Icariside II Enhances Nrf2 Nuclear Translocation to Upregulate Phase II Detoxifying Enzyme Expression Coupled with the ERK, Akt and JNK Signaling Pathways

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Abbreviation
| GSH | glutathione |
| HO-1 | heme oxygenase-1 |
| GST | glutathione S-transferase |
| ARE | antioxidant responsive element |
| Keap1 | Kelch-like ECH-associated protein 1 |
| GCL | g-glutamyl-cysteine ligase |
| PKC | Protein kinase C |
| ER | endoplasmic reticulum |
| VEGF | vascular endothelial growth factor |
| Glut4 | glucose transporter 4 |
1. Introduction

2. Results and Discussion
2.1. Cytotoxicity Assays

2.2. Icariside II Increased Nrf2 Nuclear Translocation

2.3. Icariside II Induce Nrf2-Related Protein Expression

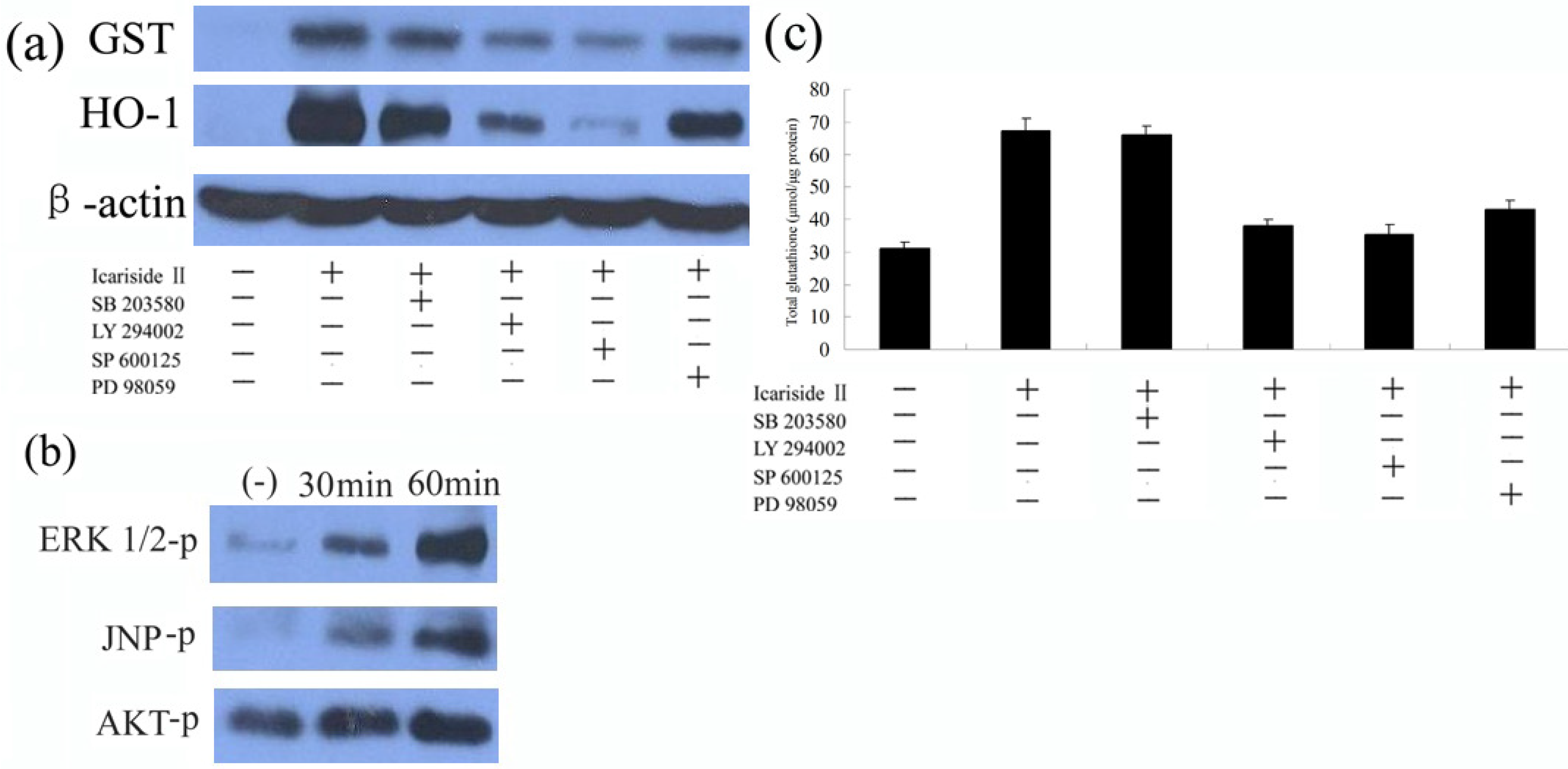
2.4. ERK, AKT, and JNK Signaling Pathways Participate in the Icariside II-Induced Phase II Protein Expression

3. Experimental
3.1. Cell Culture
3.2. Chemicals and Antibodies
3.3. Viability Assay
3.4. Determination of Intracellular GSH Levels
3.5. Western-Blot Analysis
3.6. Statistical Analyses
4. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Chen, C.; Kong, A.N. Dietary cancer-chemopreventive compounds: From signaling and gene expression to pharmacological effects. Trends Pharmacol. Sci. 2005, 26, 318–326. [Google Scholar] [CrossRef]
- Kong, A.N. Signal transduction in cancer chemoprevention. Mutat. Res. 2004, 555, 1–2. [Google Scholar] [CrossRef]
- Yu, X.; Kensler, T. Nrf2 as a target for cancer chemoprevention. Mutat. Res. 2005, 591, 93–102. [Google Scholar] [CrossRef]
- Kong, A.N.; Owuor, E.; Yu, R. Induction of xenobiotic enzymes by the MAP kinase pathway and the antioxidant or electrophile response element (ARE/EpRE). Drug Metab. Rev. 2001, 33, 255–271. [Google Scholar] [CrossRef]
- Keum, Y.S.; Owuor, E.D.; Kim, B.R.; Hu, R.; Kong, A.N. Involvement of Nrf2 and JNK1 in the activation of antioxidant responsive element (ARE) by chemopreventive agent phenethyl isothiocyanate (PEITC). Pharm. Res. 2003, 20, 1351–1356. [Google Scholar] [CrossRef]
- Rushmore, T.H.; Kong, A.N. Pharmacogenomics, regulation and signaling pathways of phase I and II drug metabolizing enzymes. Curr. Drug Metab. 2002, 3, 481–490. [Google Scholar] [CrossRef]
- Chen, C.; Pung, D.; Leong, V. Induction of detoxifying enzymes by garlic organosulfur compounds through transcription factor Nrf2: Effect of chemical structure and stress signals. Free Radic. Biol. Med. 2004, 37, 1578–1590. [Google Scholar] [CrossRef]
