The Triterpenes 3β-Lup-20(29)-en-3-ol and 3β-Lup-20(29)-en-3-yl Acetate and the Carbohydrate 1,2,3,4,5,6-Hexa-O-acetyl-dulcitol as Photosynthesis Light Reactions Inhibitors

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Lupeol (1), 3β-Lup-20(29)-en-3-yl Acetate (2) and 1,2,3,4,5,6-Hexa-O-acetyl-dulcitol (3) on Photophosphorylation

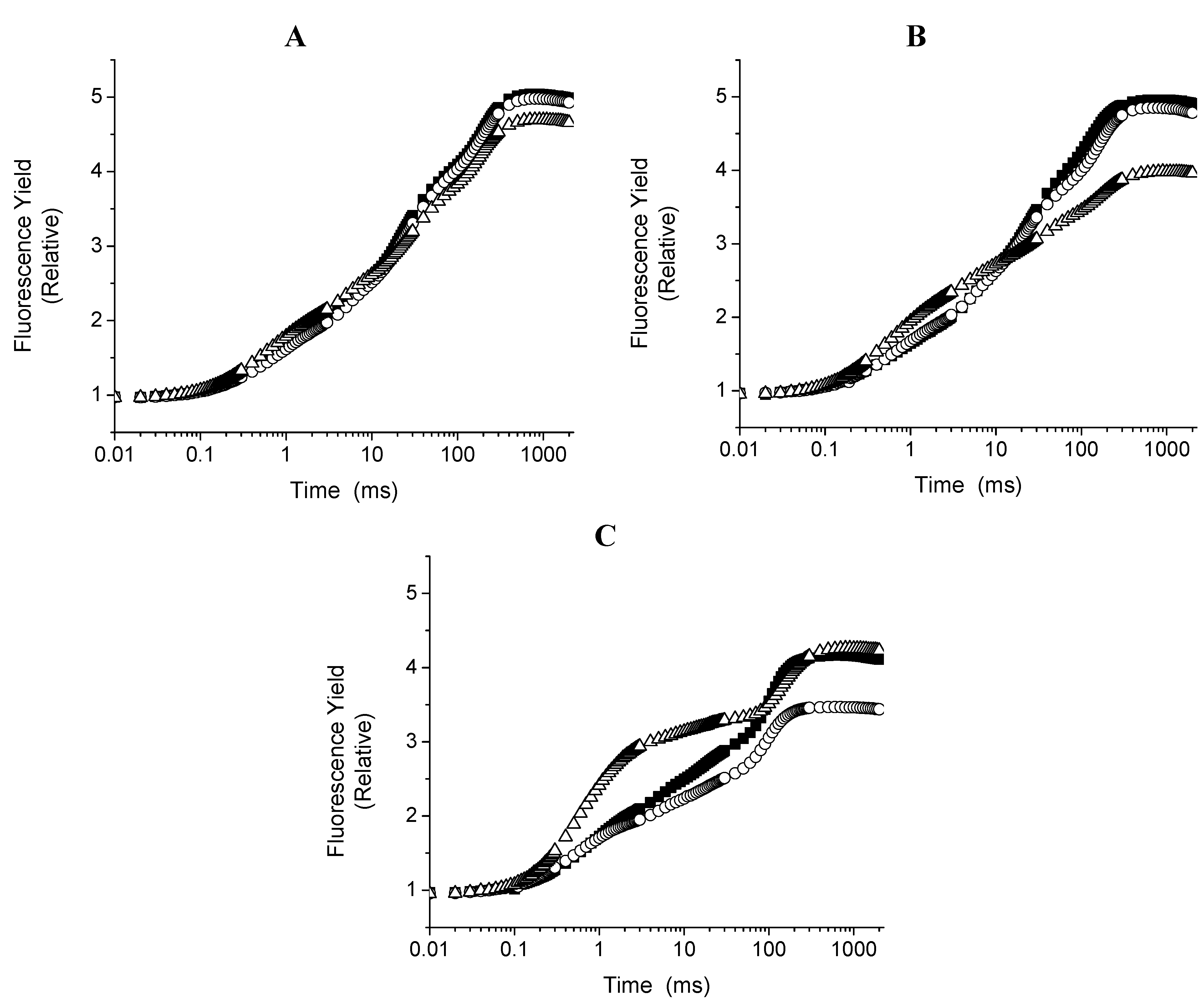
2.2. The Effect of Compounds 1 and 3 on Different Photosynthetic Activities

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Conc. (µM) | Basal | Phosphorylating | Uncoupled | |||
|---|---|---|---|---|---|---|---|
| a | b | a | b | a | b | ||
| 1 | 0 | 624 | 100 | 835 | 100 | 930 | 100 |
| 300 | 499 | 80 | 710 | 85 | 930 | 100 | |
| 7 | 0 | 334 | 100 | 667 | 100 | 1480 | 100 |
| 300 | 0 | 0 | 180 | 27 | 1401 | 95 | |
| Lup-20(29)-in-3β-ol [μM] | Mg2+-ATPase (%) | Ca2+-ATPase (%) |
|---|---|---|
| Control | 100 | 100 |
| 100 | 65 | 182 |
| 200 | 86 | 178.3 |
| 300 | 98 | 178.8 |

2.3. Chl a Fluorescence Measurements in Spinach Leaf Discs with and without Compounds 1 and 3



2.4. In Vivo Assays


3. Experimental
3.1. General
3.1.1. Reagents
| Adenosine diphosphate | Adenosine triphosphate | Ascorbic acid |
| CaCl2 | Chloroform | Ethyl acetate |
| 2,5-Dibromo-6-isopropyl-3-methyl-1,4-benzoquinone | 2,5-Dichloro-1,4-benzoquinone | 2,6-Dichlorophenolindophenol |
| 3-(3,4-Dichlorophenyl)-1,1-dimethylurea | Dimethyl sulfoxide (DMSO) | Dithiothreitol (DTT) |
| Ethylenediaminetetraacetic acid (EDTA) | n-Hexane | N-[2-Hydroxyethyl]piperazine-N′-[2-ethanesulfonic acid] |
| Isopropyl alcohol | KCl | KOH |
| MgCl2 | Methanol | Methylviologen |
| NaCl | NaHCO3 | Na2SO4 |
| 2-(N-Morpholino)-ethanesulfonic acid | Polyoxyethylensorbitan monolaurate (Tween-20) | Silica-gel 60-230 mesh |
