Bioflocculant Production by Virgibacillus sp. Rob Isolated from the Bottom Sediment of Algoa Bay in the Eastern Cape, South Africa


Abstract
:1. Introduction
2. Results and Discussion
2.1. Screening for flocculant producing microorganism

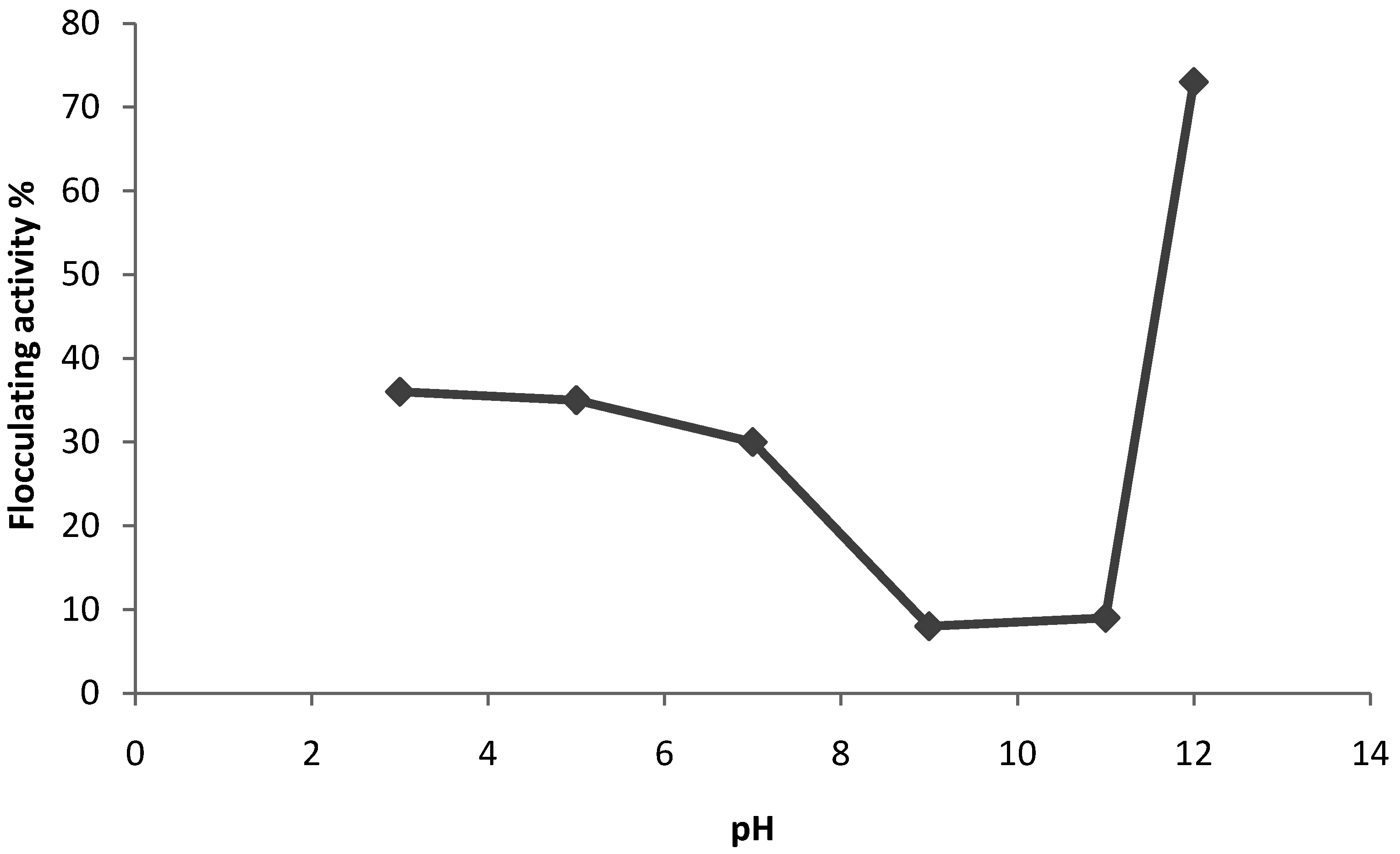
2.2. Factors affecting the bioflocculant production and flocculating activity

{kind=link}
{kind=link}
{kind=link}
| Carbon Source | Glucose | Sucrose | Fructose | Starch |
|---|---|---|---|---|
| Flocculating Activity % | 70.4 | - | 18 | 5 |
| Nitrogen Source | Peptone | Ammonium Sulphate | Urea | Ammonium Chloride |
| Flocculating Activity % | 70.4 | 22 | 38 | 37 |
| Salts | Calcium Chloride | Magnesium Chloride | Iron Sulphate | Potassium Chloride |
| Flocculating Activity % | 70.4 | - | 74 | - |

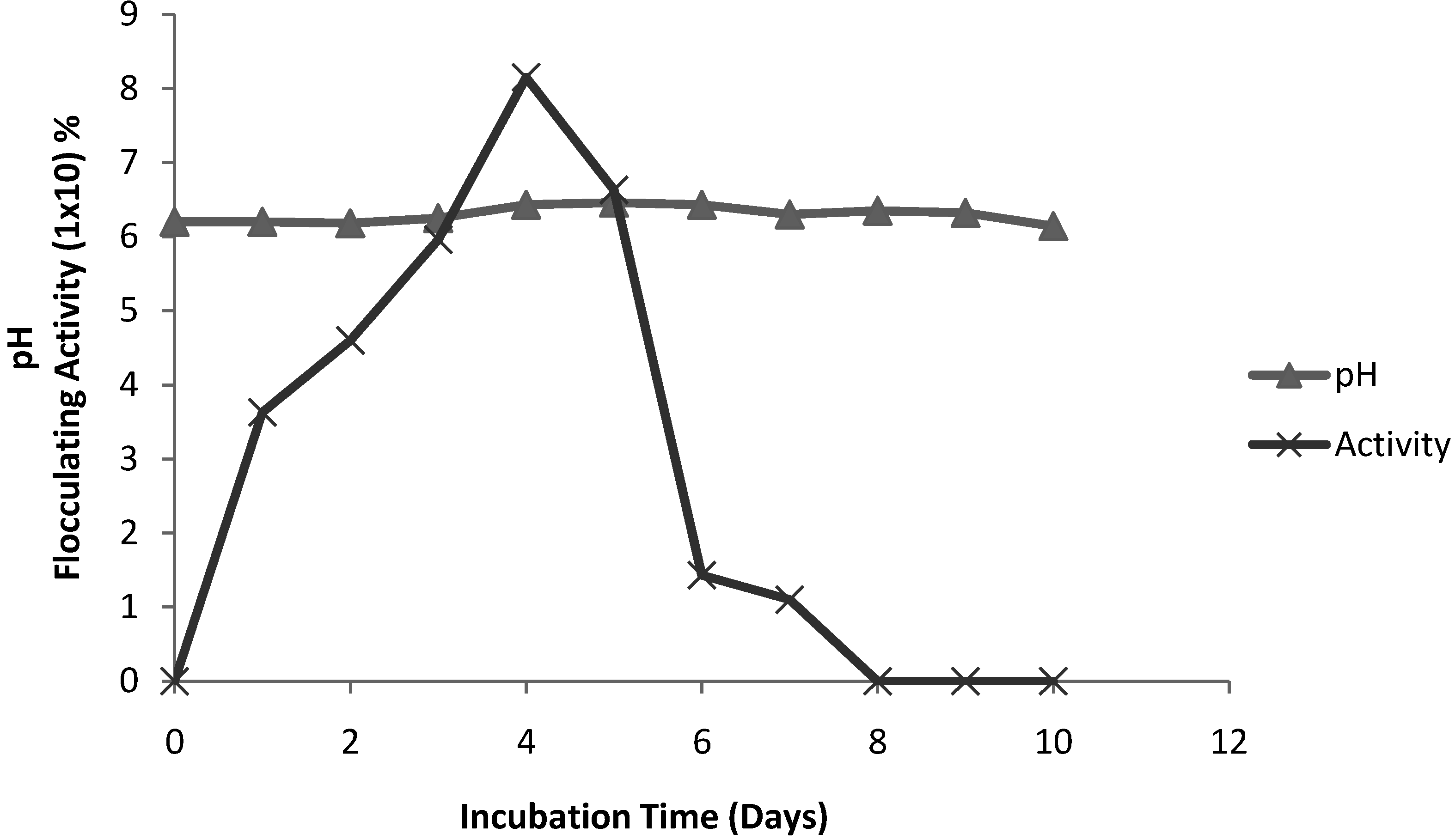
2.3. Time course of bioflocculant production

2.4. Analysis of bioflocculant composition
3. Experimental
3.1. Screening for bioflocculant-producing bacteria
3.2. Determination of flocculating activity
3.3. The effect of carbon and nitrogen sources on bioflocculant production
3.4. Effects of various cations and pH on bioflocculant production
3.5. Time course of bioflocculant production
3.6. Extraction and purification of bioflocculant
3.7. Analysis/ characterization of purified bioflocculant
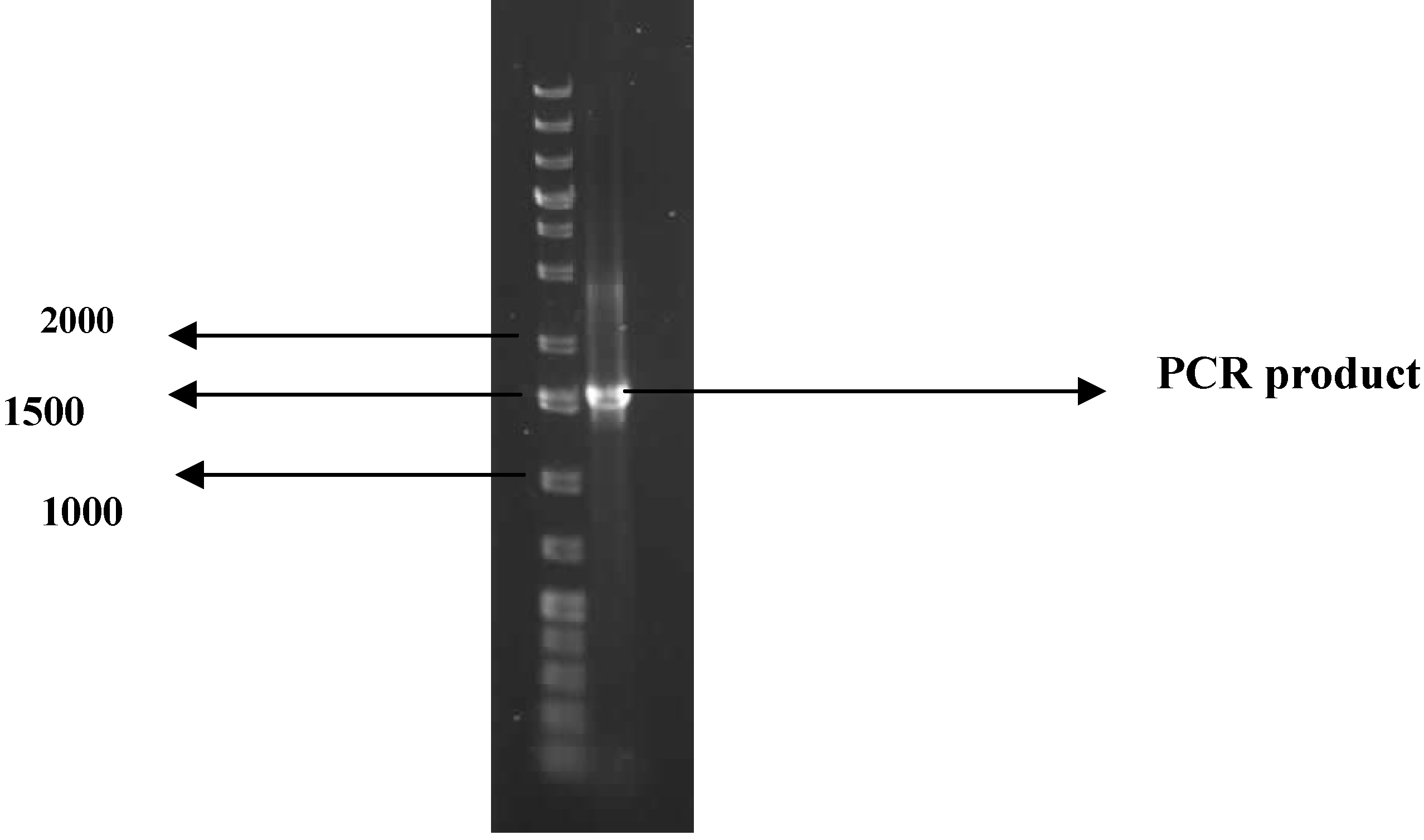
3.8. Identification of the bioflocculant-producing microorganism
4. Conclusions
Acknowledgements
References
- Li, Z.; Zhong, S.; Lei, H.; Chen, R.; Yu, Q.; Li, H. Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresour. Technol. 2009, 100, 3650–3656. [Google Scholar] [CrossRef]
- Salehizadeh, H.; Shojaosadati, S.A. Extracellular biopolymeric flocculants-recent trends and biotechnological importance. Biotechnol. Adv. 2001, 19, 371–385. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Wang, S.; Jiang, P. Characterization of a bioflocculant produced by the marine myxobacteriumNannocystissp. NU2. Appl. Microbiol. Biotechnol. 2002, 59, 517–522. [Google Scholar] [CrossRef]