- Hu, R.; Hebbar, V.; Kim, B.R. In vivo pharmacokinetics and regulation of gene expression profiles by isothiocyanate sulforaphane in the rat. J. Pharmacol. Exp. Ther. 2004, 310, 263–271. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef]
- Forman, H.J.; Ruden, D. Introduction to serial reviews on EpRE and its signaling pathway. Free Radic. Biol. Med. 2004, 36, 1197–1198. [Google Scholar] [CrossRef]
- Jaiswal, A.K. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic. Biol. Med. 2004, 36, 1199–1207. [Google Scholar] [CrossRef]
- Itoh, K.; Tong, K.I.; Yamamoto, M. Molecular mechanism activating Nrf2-Keap1 pathway in regulation of adaptive response to electrophiles. Free Radic. Biol. Med. 2004, 36, 1208–1213. [Google Scholar] [CrossRef]
- Nguyen, T.; Yang, C.S.; Pickett, C.B. The pathways and molecular mechanisms regulating Nrf2 activation in response to chemical stress. Free Radic. Biol. Med. 2004, 37, 433–441. [Google Scholar] [CrossRef]
- Chen, C.; Yu, R.; Owuor, E.D.; Kong, A.N. Activation of antioxidant-response element (ARE), mitogen-activated protein kinases (MAPKs) and caspases by major green tea polyphenol components during cell survival and death. Arch. Pharm. Res. 2000, 23, 605–612. [Google Scholar] [CrossRef]
- Shen, G.; Hebbar, V.; Nair, S. Regulation of Nrf2 transactivation domain activity. The differential effects of mitogen-activated protein kinase cascades and synergistic stimulatory effect of Raf and CREB-binding protein. J. Biol. Chem. 2004, 279, 23052–23060. [Google Scholar] [CrossRef]
- Yu, R.; Lei, W.; Mandlekar, S. Role of a mitogen-activated protein kinase pathway in the induction of phase II detoxifying enzymes by chemicals. J. Biol. Chem. 1999, 274, 27545–27552. [Google Scholar] [CrossRef]
- Yu, R.; Mandlekar, S.; Lei, W.; Fahl, W.E.; Tan, T.H.; Kong, A.T. p38 mitogen-activated protein kinase negatively regulates the induction of phase II drug-metabolizing enzymes that detoxify carcinogens. J. Biol. Chem. 2000, 275, 2322–2327. [Google Scholar]
- Kang, K.W.; Lee, S.J.; Park, J.W.; Kim, S.G. Phosphatidylinositol 3-kinase regulates nuclear translocation of NF-E2-related factor 2 through actin rearrangement in response to oxidative stress. Mol. Pharmacol. 2002, 62, 1001–1010. [Google Scholar] [CrossRef]
- Huang, H.C.; Nguyen, T.; Pickett, C.B. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. J. Biol. Chem. 2002, 277, 42769–42774. [Google Scholar] [CrossRef]
