2D-DIGE Proteomic Analysis of Changes in Estrogen/Progesterone-Induced Rat Breast Hyperplasia upon Treatment with the Mongolian Remedy RuXian-I

Abstract
:1. Introduction
2. Results and Discussion
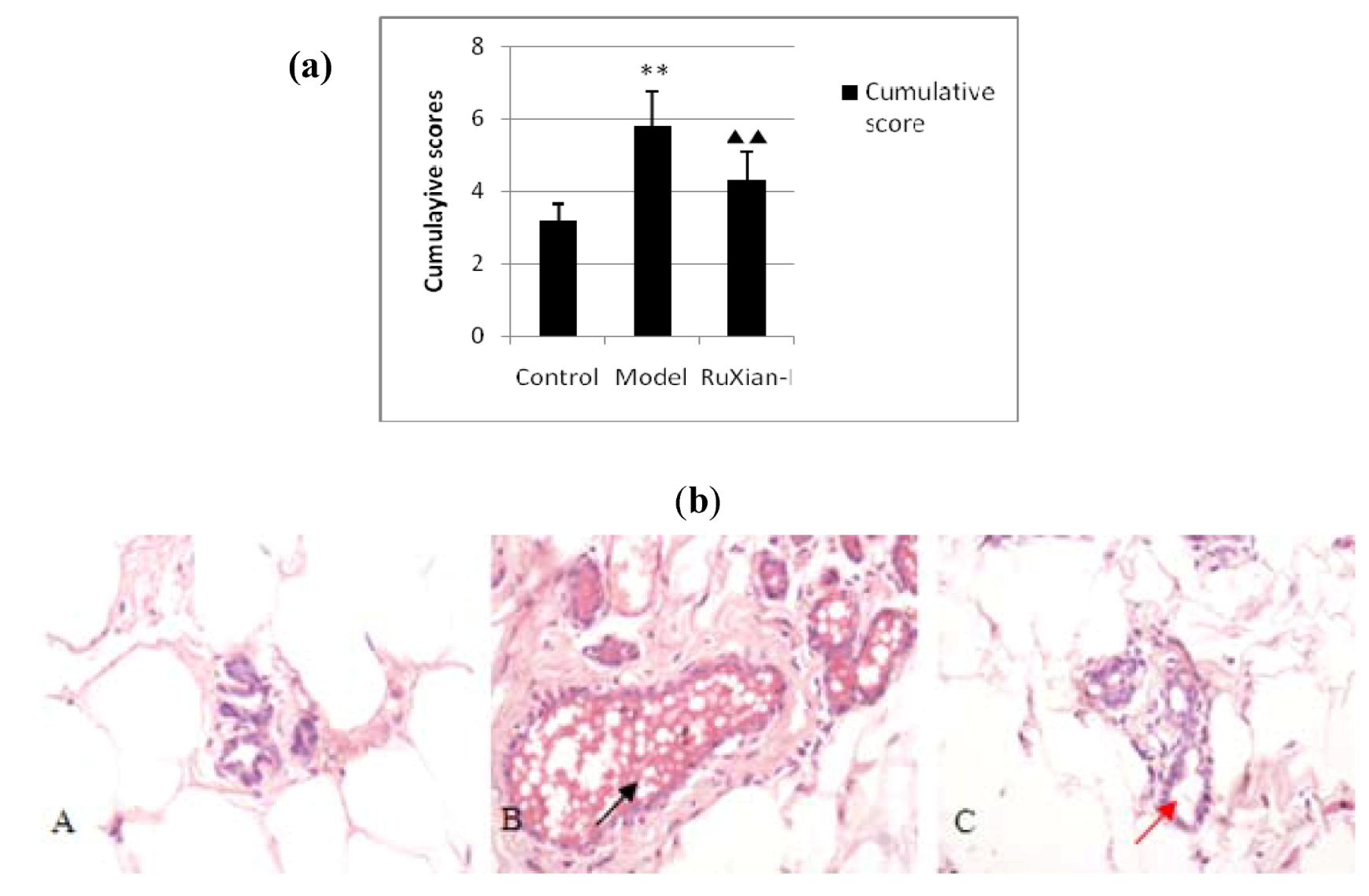
2.1. Histopathological observation of breast tissue by H&E staining

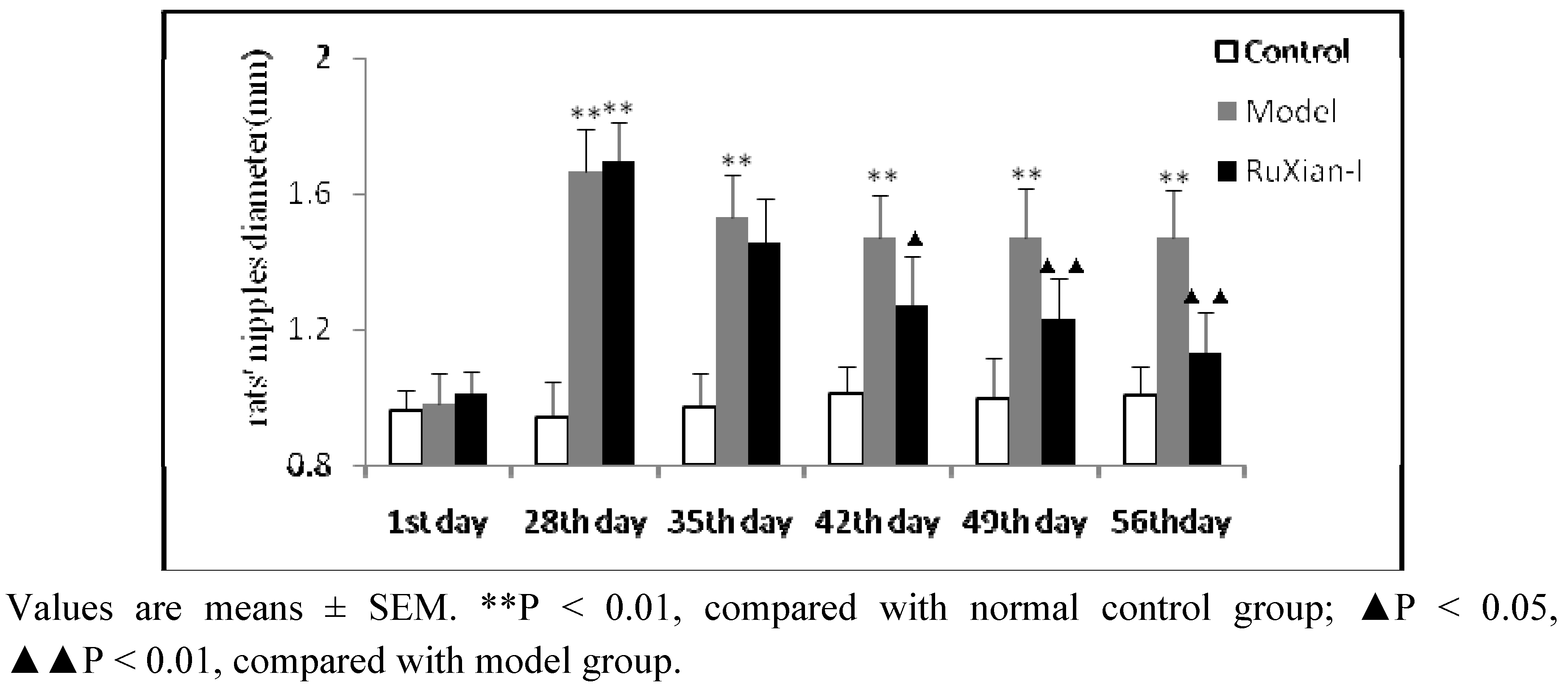
2.2. Comparison of nipple diameters in rats

2.3. Proteomic analysis to identify changes in protein expression stimulated by RuXian-I treatment in rat breast hyperplasia
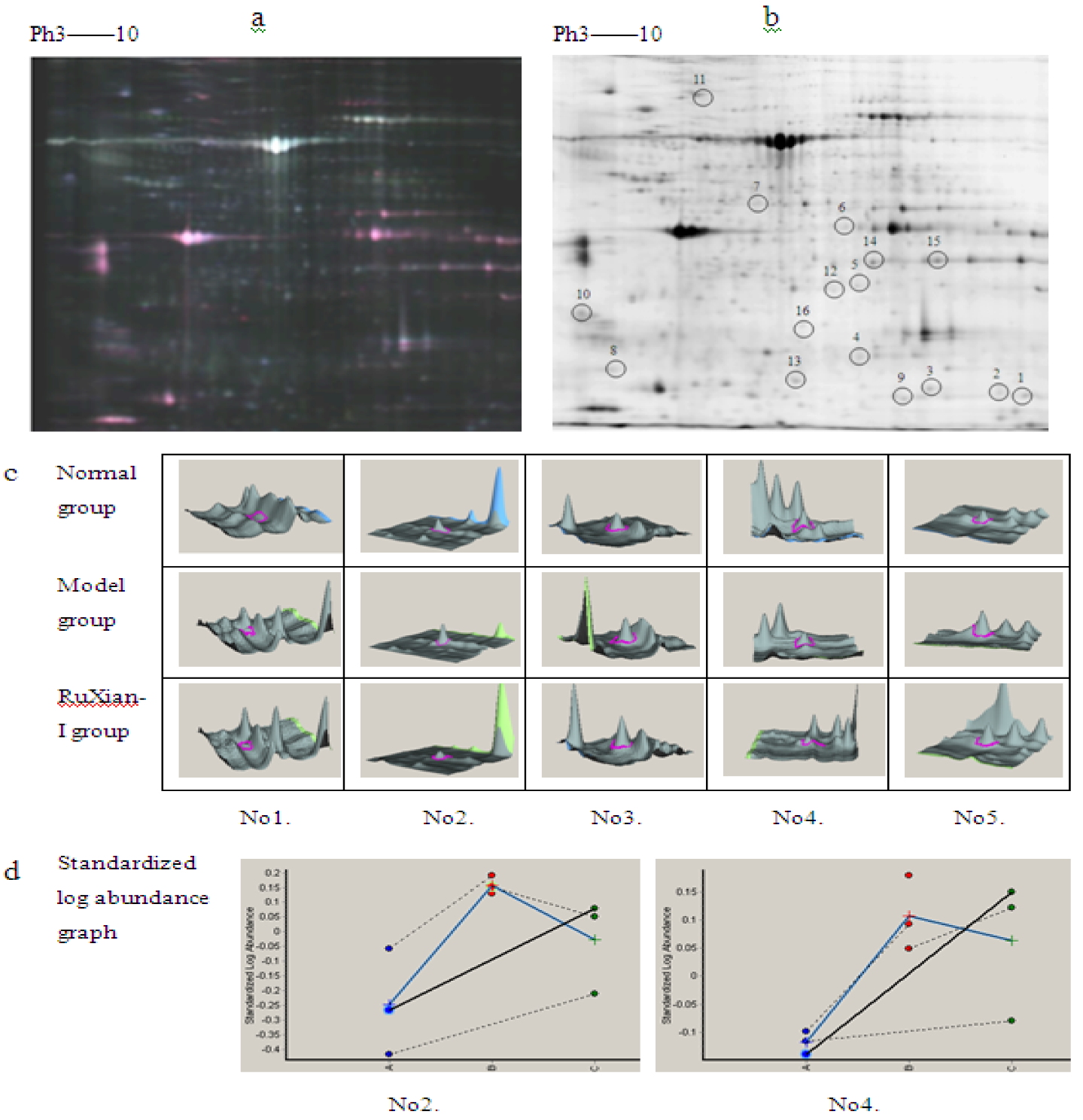
2.3.1. Analysis of Differentially Expressed Proteins

2.3.2. Identfication of Differentially Expressed Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO.a | Protein description | Protein ID | Protein levelb | Nominal | Percentage | Mascot Scoree | Molecular function |
|---|---|---|---|---|---|---|---|
| Mr(kDa)/p Ic | coverage (%)d | ||||||
| 1 | Peroxiredoxin-1 | Q63716 | ↑ | 22.07/8.34 | 35 | 197 | oxidoreductase activity and peroxidase activity |
| 2 | Superoxide dismutase [Mn] | P07895 | ↑ | 24.659/8.96 | 41 | 108 | oxidoreductase activity |
| 3 | Tpi1 protein | P48500 | ↓ | 26.701/7.07 | 43 | 112 | isomerase activity |
| 4 | Glutathione S-transferase P | P04906 | ↑ | 23.424/6.89 | 39 | 117 | transferase activity |
| 5 | Omega-amidase NIT2 | P30919 | ↑ | 30.682/6.9 | 35 | 156 | hydrolase activity |
| 6 | Glutamine synthetase | P09606 | ↑ | 42.24/6.64 | 25 | 103 | ligase activity |
| 7 | Xaa-Pro dipeptidase | Q510D7 | ↑ | 54.71/5.61 | 42 | 136 | peptidase activity, DNA binding, transcription factor activity |
| 8 | Translationally-controlled tumor protein | P63029 | ↑ | 14.949/5.09 | 41 | 102 | structural constituent of cytoskeleton and cytoskeletal protein binding |
| 9 | Alpha B-crystallin | P02511 | ↑ | 19.945/6.84 | 66 | 316 | structural molecule activity |
| 10 | Tropomyosin alpha-4 chain | P09495 | ↑ | 28.492/4.66 | 47 | 107 | motor activity and structural constituent of cytoskeleton |
| 11 | Myosin-4 | Q29RW1 | ↓ | 222.7/5.58 | 16 | 118 | motor activity, structural constituent of cytoskeleton, protein binding and small GTPase requlator activity |
