Camptothecin-20(s)-O-[N-(3’α,12’α-dihydroxy-24’-carbonyl-5’β-cholan)]-lysine, a Novel Camptothecin Analogue, Induces Apoptosis towards Hepatocellular Carcinoma SMMC-7721 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
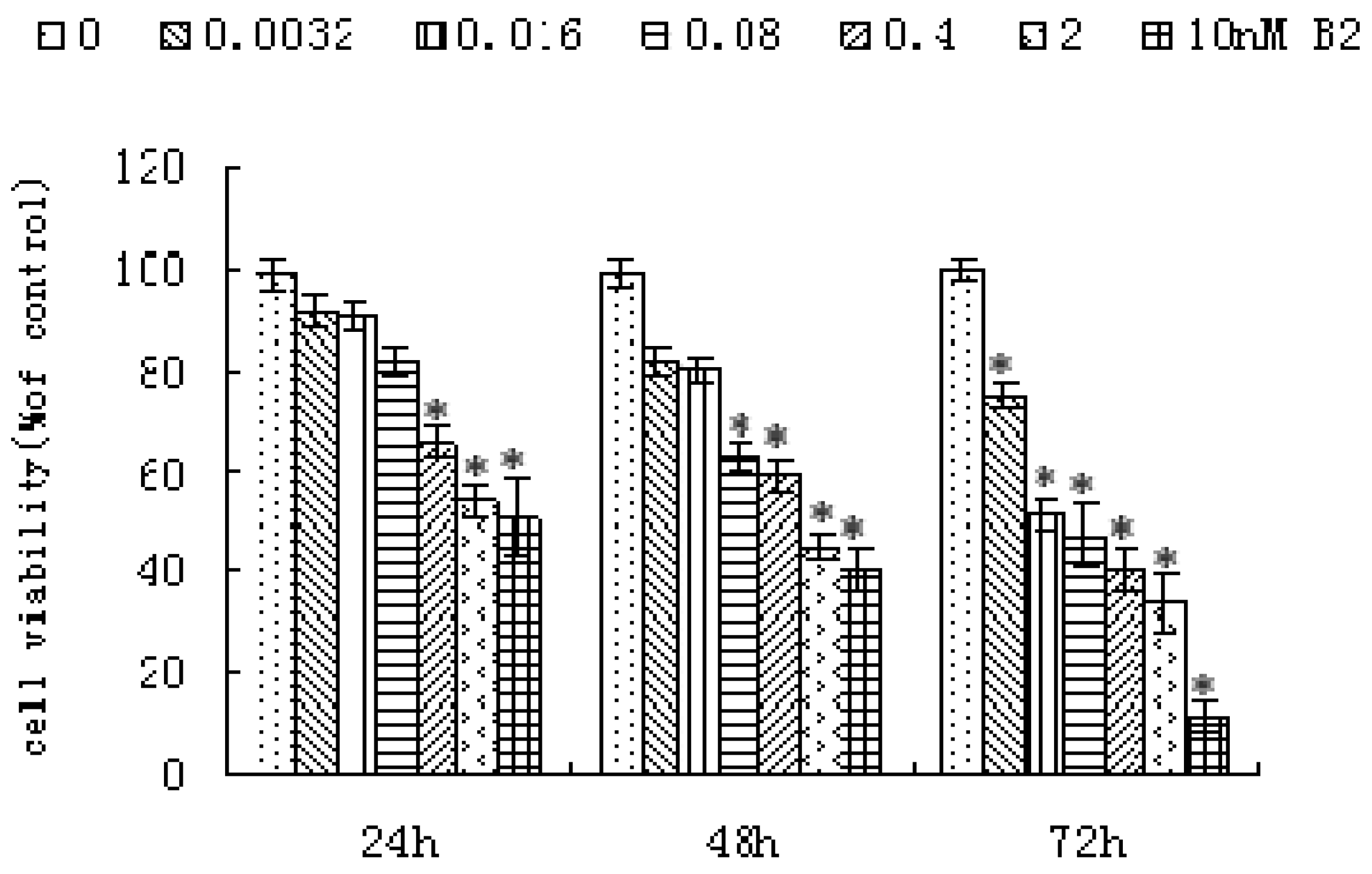
2.1. Anti-Proliferative Activity of B2
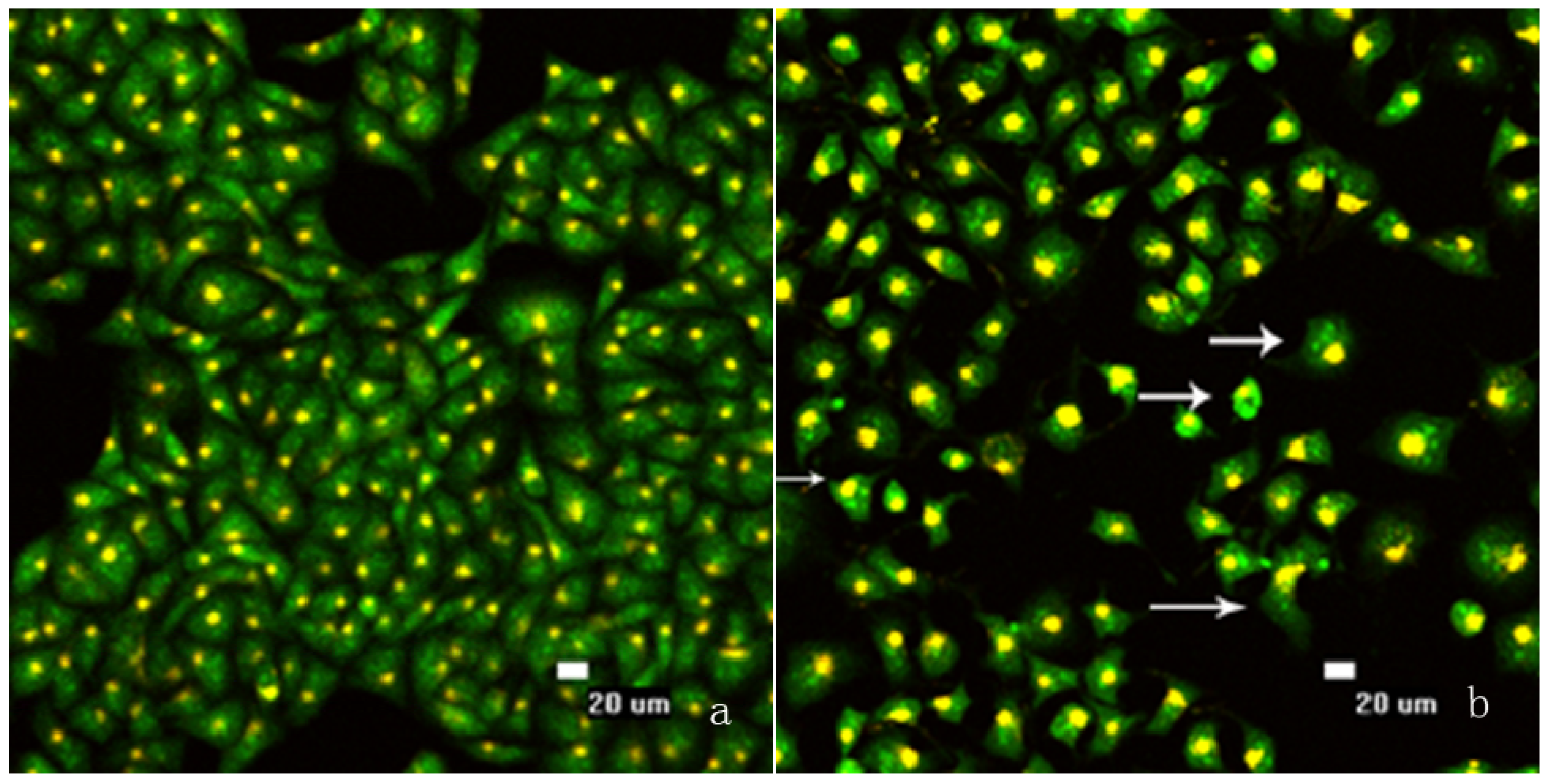
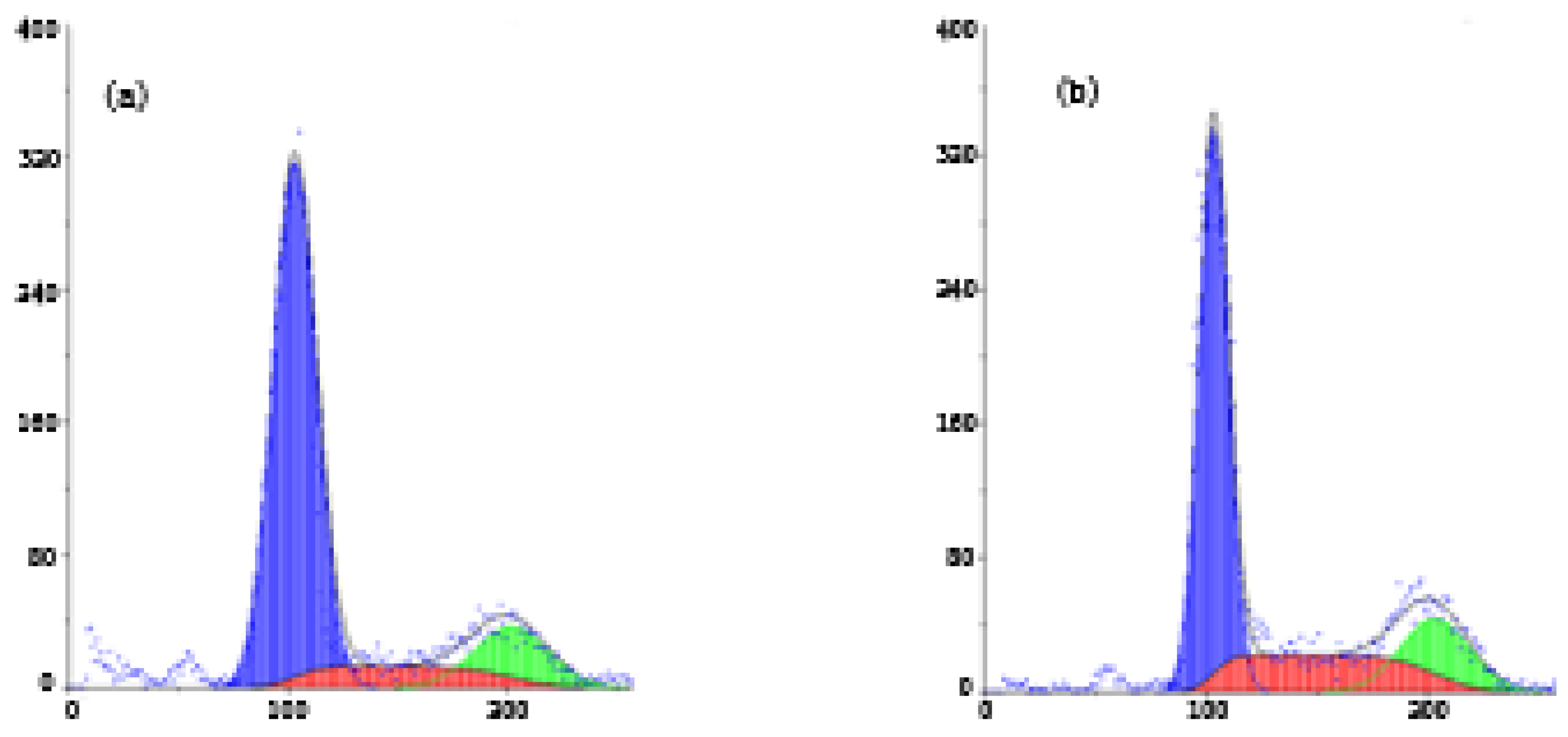
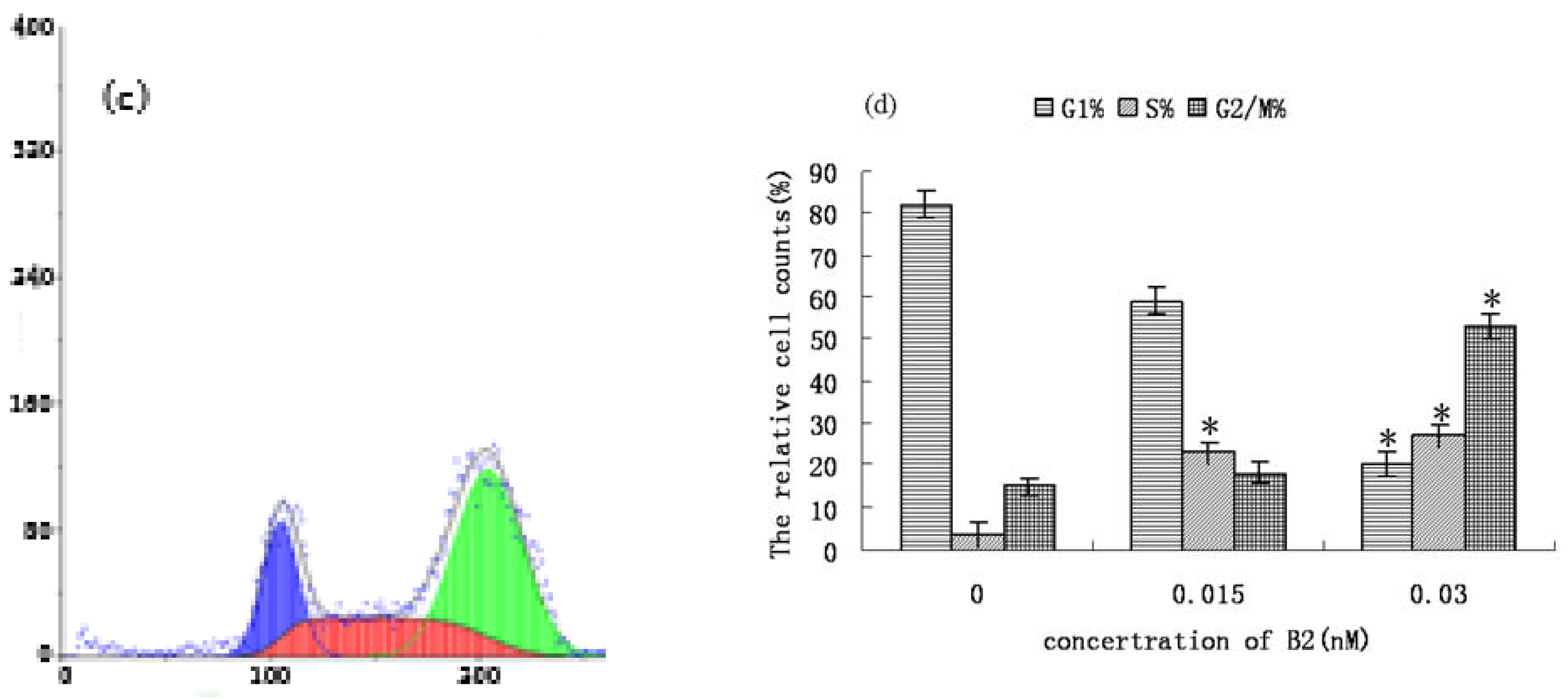
2.2. FCM Analysis of SMMC-7721 Cell Cycle Distribution and Apoptosis
2.3. B2 Induces SMMC-7721 Apoptosis via Mitochondrion Pathway
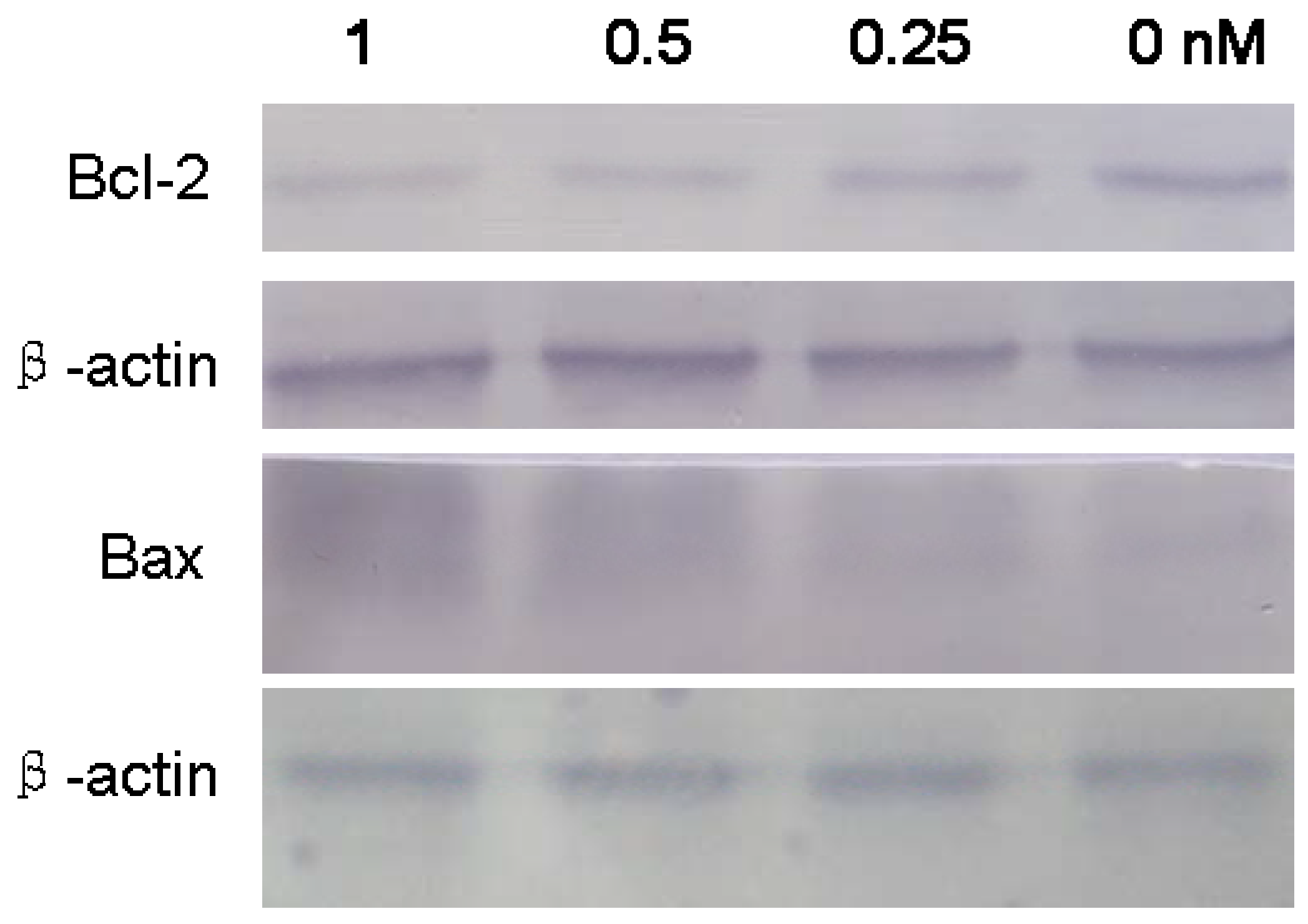
2.4. Apoptosis-Related Proteins in SMMC-7721 Treated with B2
3. Experimental
3.1. Materials
3.2. Cell Culture
3.3. Cell Proliferation Assay
3.4. Morphological Observation of Nuclear Change
3.5. Apoptosis Assays and Cell Cycle Analysis
3.6. Assessment of Mitochondrial Membrane Potential (ΔΨ m)
3.7. Determination of the Changes of Calcium and ROS in Cells
3.8. Measurement of Caspase-3 and Caspase-9 Activities
3.9. Western Blot Analysis
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Cai, R.L.; Meng, W.; Lu, H.Y.; Lin, W.Y.; Jiang, F.; Shen, F.M. Segregation analysis of hepatocellular carcinoma in a moderately high-incidence area of East China. World J. Gastroenterol. 2003, 9, 2428–2432. [Google Scholar] [CrossRef] [PubMed]