- Cullinan, S.B.; Zhang, D.; Hannink, M.; Arvisais, E.; Kaufman, R.J.; Diehl, J.A. Nrf2 is a direct PERK substrate and effector of PERK-dependent cell survival. Mol. Cell. Biol. 2003, 23, 7198–7209. [Google Scholar] [CrossRef]
- Cullinan, S.B.; Diehl, J.A. PERK-dependent activation of Nrf2 contributes to redox homeostasis and cell survival following endoplasmic reticulum stress. J. Biol. Chem. 2004, 279, 20108–20117. [Google Scholar] [CrossRef]
- Kee, C.H. The Pharmacology of Chinese Herbs, 2nd ed; CRC Press: Boca Raton, FL, USA, 2000; pp. 106–107. [Google Scholar]
- Oh, M.H.; Houghton, P.J.; Whang, W.K.; Cho, J.H. Screening of Korean herbal medicines used to improve cognitive function for anti-cholinesterase activity. Phytomedicine 2004, 11, 544–548. [Google Scholar] [CrossRef]
- Meng, F.H.; Li, Y.B.; Xiong, Z.L.; Jiang, Z.M.; Li, F.M. Osteoblastic proliferative activity of Epimedium brevicornum Maxim. Phytomedicine 2005, 12, 189–193. [Google Scholar] [CrossRef]
- Li, W.K.; Zhang, R.Y.; Xiao, P.G. Flavonoids from Epimedium wanshanense. Phytochemistry 1996, 43, 527–530. [Google Scholar] [CrossRef]
- Lee, K.S.; Lee, H.J.; Kwang, S.A.; Kim, S.H.; Nam, D.; Kim, D.K.; Choi, D.Y.; Ahn, K.S.; Lu, J.; Kim, S.H. Cyclooxygenase-2/prostaglandin E 2 pathway mediates icariside II induced apoptosis in human PC-3 prostate cancer cells. Cancer Lett. 2009, 280, 93–100. [Google Scholar] [CrossRef]
- Kureishi, Y.; Luo, Z.; Shiojima, I. The HMG-CoA reductase inhibitor simvastatin activates the protein kinase Akt and promotes angiogenesis in normocholes-terolemic animals. Nat. Med. 2000, 6, 1004–1010. [Google Scholar] [CrossRef]
- Miyashita, Y.; Ozaki, H.; Koide, N. Oxysterol-induced apoptosis of vascular smooth muscle cells is reduced by HMG-CoA reductase inhibitor, pravastatin. J. Atheroscler. Thromb. 2002, 9, 65–71. [Google Scholar] [CrossRef]
- Son, B.K.; Kozaki, K.; Iijima, K. Statins protect human aortic smooth muscle cells from inorganic phosphate-induced calcification by restoring Gas6-Axl survival pathway. Circ. Res. 2006, 98, 1024–1031. [Google Scholar] [CrossRef]
- Guijarro, C.; Blanco-Colio, L.M.; Ortego, M. 3-Hydroxy-3 methylglutaryl coenzyme A reductase and isoprenylation inhibitors induce apoptosis of vascular smooth muscle cells in culture. Circ. Res. 1998, 83, 490–500. [Google Scholar] [CrossRef]