- Deng, S.; Yu, G.; Ting, Y.P. Production of a bioflocculant by Aspergillusparasiticus and its application in dye removal. Colloid. Surface B 2005, 44, 179–186. [Google Scholar] [CrossRef]
- Zouboulis, A.I.; Chai, X.L.; Katsoyiannis, A.I. The application of bio-flocculant for the removal of humic acids from stabilized landfill leachates. J. Environ. Manage. 2004, 70, 30–35. [Google Scholar]
- Salehizadeh, H.; Shojaosadati, S.A. Removal of metal ions from aqueous solution by polysaccharide produced from Bacillus firmus. Water Res. 2003, 37, 4231–4235. [Google Scholar] [CrossRef]
- Oh, H.M.; Lee, S.J.; Park, M.H. Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp. AM49. Biotechnol. Lett. 2001, 23, 1229–1234. [Google Scholar] [CrossRef]
- He, J.; Zou, J.; Shao, Z.; Zhang, J. Characteristics and flocculating mechanism of a novel bioflocculant HBF-3 produced by deep-sea bacterium mutant Halomona sp. V3a. World J. Microbiol. Biotechnol. 2010, 26, 1135–1141. [Google Scholar] [CrossRef]
- Liu, W.; Yuan, H.; Yang, J.; Li, B. Characterization of bioflocculants from biologically aerated filter backwashed sludge and its application in dying wastewater treatment. Bioresour. Technol. 2009, 100, 2629–2632. [Google Scholar] [CrossRef]
- Lian, B.; Chen, Y.; Zhao, J.; Teng, H.H.; Zhu, L.; Yuan, S. Microbial flocculation by Bacillus mucilaginosus: Applications and mechanisms. Bioresour. Technol. 2007, 99, 4825–4831. [Google Scholar]
- Zheng, Y.; Ye, Z.L.; Fang, X.L.; Li, Y.H.; Cai, W.M. Production and characteristics of a bioflocculant produced by Bacillus sp. F19. Bioresour. Technol. 2008, 99, 7686–7691. [Google Scholar] [CrossRef]
- Kurane, R.; Toeda, K.; Tadeka, K. Culture condition for production of microbial flocculant by Rhodococcuserythropolis. Agric. Biol. Chem. 1986, 50, 2309–2313. [Google Scholar] [CrossRef]
- Yokoi, H.; Yoshida, T.; Mori, S.; Hirose, J.; Hayashi, S.; Takasaki, Y. Biopolymer flocculant produced by Enterobacter sp. Biotechnol. Lett. 1996, 19, 572. [Google Scholar]