- Farmer, D.G.; Rosove, M.H.; Shaked, A.; Busuttil, R.W. Current treatment modalities for hepatocellular carcinoma. Ann. Surg. 1994, 219, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Ying, M.D.; Tu, C.X.; Ying, H.Z.; Hu, Y.Z.; He, Q.J.; Yang, B. MSFTZ, a flavanone derivative, induces human hepatoma cell apoptosis via a reactive oxygen species- and caspase dependent mitochondrial pathway. J. Pharmacol. Exp. Ther. 2008, 325, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Wall, M.E.; Wani, M.C.; Cook, C.E.; Palmer, K.H.; Mcphail, A.T.; Sim, G.A. The isolation and structure of Camptothecin, a novel alkaloidal leukemia and tumor inhibitor from camptotheca acuminate. J. Am. Chem. Soc. 1966, 88, 3888–3890. [Google Scholar] [CrossRef]
- He, X.G.; Lu, W.; Jiang, X.Q.; Cai, J.C.; Zhang, X.W.; Ding, J. Synthesis and biological evaluation of bis and monocarbonate prodrugs of 10-hydroxycamptothecins. Bioorg. Med. Chem. 2004, 12, 4003–4008. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Gao, Y.; Qiu, W.; Zu, Y.G.; Su, L. synthesis and anti-tumor activity of novel camptothecin-bile acid analogues. Lett. Drug Discov. 2011, 8, 698–703. [Google Scholar] [CrossRef]
