; Ye, W.; Ip, N.Y. Dammarane saponins from Gynostemma pentaphyllum. Phytochemistry 2010, 71, 1149–1157. [Google Scholar]
- Selye, H. A syndrome produced by diverse nocuous agents. Nature 1936, 138, 32–34. [Google Scholar] [CrossRef]
- Sterling, P.; Eyer, J. Allostasis: A new paradigm to explain arousal pathology. In Handbook of Life Stress, Cognition and Health; Fisher, S., Reason, J., Eds.; John Wiley & Sons: New York, NY, USA, 1988; pp. 629–649. [Google Scholar]
- McEwen, B.S.; Seeman, T. Protective and damaging effects of mediators of stress. Elaborating and testing the concepts of allostasis and allostatic load. Ann. NY. Acad. Sci. 1999, 896, 30–47. [Google Scholar] [CrossRef]
- McEwen, B.S.; Stellar, E. Stress and the individual. Mechanisms leading to disease. Arch. Intern. Med. 1993, 153, 2093–2101. [Google Scholar] [CrossRef]
- Kovacs, K.J.; Mikios, I.H.; Bali, B. Psychological and Physical Stressors. In Handbook of Stress and the Brain, Part I; Steckler, T., Kalin, N.H., Reul, J.M.H.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 775–792. [Google Scholar]
- Ulrich-Lai, P.M.; Engeland, W.C. Sympatho-adrenal Activity and Hypothalamic-Pituitary-Adrenal Axis Regulation. In Handbook of Stress and the Brain, Part I; Steckler, T., Kalin, N.H., Reul, J.M.H.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 419–436. [Google Scholar]
- Frank, M.G.; Watkins, L.R.; Maier, S.F. Stress- and glucocorticoid-induced priming of neuroinflammatory responses: Potential mechanisms of stress-induced vulnerability to drugs of abuse. Brain Behav. Immun. 2011, 25, S21–S28. [Google Scholar] [CrossRef]
- Besedovsky, H.O.; del Rey, A. Regulating inflammation by glucocorticoids. Nat. Immunol. 2006, 7, 537. [Google Scholar] [CrossRef]
- Besedovsky, H.O.; del Rey, A. The cytokine-HPA axis feedback circuit. Z. Rheumatol. 2000, 59 Suppl. 2, II/26-30. [Google Scholar]
- Bourgeon, S.; Kauffmann, M.; Geiger, S.; Raclot, T.; Robin, J.P. Relationships between metabolic status, corticosterone secretion and maintenance of innate and adaptive humoral immunities in fasted re-fed mallards. J. Exp. Biol. 2010, 213, 3810–3818. [Google Scholar] [CrossRef]
- Silva, E.J.; Queiróz, D.B.; Rodrigues, A.; Honda, L.; Avellar, M.C. Innate immunity and glucocorticoids: Potential regulatory mechanisms in epididymal biology. J. Androl. 2011, 32, 614–624. [Google Scholar] [CrossRef]
- Azuma, M. Role of the glucocorticoid-induced TNFR-related protein (GITR)-GITR ligand pathway in innate and adaptive immunity. Crit. Rev. Immunol. 2010, 30, 547–557. [Google Scholar] [CrossRef]
- Baschant, U.; Tuckermann, J. The role of the glucocorticoid receptor in inflammation and immunity. J. Steroid Biochem. Mol. Biol. 2010, 120, 69–75. [Google Scholar] [CrossRef]
- Davison, T.F.; Flack, I.H. Changes in the peripheral blood leucocyte populations following an injection of corticotrophinin the immature chicken. Res. Vet. Sci. 1981, 30, 79–82. [Google Scholar]
- Mumma, J.O.; Thaxton, J.P.; Vizzier-Thaxton, Y.; Dodson, W.L. Physiological stress in laying hens. Poult. Sci. 2006, 85, 761–769. [Google Scholar]
- Puvadolpirod, S.; Thaxton, J.P. Model of physiological stress in chickens. 1. Response parameters. Poult. Sci. 2000, 79, 363–369. [Google Scholar]
- Croiset, G.; Heijnen, C.J.; Veldhuis, H.D.; de Wied, D.; Ballieux, R.E. Modulation of the immune response by emotional stress. Life Sci. 1987, 40, 775–782. [Google Scholar] [CrossRef]
- Persoons, J.H.; Moes, N.M.; Broug-Holub, E.; Schornagel, K.; Tilders, F.J.; Kraal, G. Acute and long-term effects of stressors on pulmonary immune functions. Am. J. Respir. Cell. Mol. Biol. 1997, 17, 203–208. [Google Scholar]