| 12 | Complement component 4 | Q6MG90 | ↑ | 192.00/6.62 | 9 | 156 | receptor binding and peptidase inhibitor activity |
| 13 | Immunoglobulin kappa constant region | XP575527 | ↑ | 23.42/5.94 | 39 | 142 | antigen binding |
| 14 | Annexin A1 | P07150 | ↑ | 38.805/6.97 | 71 | 417 | calcium ion binding, calcium-dependent phospholipid binding |
| 15 | Annexin A2 | Q07936 | ↑ | 38.6/7.55 | 62 | 527 | calcium ion binding, calcium-dependent phospholipid binding |
| 16 | Beta-casein | P02665 | ↑ | 25.353/6.04 | 19 | 88 |
| Biological functions | Protein description |
|---|---|
| immune system process | complement component 4; immunoglobulin kappa V region; alpha B-crystallin; translationally controlled tumor protein; glutathione S-transferase P; superoxide dismutase [Mn]; peroxiredoxin-1 |
| metabolic process | |
| protein metabolic process | alpha B-crystallin; complement component 4; xaa-Pro dipeptidase; omega-amidase NIT2 |
| lipid metabolic process | annexin A1; annexin A2 |
| nitrogen compound metabolic process | glutamine synthetase |
| cellular amino acid and derivative metabolic process | glutamine synthetase |
| carbohydrate metabolic process | Tpi1 protein |
| oxygen and reactive oxygen species metabolic process | superoxide dismutase [Mn]; peroxiredoxin-1 |
| response to stimulus | alpha B-crystallin; glutathione S-transferase P; complement component 4; immunoglobulin kappa V region |
| transport | annexin A1; annexin A2; Beta-casein; myosin-4 |
| signal transduction | complement component 4; myosin-4; annexin A1; annexin A2 |
| developmental process | immunoglobulin kappa V region; tropomyosin alpha-4 chain; myosin-4; annexin A2 |
| cell motion | tropomyosin alpha-4 chain; annexin A1; annexin A2 |
| cellular component organization | tropomyosin alpha-4 chain; myosin-4 |
| cell cycle | myosin-4 |
| muscle contraction | alpha B-crystallin; tropomyosin alpha-4 chain; myosin-4 |
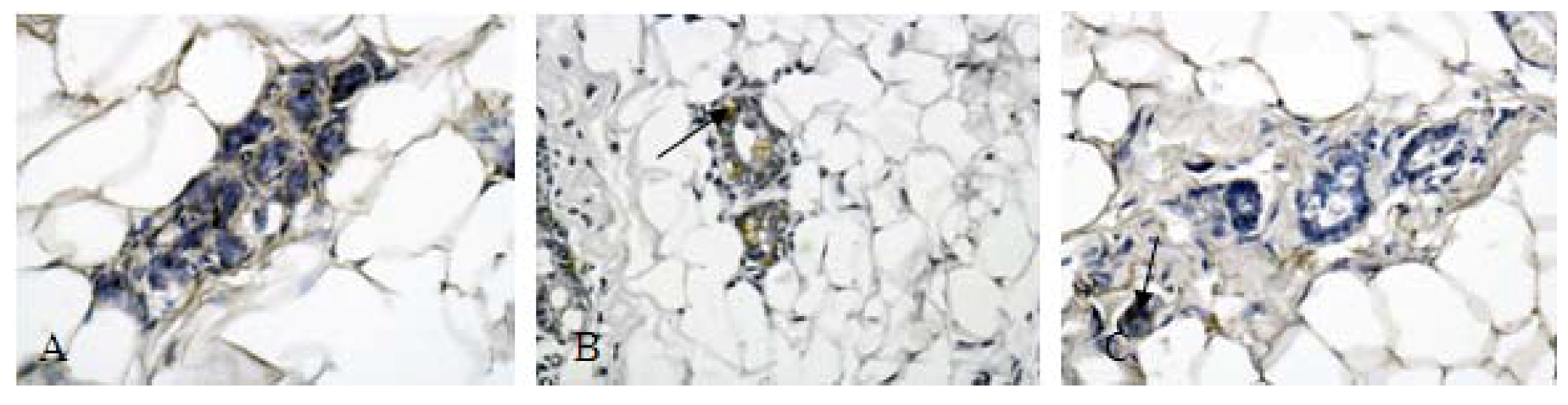
2.4. Protein Validation


3. Experimental
3.1. Reagents
3.2. Preparation of RuXian-I
| Raw materials a and dose | |||
|---|---|---|---|
| Latin name | Medicine site | Dose | |