- Frankfurt, O.S.; Krishan, A. Apoptosis-based drug screening and detection of selective toxicity to cancer cells. Anticancer Drugs 2003, 14, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.F. DNA topoisomerase poisons as antitumor drugs. Annu. Rev. Biochem. 1989, 58, 351–75. [Google Scholar] [CrossRef] [PubMed]
- D’Arpa, P.; Beardmore, C.; Liu, L.F. Involvement of nucleic acid synthesis in cell killing mechanisms of topoisomerase poisons. Cancer Res. 1990, 50, 6919–6924. [Google Scholar] [PubMed]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutelingsperger, C. A novel assay for apoptosis, Flow cytometric detection of phosphatidylserin expression on early apoptotic cells using fluorescein labeled annexin-V. J. Immunol. Meth. 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Yan, Z.H.; Yang, R.L.; Jiang, Y.; Yang, Z.H. Induction of apoptosis in human promyelocytic Leukemia HL60 cells by panaxynol and panaxydol. Molecules 2011, 16, 5561–5573. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.F. Relation between intracellular acidification and camptothecin-induced apoptosis in leukemia cells. Eur. J. Pharm. Sci. 2000, 10, 125–131. [Google Scholar] [CrossRef]
- Fu, Y.R.; Yi, Z.J.; Yan, Y.R.; Qiu, Z.Y. Hydroxycamptothecin-induced apoptosis in hepatoma SMMC-7721 cells and the role of mitochondrial pathway. Mitochondrion 2006, 6, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Gogvadze, V.; Orrenius, S. Mitochondrial regulation of apoptotic cell death. Chem. Biol. Interact. 2006, 163, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R. Apoptotic pathways: The roads to run. Cell 1998, 94, 695–698. [Google Scholar] [CrossRef]
- Yoneyama, K. Validation of confocal laser scanning microscopy for detecting intracellular calcium heterogeneity in live slices. J. Pharmacol. Toxicol. Meth. 2001, 45, 187–193. [Google Scholar] [CrossRef]
- Demaurex, N.; Distelhorst, C. Apoptosis--the Calcium Connection. Science 2003, 300, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Goldstein, L.A.; Gastman, B.R.; Rabinowich, H. Interrelated roles for Mcl-1 and BIM in regulation of TRAIL-mediated mitochondrial apoptosis. J. Biol. Chem. 2006, 281, 10153–10163. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Zamzami, N.; Susin, S. Mitochondrial control of apoptosis. Immunol. Today 1997, 18, 44–51. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Lazebnik, Y. Caspases: Enemies within. Science 1998, 281, 1312–1316. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, J.H. ROS-dependent caspase-9 activation in hypoxic cell death. FEBS Lett. 2003, 549, 94–98. [Google Scholar] [CrossRef]