- Terman, G.W.; Shavit, Y.; Lewis, J.W.; Cannon, J.T.; Liebeskind, J.C. Intrinsic mechanisms of pain inhibition: Activation by stress. Science 1984, 226, 1270–1277. [Google Scholar]
- Choi, H.S.; Lim, S.A.; Park, M.S.; Hwang, B.Y.; Lee, C.K.; Kim, S.H.; Lim, S.C.; Lee, M.K. Ameliorating effects of the ethanol extracts from Gynostemma Pentaphyllum on electric footshock stress. Kor. J. Pharmacogn. 2008, 39, 341–346. [Google Scholar]
- Kamei, H.; Noda, Y.; Nabeshima, T. The psychological stress model using motor suppression. Nihon Yakurigaku Zasshi. 1999, 113, 113–120. [Google Scholar] [CrossRef]
- Iizuka, S.; Ishige, A.; Komatsu, Y.; Matsumiya, T.; Inazu, M.; Takeda, H. Effects of Toki-shakuyaku-san on electric footshock stress in ovariectomized mice. Methods Find. Exp. Clin. Pharmacol. 1998, 20, 39–46. [Google Scholar] [CrossRef]
- Kohno, H.; Katayama, S.; Ohuchi, Y.; Ohkubo, Y. Possible neural mechanisms involved in footshock stress-induced enhancement of exploratory behavior in mice. Biol. Pharm. Bull. 1999, 22, 418–421. [Google Scholar] [CrossRef]
- Soria, G.; Barbano, M.F.; Maldonado, R.; Valverde, O. A reliable method to study cue-, priming-, and stress-induced reinstatement of cocaine self-administration in mice. Psychopharmacology (Berl) 2008, 199, 593–603. [Google Scholar] [CrossRef]
- Im, S.A.; Choi, H.S.; Hwang, B.Y.; Lee, M.K.; Lee, C.K. Augmentation of Immune Responses by oral administration of Gynostemma Pentaphyllum ethanol extract. Kor. J. Pharmacogn. 2009, 40, 35–40. [Google Scholar]
- Laky, K.; Fleischacker, C.; Fowlkes, B.J. TCR and Notch signaling in CD4 and CD8 T cell development. Immunol. Rev. 2006, 209, 274–283. [Google Scholar] [CrossRef]
- Purton, J.F.; Monk, J.A.; Liddicoat, D.R.; Kyparissoudis, K.; Sakkal, S.; Richardson, S.J.; Godfrey, D.I.; Cole, T.J. Expression of the glucocorticoid receptor from the 1A promoter correlates with T lymphocyte sensitivity to glucocorticoid-induced cell death. J. Immunol. 2004, 173, 3816–3824. [Google Scholar]
- Moynihan, J.A.; Ader, R.; Grota, L.J.; Schachtman, T.R.; Cohen, N. The effects of stress on the development of immunological memory following low-dose antigen priming in mice. Brain Behav. Immun. 1990, 4, 1–12. [Google Scholar] [CrossRef]
- Keller, S.E.; Weiss, J.M.; Schleifer, S.J.; Miller, N.E.; Stein, M. Suppression of immunity by stress: Effect of a graded series of stressors on lymphocyte stimulation in the rat. Science 1981, 213, 1397–1400. [Google Scholar]
- Lysle, D.T.; Lyte, M.; Fowler, H.; Rabin, B.S. Shock-induced modulation of lymphocyte reactivity: Suppression, habituation, and recovery. Life Sci. 1987, 41, 1805–1814. [Google Scholar] [CrossRef]
- Sonnenfeld, G.; Cunnick, J.E.; Armfield, A.V.; Wood, P.G.; Rabin, B.S. Stress-induced alterations in interferon production and class II histocompatibility antigen expression. Brain Behav. Immun. 1992, 6, 170–178. [Google Scholar] [CrossRef]
- Batuman, O.A.; Sajewski, D.; Ottenweller, J.E.; Pitman, D.L.; Natelson, B.H. Effects of repeated stress on T cell numbers and function in rats. Brain Behav. Immun. 1990, 4, 105–117. [Google Scholar] [CrossRef]
- Kusnecov, A.W.; Rabin, B.S. Inescapable footshock exposure differentially alters antigen- and mitogen-stimulated spleen cell proliferation in rats. J. Neuroimmunol. 1993, 44, 33–42. [Google Scholar] [CrossRef]
- Persoons, J.H.; Moes, N.M.; Broug-Holub, E.; Schornagel, K.; Tilders, F.J.; Kraal, G. Acute and long-term effects of stressors on pulmonary immune functions. Am. J. Respir. Cell. Mol. Biol. 1997, 17, 203–208. [Google Scholar]