| Raw materials processed by extracting with distilled water a | |||
| (1) | Fructus Amomi Rotundus (Zingiberaceae) | Fruit | 250.0 g |
| (2) | Herba Leonuri Japonici (Labiatae) | Whole herb | 75.0 g |
| (3) | Flos Caryophylli (Myrtaceae) | Alabastrum | 75.0 g |
| (4) | Fructus Gardeniae (Rubiaceae) | Fruit | 50.0 g |
| (5) | Rhizoma Gymnadeniae Conopseae (Orchidaceae) | Stem tuber | 50.0 g |
| (6) | Semen Myristicae (Myristicaceae) | Kernal | 50.0 g |
| (7) | Lignum Aquilariae Resinatum (Thymelaeaceae) | The wood contains Resina | 50.0 g |
| (8) | Fructus Hippophae (Elaeagnaceae) | Fruit | 50.0 g |
| (9) | Thladiantha Dubia Bge (Cucurbitaceae) | Stem tuber | 50.0 g |
| (10) | Sorbaria Sorbifolia (Rosaceae) | Kernal | 50.0 g |
| (11) | Fructus chebulae (Combretaceae) | Fruit | 25.0 g |
| (12) | Rhizoma Kaempferiae (Zingiberaceae) | Rhizome | 25.0 g |
| (13) | Flos Carthami (Compositae) | Flower | 25.0 g |
| (14) | Radix Aucklandiae (Compositae) | Root | 25.0 g |
| (15) | Inula Helenium L. (Compositae) | Root | 25.0 g |
| (16) | Sabina chinensis (Cupressaceae) | Fruit | 25.0 g |
| (17) | Faeces Trogopteri (Petauristidae) | Feces | 25.0 g |
| (18) | Semen Cyclogalanopsis Glaucae (Fagaceae) | Kernal | 25.0 g |
| (19) | Herba Artemisiae Scopariae (Compositae) | Seedlings | 15.0 g |
| (20) | Lacca (Lacciferidae) | Glue secretion in the branches | 10.0 g |
| (21) | Folium Eriobotryae Japonicae (Rosaceae) | Leaf | 10.0 g |
| (22) | Radix Arnebiae (Boraginaceae) | Root | 10.0 g |
| (23) | Trollius Chinensis Bunge (Ranunculaceae) | Flower | 10.0 g |
| (24) | Radix Sophorae Flavescentis (Leguminosae) | Root | 10.0 g |
| (25) | Fructus Toosendan (Meliaceae) | Fruit | 10.0 g |
| (26) | Radix Rubiae Cordifoliae (Rubiaceae) | Root | 10.0 g |
| Raw materials processed by direct crushing a | |||
| (27) | Calculus Bovis (Bovidae) | Gallstone | 5.0 g |
| (28) | Cornu Cervi Pantotrichum (Cervidae) | Immature horns | 3.0 g |
| (29) | Cordyceps (Clavicepitaceae) | Stroma | 2.0 g |
| (30) | Borax (Borate) | Mineral | 2.0 g |
3.3. Rat model of breast hyperplasia and medications
3.4. Histopathological observation of breast tissue