- Toeda, K.; Kurane, R. Microbial flocculant from Alcaligenescupidus KT201. Agric. Biol. Chem. 1991, 55, 2793–2799. [Google Scholar] [CrossRef]
- Shih, I.L.; Van, Y.T.; Yeh, L.C.; Lin, H.G.; Chang, Y.N. Production of a biopolymer flocculant from Bacillus licheniformis and its flocculation properties. Bioresour. Technol. 2001, 78, 267–272. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, S.O.; Jang, K.L.; Lee, T.H. Microbial flocculant from Aecuadedron sp. TS-49. Biotechnol. Lett. 1995, 17, 95–100. [Google Scholar] [CrossRef]
- Gao, J.; Bao, H.; Xin, M.; Liu, Y.; Li, Q.; Zhang, Y. Characterization of a bioflocculant from a newly isolated Vagococcus sp. W31. J. Zhejiang Univ. Sci. B. 2006, 7, 186–192. [Google Scholar] [CrossRef]
- He, N.; Li, Y.; Chen, J.; Lun, S.Y. Identification of a novel bioflocculant from a newly isolated Corynebacteriumglutamicum. Biochem. Eng. J. 2002, 11, 137–148. [Google Scholar] [CrossRef]
- Peng, Q.; Chen, J.; Zhang, Y.; Chen, Q.; Peng, D.; Cui, X.; Li, N.; Chen, Y. Virgibacillus zhangjiangensis sp. nov., a marine bacterium isolated from sea water. A. Leeuwhoek. 2009, 96, 645–652. [Google Scholar]
- Heyndrickx, M.; Lebbe, L.; Kersters, K.; De Vos, P.; Forsyth, G.; Logan, N.A. Virgibacillus: A new genus to accommodate Bacillus pantothenticus (Proom, Knight 1950) Emended description of Virgibacilluspantothenticus. Int. J. Syst. Bacteriol. 1998, 48, 99–106. [Google Scholar] [CrossRef]
- Heyrman, J.; Logan, N.A.; Busse, H.J.; Balcaen, A.; Lebbe, L.; Rodriguez-Diaz, M.; Swings, J.; De Vos, P. Virgibacilluscarmonensis sp. nov., Virgibacillus necropolis sp. nov. and Virgibacilluspicturae sp. nov., three novel species isolated from deteriorated mural paintings, transfer of the species of the genus Salibacillus to Virgibacillus, as Virgibacillusmarismortui comb. nov. and Virgibacillussalexigens comb. nov., and emended description of the genus Virgibacillus. Int. J. Syst. Evol. Microbiol. 2003, 53, 501–511. [Google Scholar] [CrossRef]