- Pramanik, S.S.; Sur, T.K.; Debnath, P.K.; Bhattacharyya, D. Effect of Pueraria tuberosa tuber extract on chronic foot shock stress in Wistar rats. Nepal. Med. Coll. J. 2010, 12, 234–238. [Google Scholar]
- Nade, V.S.; Yadav, A.V. Anti-stress effect of ethyl acetate soluble fraction of Morus alba in chronic restraint stress. Pharm. Biol. 2010, 48, 1038–1046. [Google Scholar] [CrossRef]
- Habbu, P.V.; Mahadevan, K.M.; Kulkarni, P.V.; Daulatsingh, C.; Veerapur, V.P.; Shastry, R.A. Adaptogenic and in vitro antioxidant activity of flavanoids and other fractions of Argyreia speciosa (Burm.f) Boj. in acute and chronic stress paradigms in rodents. Indian J. Exp. Biol. 2010, 48, 53–60. [Google Scholar]
- Umukoro, S.; Aladeokin, A.C. Evaluation of the anti-stress and anticonvulsant activities of leaf extract of Alchornea cordifolia in mice. J. Ethnopharmacol. 2010, 127, 768–770. [Google Scholar] [CrossRef]
- Piato, A.L.; Detanico, B.C.; Linck, V.M.; Herrmann, A.P.; Nunes, D.S.; Elisabetsky, E. Anti-stress effects of the “tonic” Ptychopetalum olacoides (Marapuama) in mice. Phytomedicine 2010, 17, 248–253. [Google Scholar] [CrossRef]
- Rai, D.; Bhatia, G.; Sen, T.; Palit, G. Anti-stress effects of Ginkgo biloba and Panax ginseng: A comparative study. J. Pharmacol. Sci. 2003, 93, 458–464. [Google Scholar] [CrossRef]
- He, R.R.; Yao, X.S.; Li, H.Y.; Dai, Y.; Duan, Y.H.; Li, Y.F.; Kurihara, H. The anti-stress effects of Sarcandra glabra extract on restraint-evoked immunocompromise. Biol. Pharm. Bull. 2009, 32, 247–252. [Google Scholar] [CrossRef]
- Lin, J.M.; Lin, C.C.; Chiu, H.F.; Yang, J.J.; Lee, S.G. Evaluation of the anti-inflammatory and liver-protective effects of Anoectochilus formosanus, Ganoderma lucidum and Gynostemma pentaphyllum in rats. Am. J. Chin. Med. 1993, 21, 59–69. [Google Scholar] [CrossRef]
- Shang, L.; Liu, J.; Zhu, Q.; Zhao, L.; Feng, Y.; Wang, X.; Cao, W.; Xin, H. Gypenosides protect primary cultures of rat cortical cells against oxidative neurotoxicity. Brain Res. 1102, 163–174. [Google Scholar]
- Wang, Z.J.; Luo, D.H. Antioxidant activities of different fractions of polysaccharide purified from Gynostemma pentaphyllum Makino. Carbohyd. Polym. 2007, 68, 54–58. [Google Scholar] [CrossRef]
- Lee, Y.H.; Lee, Y.R.; Im, S.A.; Park, S.I.; Kim, K.H.; Gerelchuluun, T.; Song, S.; Kim, K.; Lee, C.K. Calcineurin inhibitors block MHC-restricted antigen presentation in vivo. J. Immunol. 2007, 179, 5711–5716. [Google Scholar]
- Sample Availability: Samples of GP-EX are available from the author (M.K.L.: myklee@chungbuk.ac.kr).
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Im, S.-A.; Choi, H.S.; Choi, S.O.; Kim, K.-H.; Lee, S.; Hwang, B.Y.; Lee, M.K.; Lee, C.K. Restoration of Electric Footshock-Induced Immunosuppression in Mice by Gynostemma pentaphyllum Components. Molecules 2012, 17, 7695-7708. https://doi.org/10.3390/molecules17077695
Im S-A, Choi HS, Choi SO, Kim K-H, Lee S, Hwang BY, Lee MK, Lee CK. Restoration of Electric Footshock-Induced Immunosuppression in Mice by Gynostemma pentaphyllum Components. Molecules. 2012; 17(7):7695-7708. https://doi.org/10.3390/molecules17077695
Chicago/Turabian StyleIm, Sun-A, Hyun Sook Choi, Soon Ok Choi, Ki-Hyang Kim, Seungjeong Lee, Bang Yeon Hwang, Myung Koo Lee, and Chong Kil Lee. 2012. "Restoration of Electric Footshock-Induced Immunosuppression in Mice by Gynostemma pentaphyllum Components" Molecules 17, no. 7: 7695-7708. https://doi.org/10.3390/molecules17077695
APA StyleIm, S.-A., Choi, H. S., Choi, S. O., Kim, K.-H., Lee, S., Hwang, B. Y., Lee, M. K., & Lee, C. K. (2012). Restoration of Electric Footshock-Induced Immunosuppression in Mice by Gynostemma pentaphyllum Components. Molecules, 17(7), 7695-7708. https://doi.org/10.3390/molecules17077695