3.5. Protein sample preparation
3.6. Fluorescence labeling with CyDyes
3.7. 2D-DIGE
3.8. Image acquisition and analysis
3.9. In-gel digestion of proteins and MALDI-TOF/TOF-MS protein identification
3.10. Immunohistochemical detection of annexin A1 and SOD activity assay
4. Conclusions
Acknowledgements
References
- Semiglazov, V.F.; Maksimov, S.I.; Bulgatova, E.A.; Meshkova, I.E.; Chepik, O.F.; Berstein, L.M. Risk of endometrial hyperplasia and carcinoma in breast cancer patients receiving adjuvant tamoxifen. Vopr. Onkol. 2003, 49, 198–204. [Google Scholar]
- Von Eggeling, F.; Gawriljuk, A.; Fiedler, W.; Ernst, G.; Claussen, U.; Klose, J.; Römer, I. Fluorescent dual colour 2D-protein gelelectrophoresis for rapid detection of differences in protein pattern with standard image analysis software. Int. J. Mol. Med. 2001, 8, 373–377. [Google Scholar]
- Hodgkinson, V.C.; Eagle, G.L.; Drew, P.J.; Lind, M.J.; Cawkwell, L. Biomarkers of chemotherapy resistance in breast cancer identified by proteomics: current status. Cancer. Lett. 2010, 294, 13–24. [Google Scholar] [CrossRef]
- Braun, M.; Fountoulakis, M.; Papadopoulou, A.; Vougas, K.; Seidel, I.; Höller, T.; Yeghiazaryan, K.; Schild, H.H.; Kuhn, W.; Golubnitschaja, O. Down-regulation of Microfilamental Network-associated Proteins in Leukocytes of Breast Cancer Patients: Potential Application to Predictive Diagnosis. Cancer Genom.Proteom. 2009, 6, 31–40. [Google Scholar]
- Umar, A.; Kang, H.; Timmermans, A.M.; Look, M.P.; Meijer-van Gelder, M.E.; den Bakker, M.A.; Jaitly, N.; Martens, J.W.; Luider, T.M.; Foekens, J.A.; Pasa-Tolić, L. Identification of a putative protein profile associated with tamoxifen therapy resistance in breast cancer. Mol. Cell. Proteom. 2009, 8, 1278–1294. [Google Scholar] [CrossRef]
- Betancourt, A.M.; Mobley, J.A.; Russo, J.; Lamartiniere, C.A. Proteomic analysis in mammary glands of rat offspring exposed in utero to bisphenol A. J. Proteom. 2010, 73, 1241–1253. [Google Scholar]
- Bocchinfuso, W.P.; Lindzey, J.K.; Hewitt, S.C.; Clark, J.A.; Myers, P.H.; Cooper, R.; Korach, K.S. Induction of mammary gland development in estrogen receptor-alpha knockout mice. Endocrinology 2000, 141, 2982–2994. [Google Scholar]