| Silica-gel HF 60 | Sodium silicomolybdate (SiMo) | Sorbitol |
| Sucrose | Trichloacetic acid (TCA) | Tricine |
3.1.2. Equipment
| Bruker DRX 400 (or 200) spectrometer for NMR experiments |
| Corning potentiometer model 12 (Corning Medical, Acton, MA, USA) |
| Gilson recorder (Kipp & Zonen, Bohemia, NY, USA) |
| Hansatech Fluorescence Handy PEA (plant efficiency analyzer) |
| Oxygraph YSI (Yellow Spring Instrument) Model 5300 |
| Shimadzu QP-5050A Gas chromatography/Mass Spectrometry GC/MS, equipped with a PTE™-5 column (30 m, 0.25 mm, 0.25 µm, Supelco, USA) |
| Shimadzu QP5050A device for electron impact (70 eV) low-resolution mass spectra (EI-MS) |
3.1.3. Methods
3.1.3.1. Plant Material
3.1.3.2. Extraction and Isolation from M. acanthophylla
3.1.3.3. Extraction and Isolation from Xylosma flexuosa
3.1.3.4. Chloroplast Isolation and Chlorophyll Determination
3.1.3.5. ATP Synthesis and Electron Flow Determinations
3.1.3.6. Measurements of Non-Cyclic Electron Transport Rate
3.1.3.7. Mg2+-ATPase Activity Assays
3.1.3.8. Ca2+-ATPase Activity Assays
3.1.3.9. Chlorophyll a Fluorescence Measurements in Spinach Leaf Discs
3.1.3.10. Plant Material for in Vivo Assays
3.1.3.11. Chlorophyll a Fluorescence Determination in Intact Leaves in Vivo
4. Conclusions
Acknowledgments
References and Notes
- Carvalho-Okano, R.M. Estudos taxonômicos do gênero Maytenus Mol: emend. Mol. (Celastraceae) do Brasil Extra-Amazônico. Ph.D. Thesis,, Instituto de Biologia, UNICAMP, 1992; p. 252. [Google Scholar]
- Biodiversitas Revisăo das listas das espécies da flora e da fauna ameaçadas de extinção do estado de Minas gerais – relatório final; Fundação bBiodiversitas: Belo Horizonte, Brazil, 2007; Volume 2.