- Krammer, P.H.; Debatin, K.M. Death receptors in chemotherapy and cancer. Oncogene 2004, 23, 2950–2966. [Google Scholar]
- Antonsson, B.; Martinou, J.C. The Bcl-2 protein family. Exp. Cell Res. 2000, 256, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.C. Double identity for proteins of the Bcl-2 family. Nature 1997, 387, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival, application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Croce, A.C.; Bottiroli, G.; Supino, R. Subcellular localization of the camptothecin analogues, topotecan and gimatecan. Biochemi. Pharmacol. 2004, 67, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, D.; Wokosin, D.; Girkin, J.; Grant, M.H. Measurement of the intracellular distribution of reduced glutathione in cultured rat hepatocytes using monochlorobimane and confocallaser scanning microscopy. Toxicol. In Vitro 2002, 16, 609–619. [Google Scholar] [CrossRef]
- Blagosklonny, M.V.; Pardee, A.B. Exploiting cancer cell cycling for selective protection of normal cells. Cancer Res. 2002, 61, 4301–4305. [Google Scholar]
- Giri, B.; Gomes, A.; Debnath, A. Antiproliferative, cytotoxic and apoptogenic activity of Indian toad (Bufo melanostictus, Schneider) skin extract on U937 and K562 cells. Toxicon 2006, 48, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhu, Y.S.; Guo, M.; Yua, Y.; Chen, G.Q. Therapeutic efficacy of NSC606985, a novel camptothecin analog, in a mouse model of acute promyelocytic leukemia. Leuk. Res. 2007, 31, 1565–1574. [Google Scholar] [CrossRef] [PubMed]
- Zamzami, N.; Maisse, C.; Metivier, D.; Kroemer, G. Measurement of membrane permeability and permeability transition of mitochondria. Methods Cell Biol. 2001, 65, 147–158. [Google Scholar] [PubMed]
- Palmeira, C.M.; Moreno, A.J.M.; Madeira, V.M.C.; Wallace, K.B. Continuous monitoring of mitochondrial membrane potential in hepatocyte cell suspensions. J. Pharmacol. Toxicol. Methods 1996, 35, 35–43. [Google Scholar] [CrossRef]
- Bhutia, S.K.; Mallick, S.K.; Maiti, S.; Mishra, D.; Maiti, T.K. Abrus abrin derived peptides induce apoptosis by targeting mitochondria in HeLa cells. Cell Biol. Int. 2009, 33, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Li, Q.Y.; Zu, Y.G.; Fu, Y.J. Anti-proliferative and pro-apoptotic effect of CPT13, a novel camptothecin analog, on human colon cancer HCT8 cell line. Chem. Biol. Interact. 2008, 176, 165–172. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compound are available from the authors. |








© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Li, Q.; Qiu, W.; Zhu, Q.; Zu, Y.; Deng, X.; Zhao, T.; Jiang, C.; Zhang, L. Camptothecin-20(s)-O-[N-(3’α,12’α-dihydroxy-24’-carbonyl-5’β-cholan)]-lysine, a Novel Camptothecin Analogue, Induces Apoptosis towards Hepatocellular Carcinoma SMMC-7721 Cells. Molecules 2011, 16, 7803-7814. https://doi.org/10.3390/molecules16097803
Li Q, Qiu W, Zhu Q, Zu Y, Deng X, Zhao T, Jiang C, Zhang L. Camptothecin-20(s)-O-[N-(3’α,12’α-dihydroxy-24’-carbonyl-5’β-cholan)]-lysine, a Novel Camptothecin Analogue, Induces Apoptosis towards Hepatocellular Carcinoma SMMC-7721 Cells. Molecules. 2011; 16(9):7803-7814. https://doi.org/10.3390/molecules16097803
Chicago/Turabian StyleLi, Qingyong, Wei Qiu, Qiaochu Zhu, Yuangang Zu, Xiaoqiu Deng, Tengfei Zhao, Chunfei Jiang, and Li Zhang. 2011. "Camptothecin-20(s)-O-[N-(3’α,12’α-dihydroxy-24’-carbonyl-5’β-cholan)]-lysine, a Novel Camptothecin Analogue, Induces Apoptosis towards Hepatocellular Carcinoma SMMC-7721 Cells" Molecules 16, no. 9: 7803-7814. https://doi.org/10.3390/molecules16097803