- Rohan, T.E.; Negassa, A.; Chlebowski, R.T.; Lasser, N.L.; McTiernan, A.; Schenken, R.S.; Ginsberg, M.; Wassertheil-Smoller, S.; Page, D.L. Estrogen plus progestin and risk of benign proliferative breast disease. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 2337–2343. [Google Scholar]
- Crandall, C.J.; Karlamangla, A.; Huang, M.H.; Ursin, G.; Guan, M.; Greendale, G.A. Association of new-onset breast discomfort with an increase in mammographic density during hormone therapy. Arch. Intern. Med. 2006, 166, 1578–1584. [Google Scholar]
- Conner, P. Breast response to menopausal hormone therapy-aspects on proliferation, apoptosis and mammographic density. Ann. Med. 2007, 39, 28–41. [Google Scholar] [CrossRef]
- Ahn, S.H.; Sawada, H.; Ro, J.Y.; Nicolson, G.L. Differential expression of annexinI in human mammary ductal epithelial cells in normal and benign and malignant breast tissues. Clin. Exp. Metastasis 1997, 15, 151–156. [Google Scholar] [CrossRef]
- Mulla, A.; Christian, H.C.; Solito, E.; Mendoza, N.; Morris, J.F.; Buckingham, J.C. Expression, subcellular localization and phosphorylation status of annexins 1 and 5 in human pituitary adenomas and a growth hormone-secreting carcinoma. Clin. Endocrinol. (Oxf.). 2004, 60, 107–119. [Google Scholar] [CrossRef]
- Shen, D.; Nooraie, F.; Elshimali, Y.; Lonsberry, V.; He, J.; Bose, S.; Chia, D.; Seligson, D.; Chang, H.R.; Goodglick, L. Decreased expression of annexin A1 is correlated with breast cancer development and progression as determined by a tissue microarray analysis. Hum. Pathol. 2006, 37, 1583–1591. [Google Scholar] [CrossRef]
- Alldridge, L.C.; Harris, H.J.; Plevin, R.; Hannon, R.; Bryant, C.E. The annexin protein lipocortin 1 regulates the MAPK/ERK pathway. J. Biol. Chem. 1999, 274, 37620–37628. [Google Scholar]
- Kushner, P.J.; Agard, D.A.; Greene, G.L.; Scanlan, T.S.; Shiau, A.K.; Uht, R.M.; Webb, P. Estrogen receptor pathways to AP-1. J. Steroid Biochem. Mol. Biol. 2000, 74, 311–317. [Google Scholar]