- Wang, C.; Chang, C.; Ng, C.C.; Chen, T.; Shyu, Y. Virgibacilluschiguensis sp. nov., a novel halophilic bacterium isolated from Chigu, a previously commercial salten located in Southern Taiwan. Int. J. Syst. Evol. Microbiol. 2008, 58, 341–345. [Google Scholar] [CrossRef]
- Gupta, A.; Josheph, B.; Mani, A. Biosynthesis and properties of an extracellular thermostable serine alkaline protease from Virgibacilluspathothenticus. World J. Microbiol. Biotechnol. 2008, 24, 237–243. [Google Scholar] [CrossRef]
- Chamroensaksri, W.; Akaracharanya, A.; Visessanguan, W.; Tanasupawat, S. Characterization of halophilic bacterium nb2-1 from Pla-ra and its protease production. J. Food Biochem. 2008, 32, 536–555. [Google Scholar] [CrossRef]
- Sinsuwan, S.; Rodtong, S.; Yongsawatdigul, J. NaCl-activated extracellular proteinase from Virgibacillus sp. SK37 isolated from fish sauce fermentation. J. Food Sci. 2007, 72, 264–269. [Google Scholar]
- Namwong, S. Isolation, Selection and Identification of Proteinase-produce Bacteria from Fish Sauce Fermentation to be Used as Starter Cultures. Master Thesis, Suranaree University of Technology, Bangkok, Thailand, 2006. [Google Scholar]
- Essghaier, B.; Fardea, M.L.; Cayol, J.L.; Hajlaoui, M.R.; Boudabous, A.; Tijakli, H.; Sadfi-Zouaoui, N. Biological control of grey mould in strawberry fruits by halophilic bacteria. J. Appl. Microbiol. 2009, 106, 833–846. [Google Scholar] [CrossRef]
- He, N.; Li, Y.; Chen, J. Production of a novel polygalacturonic acid bioflocculant REA-11 by Corynebacteriumglutamicum. Bioresour. Technol. 2004, 94, 99–105. [Google Scholar] [CrossRef]
- Xia, S.; Zhang, Z.; Wang, X.; Yang, A.; Chen, L.; Zhao, J.; Leonard, D.; Jaffrezic-Renault, N. Production and characterization of bioflocculant by Proteus mirabilis TJ-1. Bioresour. Technol. 2008, 99, 6520–6527. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhang, Q.; Sheng, Y.; Li, C.; Wang, H. Screening and flocculating properties of bioflocculant-producing microorganisms. Sci. Technol. 2006, 13, 289–292. [Google Scholar]
- Gong, W.; Wang, S.; Sun, F.; Liu, X.W.; Yue, Q.Y.; Gao, B.Y. Bioflocculant production by culture of Serratiaficaria and its application in wastewater treatment. Bioresour. Technol. 2008, 99, 4668–4674. [Google Scholar] [CrossRef]