- Lee, T.D.; Yang, H.; Whang, J.; Lu, S.C. Cloning and characterization of the human glutathione synthetase 5′-flanking region. Biochem. J. 2005, 390, 521–528. [Google Scholar] [CrossRef]
- Pi, J.; Bai, Y.; Reece, J.M.; Williams, J.; Liu, D.; Freeman, M.L.; Fahl, W.E.; Shugar, D.; Liu, J.; Qu, W.; Collins, S.; Waalkes, M.P. Molecular mechanism of human Nrf2 activation and degradation: role of sequential phosphorylation by protein kinase CK2. Free Radic. Biol. Med. 2007, 42, 1797–1806. [Google Scholar] [CrossRef]
- Nguyen, T.; Yang, C.S.; Pickett, C.B. The pathways and molecular mechanisms regulating Nrf2 activation in response to chemical stress. Free Radic. Biol. Med. 2004, 37, 433–441. [Google Scholar] [CrossRef]
- Li, J.; Ichikawa, T.; Janicki, J.S.; Cui, T. Targeting the Nrf2 pathway against cardiovascular disease. Expert Opin. Ther. Targets 2009, 13, 785–794. [Google Scholar] [CrossRef]
- Warabi, E.; Takabe, W.; Minami, T. Shear stress stabilizes NF-E2-related factor 2 and induces antioxidant genes in endothelial cells: Role of reactive oxygen/nitrogen species. Free Radic. Biol. Med. 2007, 42, 260–269. [Google Scholar] [CrossRef]
- Haridas, M.; Hanausek, G.; Nishimura, H.; Soehnge, W.; Zbigniew, M.; Narog, E.; Spears, R.; Gutterman, J.U. Triterpenoid electrophiles (avicins) activate innate stress response by redox regulation of a gene battery. J. Clin. Invest. 2004, 113, 65–73. [Google Scholar]
- Wasserman, W.W.; Fahl, W.E. Functional antioxidant responsive elements. Proc. Natl. Acad. Sci. USA 1997, 94, 5361–5366. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gu, J.; Sun, X.; Wang, G.; Li, M.; Chi, M. Icariside II Enhances Nrf2 Nuclear Translocation to Upregulate Phase II Detoxifying Enzyme Expression Coupled with the ERK, Akt and JNK Signaling Pathways. Molecules 2011, 16, 9234-9244. https://doi.org/10.3390/molecules16119234
Gu J, Sun X, Wang G, Li M, Chi M. Icariside II Enhances Nrf2 Nuclear Translocation to Upregulate Phase II Detoxifying Enzyme Expression Coupled with the ERK, Akt and JNK Signaling Pathways. Molecules. 2011; 16(11):9234-9244. https://doi.org/10.3390/molecules16119234
Chicago/Turabian StyleGu, Jing, Xuechao Sun, Guonian Wang, Mingming Li, and Meng Chi. 2011. "Icariside II Enhances Nrf2 Nuclear Translocation to Upregulate Phase II Detoxifying Enzyme Expression Coupled with the ERK, Akt and JNK Signaling Pathways" Molecules 16, no. 11: 9234-9244. https://doi.org/10.3390/molecules16119234