- Parmar, H.; Melov, S.; Samper, E.; Ljung, B.M.; Cunha, G.R.; Benz, C.C. Hyperplasia, reduced E-cadherin expression, and developmental arrest in mammary glands oxidatively stressed by loss of mitochondrial superoxide dismutase. Breast 2005, 14, 256–263. [Google Scholar]
- Mense, S.M.; Remotti, F.; Bhan, A.; Singh, B.; El-Tamer, M.; Hei, T.K.; Bhat, H.K. Estrogen-induced breast cancer: alterations in breast morphology and oxidative stress as a function of estrogen exposure. Toxicol. Appl. Pharmacol. 2008, 232, 78–85. [Google Scholar] [CrossRef]
- Mobley, J.A.; Brueggemeier, R.W. Brueggemeier estrogen receptor-mediated regulation of oxidative stress and DNA damage in breast cancer. Carcinogenesis 2004, 25, 3–9. [Google Scholar]
- Sastre-Serra, J.; Valle, A.; Company, M.M.; Garau, I.; Oliver, J.; Roca, P. Estrogen down-regulates uncoupling proteins and increases oxidative stress in breast cancer. Free Radic. Biol. Med. 2010, 48, 506–512. [Google Scholar] [CrossRef]
- Wójcik, M.; Kosior-Korzecka, U.; Bobowiec, R. Cytoprotective action of 17β-oestradiol against iron-induced hepatic oxidative stress in vitro. Bull Vet. Inst. Pulawy 2010, 54, 259–263. [Google Scholar]
- Kim, S.U.; Hwang, C.N.; Sun, H.N.; Jin, M.H.; Han, Y.H.; Lee, H.; Kim, J.M.; Kim, S.K.; Yu, D.Y.; Lee, D.S.; Lee, S.H. Peroxiredoxin I is an indicator of microglia activation and protects againsthydrogen peroxide-mediatedmicroglial death. Biol. Pharm. Bull. 2008, 31, 820–825. [Google Scholar] [CrossRef]
- Immenschuh, S.; Baumgart-Vogt, E. Peroxiredoxins, oxidative stress, and cell proliferation. Antioxid. Redox Signal. 2005, 7, 768–777. [Google Scholar] [CrossRef]
- Lee, Y.M.; Park, S.H.; Shin, D.I.; Hwang, J.Y.; Park, B.; Park, Y.J.; Lee, T.H.; Chae, H.Z.; Jin, B.K.; Oh, T.H.; Oh, Y.J. Oxidative modification of peroxiredoxin is associated with drug-induced apoptotic signaling in experimental models of Parkinson disease. J. Biol. Chem. 2008, 283, 9986–9998. [Google Scholar]
- Mu, Z.M.; Yin, X.Y.; Prochownik, E.V. Pag, a putative tumor suppressor, interacts with the Myc Box II domain of c-Myc and selectively alters its biological function and target gene expression. J. Biol. Chem. 2002, 277, 43175–43184. [Google Scholar]