- Lachhwani, P. Studies on Polymeric Bioflocculant Producing Microorganisms. Master dissertation, Thapar Institute of Engineering and Technology, Patiala, India, 2005. [Google Scholar]
- Kurane, R.; Nohata, Y. A new water-absorbing polysaccharide from Alcaligeneslatus. Biosci. Biotechnol. Biochem. 1994, 58, 236. [Google Scholar]
- Li, X.M.; Haung, Q.Y.; Zeng, G.M.; Liao, X.; Liu, J.J.; Long, W.F. Screening and characterization of a bioflocculant produced by Aeromonas sp. Biomed. Environ. Sci. 2007, 20, 274–278. [Google Scholar]
- Li, W.W.; Zhou, W.Z.; Zhang, Y.Z.; Wang, J.; Zhu, X.B. Flocculation behavior and mechanism of an exopolysaccharide from the deep-sea psychrophilic bacterium Pseudoalteromonas sp. SM9913. Bioresour. Technol. 2008, 99, 6893–6899. [Google Scholar] [CrossRef]
- Shimofuruya, H.; Koide, A.; Shirota, K.; Tsuji, T.; Nakamura, M.; Suzuki, J. The production of flocculating substance (s) by Streptomyces griseus. Biosci. Biotechnol. Biochem. 1995, 60, 498–500. [Google Scholar]
- Kurane, R.; Hatamochi, K.; Kiyohara, T.; Hirao, M.; Taniguchi, Y. Production of a bioflocculant by Rhodococcuserythropolis S-1 grown on alcohols. Biosci. Biotechnol. Biochem. 1994, 58, 428–429. [Google Scholar] [CrossRef]
- Takadi, H.; Kodowaki, K. Flocculant production by Paecilomyces sp. Taxanomic studies and culture conditions for production. Agric. Biol. Chem. 1985, 49, 3151–3157. [Google Scholar] [CrossRef]
- Fugita, M.; Ike, M.; Jang, J.H.; Kim, S.M.; Hirao, T. Bioflocculant production from lower-molecular fatty acids as a novel strategy for utilization of sludge digestion liquor. World Sci. Technol. 2001, 44, 237–243. [Google Scholar]
- Zhang, Z.; Lin, B.; Xia, S.; Wang, X.; Yang, A. Production and application of a novel bioflocculant by multi-microorganism consortia using brewery wastewater as carbon source. J. Environ. Sci. 2007, 19, 667–673. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, S.J.; Ahn, S.H.; Lee, H.K. Characterization of novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodiumimpudicum KG03. Bioresour. Technol. 2006, 98, 361–367. [Google Scholar]