- de Oliveira, D.M.; Silva, G.; Duarte, L.P.; Vieira, A.S. Chemical constituents isolated from roots of Maytenus acanthophylla Reissek (Celastraceae). Biochem. System. Ecol. 2006, 34, 661–665. [Google Scholar] [CrossRef]
- Morales, F.; Fonseca, R.M.; Diego, P. Flora de Guerrero. Rev de la Fac de Ciencias, UNAM, México. 2002, 9, 109–110. [Google Scholar]
- Gibbons, S.; Gray, A.I.; Waterman, P.G.; Hockless, D.C.R.; Skelton, B.W.; White, A.H. Beonxoylated derivatives od 2-neta.glucopyranosyloxy-2,5-dihydroxybenzyl alcohol from Xylosma flexuosum: Structure and relative configuration of xylosmin. J. Nat. Prod. 1995, 58, 554–559. [Google Scholar] [CrossRef]
- Macias, F.A.; Molinillo, J.M.G.; Galindo, J.C.G.; Varela, R.M.; Simonet, A.M. Castellano, D. The use of allelophatic Studies in the search for natural herbicides. J. Crop Prod. 2001, 4, 237–255. [Google Scholar] [CrossRef]
- Macias, F.A.; Molinillo, J.M.G.; Galindo, J.C.G.; Varela, R.M.; Torres, A.; Simonet, A.M. Terpenoids with potencial use as natural herbicide templates. In Biologically Active Natural Products: Agrochemicals; Cutler, H.G., Cutler, S.J., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 15–31. [Google Scholar]
- King-Díaz, B.; Castelo-Branco, P.A.; Leite dos Santos, F.J.; Rubinger, M.M.M.; Ferreira-Alves, D.L.; Piló-Veloso, D.; Lotina-Hennsen, B. Furanoditerpene ester and thiocarbonyldioxy derivatives inhibit photosynthesis. Pestic. Biochem. Physiol. 2010, 96, 119–126. [Google Scholar] [CrossRef]
- Torres-Romero, D.; King-Díaz, B.; Strasser, R.J.; Jiménez, I.A.; Lotina-Hennsen, B.; Bazzocchi, I.L. Friedelane triterpenes from Celastrus vulcanicola as Photosynthetic Inhibitors. J. Agric. Food Chem. 2010, 58, 10847–10854. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll a fluorescence transients in plants and cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Vázquez, R.; King-Díaz, B.; Aguilar, M.I.; Diego, N.; Lotina-Hennsen, B. Pachypodol from Croton ciliatoglanduliferus Ort as water-splitting enzyme inhibitor on thylakoids. J. Agric. Food Chem. 2006, 54, 1217–1221. [Google Scholar] [CrossRef]
- McCarthy, R.E. Energy Transfer inhibitors of photophosphorylation in chloroplasts. In Encyclopedia of Plant Physiology New Series, Chapter 12. Photosynthesis; Trebst, A., Avron, M., Eds.; Springer-Verlag: Berlin, Germany, 1977; Volume 5, pp. 437–447. [Google Scholar]
- Srivastava, A.; Guisse, B.; Greppin, H.; Strasser, R.J. Regulation of antenna structure and electron transport in PSII of Pisumsativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient OKJIP. Biochim. Biophys. Acta 1320, 95–106. [Google Scholar]
- Guisse, B.; Srivastava, A.; Strasser, R.J. The polyphasic rise of the chlorophyll a fluorescence (O–K–J–I–P) in heat stressed leaves. Arch. Sci. Geneve 1995, 48, 147–160. [Google Scholar]
- Morales-Flores, F.; Aguilar, M.I.; King-Díaz, B.; de Santiago-Gómez, J.-R.; Lotina-Hennsen, B. Natural diterpenes from Croton ciliatoglanduliferus as photosystem II and photosystem I inhibitors as spinach chloroplasts. Photosynth. Res. 2007, 91, 71–80. [Google Scholar] [CrossRef]
- González-Bernardo, E.; Aguilar, M.I.; Delgado, G.; King-Díaz, B.; Lotina-Hennsen, B. Photosynthetic electron transport interaction of xanthorrhizol isolated from Iostephane heterophylla and its derivatives. Physiol. Plant. 2003, 119, 598–604. [Google Scholar] [CrossRef]
- Oukarroum, A.; El Madidi, S.; Strasser, R.J. Drougth stress induced in barley cultivars (Hordeum vulgare L.) by polyethylene glycol, probed by the fast fluorescence rise OJIP, germination and root length. Arch. Sci. Genève 2006, 59, 20–21. [Google Scholar]
- Strasser, B.J. Donor side capacity of Photosystem II probed by chlorophyll a fluorescence transients. Photosyn.Res. 1997, 52, 147–155. [Google Scholar] [CrossRef]