- Daly, K.A.; Lefévre, C.; Nicholas, K.; Deane, E.; Williamson, P. Characterization and expression of Peroxiredoxin 1 in the neonatal tammarwallaby (Macropuseugenii). Comp. Biochem. Physiol B Biochem. Mol. Biol. 2008, 49, 108–119. [Google Scholar]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Meiers, I.; Shanks, J.H. Bostwick DG. Glutathione S-transferase pi (GSTP1) hypermethylation in prostate cancer: review 2007. Pathology 2007, 39, 299–304. [Google Scholar] [CrossRef]
- Lee, J.S. GSTP1 promoter hypermethylation is an early event in breast carcinogenesis. Virchows. Arch. 2007, 450, 637–642. [Google Scholar] [CrossRef]
- McIlwain, C.C.; Townsend, D.M.; Tew, K.D. Glutathione S-transferase polymorphisms: cancer incidence and therapy. Oncogene 2006, 25, 1639–1648. [Google Scholar]
- Pasquali, L.; Bedeir, A.; Ringquist, S.; Styche, A.; Bhargava, R.; Trucco, G. Quantification of CpG island methylation in progressive breast lesions from normal to invasive carcinoma. Cancer Lett. 2007, 257, 136–144. [Google Scholar] [CrossRef]
- Parcellier, A.; Schmitt, E.; Brunet, M.; Hammann, A.; Solary, E.; Garrido, C. Small heat shock proteins HSP27 and α B-crystallin: Cytoprotective and oncogenic functions. Antioxid. Redox Signal. 2005, 7, 404–413. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Raman, B.; Ramakrishna, T.; Rao, Ch.M. Effect of phosphorylation on alpha B - crystallin: differences in stability, subunit exchange and chaperone activity of homo and mixed oligomers of alpha B - crystallin and its phosphorylation -mimicking mutant. J. Mol. Biol. 2008, 375, 1040–1051. [Google Scholar] [CrossRef]
- Kamradt, M.C.; Chen, F.; Sam, S.; Cryns, V.L. The small heat shock protein alpha B-crystallin negatively regulates apoptosis during myogenic differentiation by inhibiting caspase-3 activation. J. Biol. Chem. 2002, 277, 38731–38736. [Google Scholar]
- Sitterding, S.M.; Wiseman, W.R.; Schiller, C.L.; Luan, C.; Chen, F.; Moyano, J.V.; Watkin, W.G.; Wiley, E.L.; Cryns, V.L.; Diaz, L.K. AlphaB-crystallin: a novel marker of invasive basal-like and metaplastic breast carcinomas. Ann. Diagn. Pathol. 2008, 12, 33–40. [Google Scholar] [CrossRef]