- Chang, W.C.; Soon, A.Y.; In, H.O.; Sang, H.P. Characterization of an extracellular flocculating substance produced by a planktonic cyanobacterium, Anabaena sp. Biotechnol. Lett. 1998, 20, 643–646. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, J.F.; Jiang, P.J.; Yang, S.L.; Liu, Z.L. Composition and characterization of microbiological flocculant SC06. Environ. Chem. 2002, 21, 360–364. [Google Scholar]
- Cook, A.E.; Meyers, P.R. Rapid identification of filamentous actinomycetes to the genus level using genus-specific 16S rRNA gene restriction fragment patterns. Int. J. Syst. Evol. Microbiol. 2003, 53, 1907–1915. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cosa, S.; Mabinya, L.V.; Olaniran, A.O.; Okoh, O.O.; Bernard, K.; Deyzel, S.; Okoh, A.I. Bioflocculant Production by Virgibacillus sp. Rob Isolated from the Bottom Sediment of Algoa Bay in the Eastern Cape, South Africa. Molecules 2011, 16, 2431-2442. https://doi.org/10.3390/molecules16032431
Cosa S, Mabinya LV, Olaniran AO, Okoh OO, Bernard K, Deyzel S, Okoh AI. Bioflocculant Production by Virgibacillus sp. Rob Isolated from the Bottom Sediment of Algoa Bay in the Eastern Cape, South Africa. Molecules. 2011; 16(3):2431-2442. https://doi.org/10.3390/molecules16032431
Chicago/Turabian StyleCosa, Sekelwa, Leonard V. Mabinya, Ademola O. Olaniran, Omobola O. Okoh, Kim Bernard, Shaun Deyzel, and Anthony I. Okoh. 2011. "Bioflocculant Production by Virgibacillus sp. Rob Isolated from the Bottom Sediment of Algoa Bay in the Eastern Cape, South Africa" Molecules 16, no. 3: 2431-2442. https://doi.org/10.3390/molecules16032431
APA StyleCosa, S., Mabinya, L. V., Olaniran, A. O., Okoh, O. O., Bernard, K., Deyzel, S., & Okoh, A. I. (2011). Bioflocculant Production by Virgibacillus sp. Rob Isolated from the Bottom Sediment of Algoa Bay in the Eastern Cape, South Africa. Molecules, 16(3), 2431-2442. https://doi.org/10.3390/molecules16032431