- Céspedes, C.L.; Calderón, J.S.; King-Díaz, B.; Lotina-Hennsen, B. Phytochemical and biochemical characterization of epimeric photogedunin derivatives. Their different sites of interaction on the redox electron transport carrier of Spinaceae oleracea L. chloroplasts. J. Agric. Food Chem. 1998, 46, 2810–2816. [Google Scholar]
- Burns, D.; Reynolds, W.F.; Buchanan, G.; Reese, P.B.; Enriquez, R.G. Assignment of spectra and investigation of hindered side-chain rotation in lupeol derivatives. Magn. Reson. Chem. 2000, 38, 488–493. [Google Scholar]
- Silva, J.R.D.A.; Rezende, C.M.; Pinto, A.C.; Pinheiro, M.L.B.; Cordeiro, M.C.; Tamborini, E.; Young, C.; Bolzani, V.D.S. Ésteres triterpénicos de Himatanthus succuba (Spruce) Woodson. Quím. Nova 1998, 21, 702–704. [Google Scholar] [CrossRef]
- Angyal, S.J.; Fur, R.L. The 13C-NMR espectra of alditols. Carbohyd. Res. 1980, 84, 201–209. [Google Scholar] [CrossRef]
- Atta-ur, R. Handbook of Natural Products. Pentacyclic Triterpenoids; Elsevier: Amsterdam, The Netherlands, 1994; pp. 1038–1040. [Google Scholar]
- Strain, H.H.; Cope, B.T.; Svec, W.A. Analytical procedures for the isolation, identification, estimation and investigation of the chlorophylls. Meth. Enzymol. 1971, 23, 452–276. [Google Scholar] [CrossRef]
- Dilley, R.A. Ion transport (H+, K+, Mg2+ exchange phenomena). Methods. Enzymol. 1972, 24, 68–74. [Google Scholar] [CrossRef]
- Mills, J.D.; Mitchell, P.; Schurrmann, P. Modulation of coupling ATPase activity in intact chloroplasts. FEBS Lett. 1980, 112, 173–177. [Google Scholar] [CrossRef]
- Sumner, J.B. Scientific apparatus and laboratory methods. A method for the colorimetric determination of phosphorus. Science 1944, 100, 413–418. [Google Scholar]
- Aguilar, M.I.; Romero, M.G.; Chávez, M.I.; King-Díaz, B.; Lotina-Hennsen, B. Biflavonoids isolated from Selaginella lepidophylla inhibit photosynthesis in spinach chloroplasts. J. Agric. Food Chem. 2008, 56, 6994–7000. [Google Scholar]
- Panda, D.; Rao, D.N.; Sharma, S.G.; Strasser, R.J.; Sarkar, R.K. Subemergence effects on rice genotypes during seedling stage: Probing of subemergence driven changes of photosystem II by chlorophyll a fluorescence induction O-J-I-P transients. Photosynthetica 2006, 44, 69–75. [Google Scholar] [CrossRef]
- Jiang, H.-X.; Chen, L.-S.; Zheeng, J.-G.; Han, S.; Tang, N.; Smith, B.R. Aluminum-induced effects on photosystem II photochemistry in Citrus leaves assessed by the chlorophyll a fluorescence transient. Tree Physiol. 2008, 28, 1863–1871. [Google Scholar] [CrossRef]
- González-Ibarra, M.; Farfán, N.; Trejo, C.; Uribe, S.; Lotina-Hennsen, B. Selective herbicide activity of 2,5-di(benzylamine)-p-benzoquinone against the monocot weed Echinochloa crusgalli. An in vivo analysis of photosynthesis and growth. J. Agric. Food Chem. 2005, 53, 3415–3420. [Google Scholar]
- Samples Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Menezes-de-Oliveira, D.; Aguilar, M.-I.; King-Díaz, B.; Vieira-Filho, S.A.; Pains-Duarte, L.; Silva, G.-D.d.F.; Lotina-Hennsen, B. The Triterpenes 3β-Lup-20(29)-en-3-ol and 3β-Lup-20(29)-en-3-yl Acetate and the Carbohydrate 1,2,3,4,5,6-Hexa-O-acetyl-dulcitol as Photosynthesis Light Reactions Inhibitors. Molecules 2011, 16, 9939-9956. https://doi.org/10.3390/molecules16129939
Menezes-de-Oliveira D, Aguilar M-I, King-Díaz B, Vieira-Filho SA, Pains-Duarte L, Silva G-DdF, Lotina-Hennsen B. The Triterpenes 3β-Lup-20(29)-en-3-ol and 3β-Lup-20(29)-en-3-yl Acetate and the Carbohydrate 1,2,3,4,5,6-Hexa-O-acetyl-dulcitol as Photosynthesis Light Reactions Inhibitors. Molecules. 2011; 16(12):9939-9956. https://doi.org/10.3390/molecules16129939
Chicago/Turabian StyleMenezes-de-Oliveira, Djalma, Maria-Isabel Aguilar, Beatriz King-Díaz, Sidney Augusto Vieira-Filho, Lucinier Pains-Duarte, Grácia-Divina de Fátima Silva, and Blas Lotina-Hennsen. 2011. "The Triterpenes 3β-Lup-20(29)-en-3-ol and 3β-Lup-20(29)-en-3-yl Acetate and the Carbohydrate 1,2,3,4,5,6-Hexa-O-acetyl-dulcitol as Photosynthesis Light Reactions Inhibitors" Molecules 16, no. 12: 9939-9956. https://doi.org/10.3390/molecules16129939