- Moyano, J.V.; Evans, J.R.; Chen, F.; Lu, M.; Werner, M.E.; Yehiely, F.; Diaz, L.K.; Turbin, D.; Karaca, G.; Wiley, E.; et al. AlphaB- crystallin is a novel oncoprotein that predicts poor clinical outcome in breast cancer. J. Clin. Invest. 2006, 116, 261–270. [Google Scholar]
- Launay, N.; Goudeau, B.; Kato, K.; Vicart, P.; Lilienbaum, A. Cell signaling pathways to alpha B-crystallin following stresses of the cytoskeleton. Exp. Cell. Res. 2006, 312, 3570–3584. [Google Scholar] [CrossRef]
- Polavarapu, R.; Spitz, D.R.; Sim, J.E.; Follansbee, M.H.; Oberley, L.W.; Rahemtulla, A.; Nanji, A.A. Increased lipid peroxidation and impaired antioxidant enzyme function is associated with pathological liver injury in experimental alcoholic liver disease in rats fed diets high in corn oil and fish oil. Hepatology 1998, 27, 1317–1323. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Z.-C.; E, D.; Batu, D.-L.; Saixi, Y.-L.; Zhang, B.; Ren, L.-Q. 2D-DIGE Proteomic Analysis of Changes in Estrogen/Progesterone-Induced Rat Breast Hyperplasia upon Treatment with the Mongolian Remedy RuXian-I. Molecules 2011, 16, 3048-3065. https://doi.org/10.3390/molecules16043048
Wang Z-C, E D, Batu D-L, Saixi Y-L, Zhang B, Ren L-Q. 2D-DIGE Proteomic Analysis of Changes in Estrogen/Progesterone-Induced Rat Breast Hyperplasia upon Treatment with the Mongolian Remedy RuXian-I. Molecules. 2011; 16(4):3048-3065. https://doi.org/10.3390/molecules16043048
Chicago/Turabian StyleWang, Zhong-Chao, Du E, De-Ligen Batu, Ya-Latu Saixi, Bin Zhang, and Li-Qun Ren. 2011. "2D-DIGE Proteomic Analysis of Changes in Estrogen/Progesterone-Induced Rat Breast Hyperplasia upon Treatment with the Mongolian Remedy RuXian-I" Molecules 16, no. 4: 3048-3065. https://doi.org/10.3390/molecules16043048
APA StyleWang, Z.-C., E, D., Batu, D.-L., Saixi, Y.-L., Zhang, B., & Ren, L.-Q. (2011). 2D-DIGE Proteomic Analysis of Changes in Estrogen/Progesterone-Induced Rat Breast Hyperplasia upon Treatment with the Mongolian Remedy RuXian-I. Molecules, 16(4), 3048-3065. https://doi.org/10.3390/molecules16043048