Differential Expression of Antimicrobial Peptides in Streptococcus pneumoniae Keratitis and STAT3-Dependent Expression of LL-37 by Streptococcus pneumoniae in Human Corneal Epithelial Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Corneal Scrapings
2.2. Identification of Bacterial Strains
2.3. Bacterial Culture
2.4. Cell Culture
2.5. RNA Isolation, Complementary DNA (cDNA) Synthesis and Quantitative PCR Analysis
2.6. Western Blot
2.7. Immunostaining
2.8. Immunohistochemistry
2.9. In Vitro Susceptibility Test of S. pneumoniae with LL-37
2.10. Statistical Analysis
3. Results
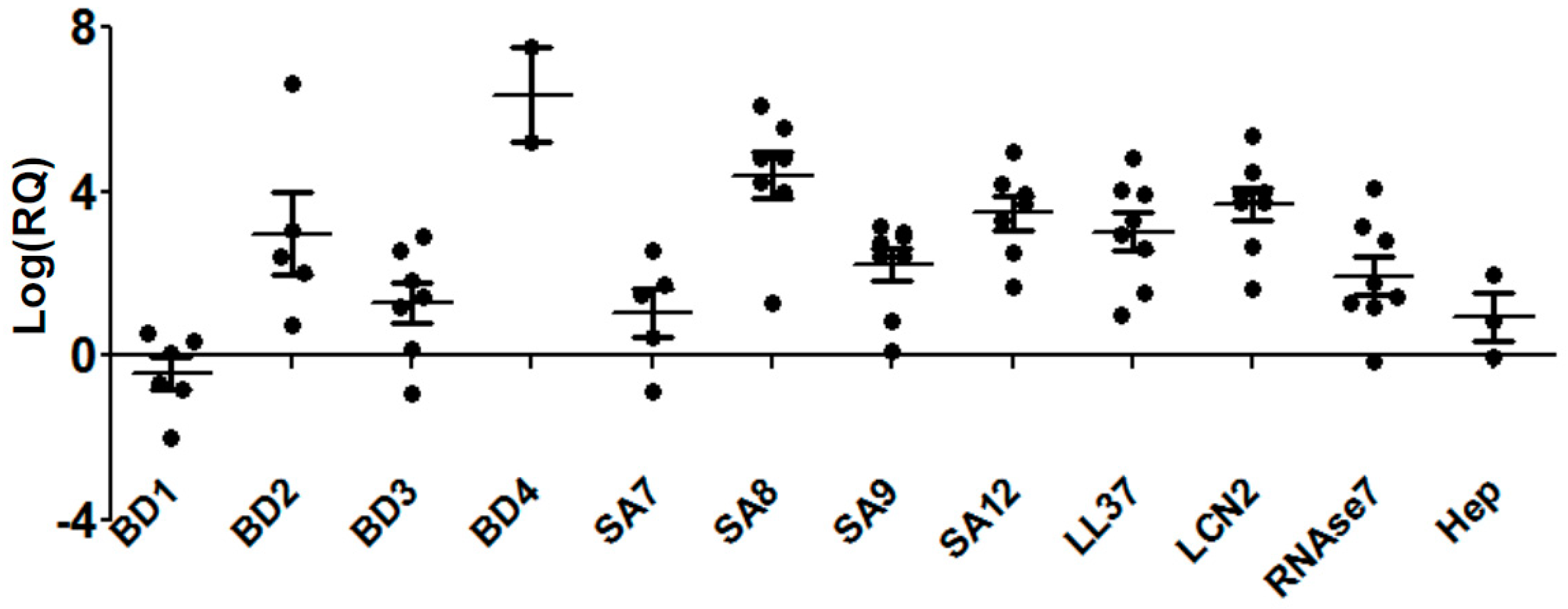
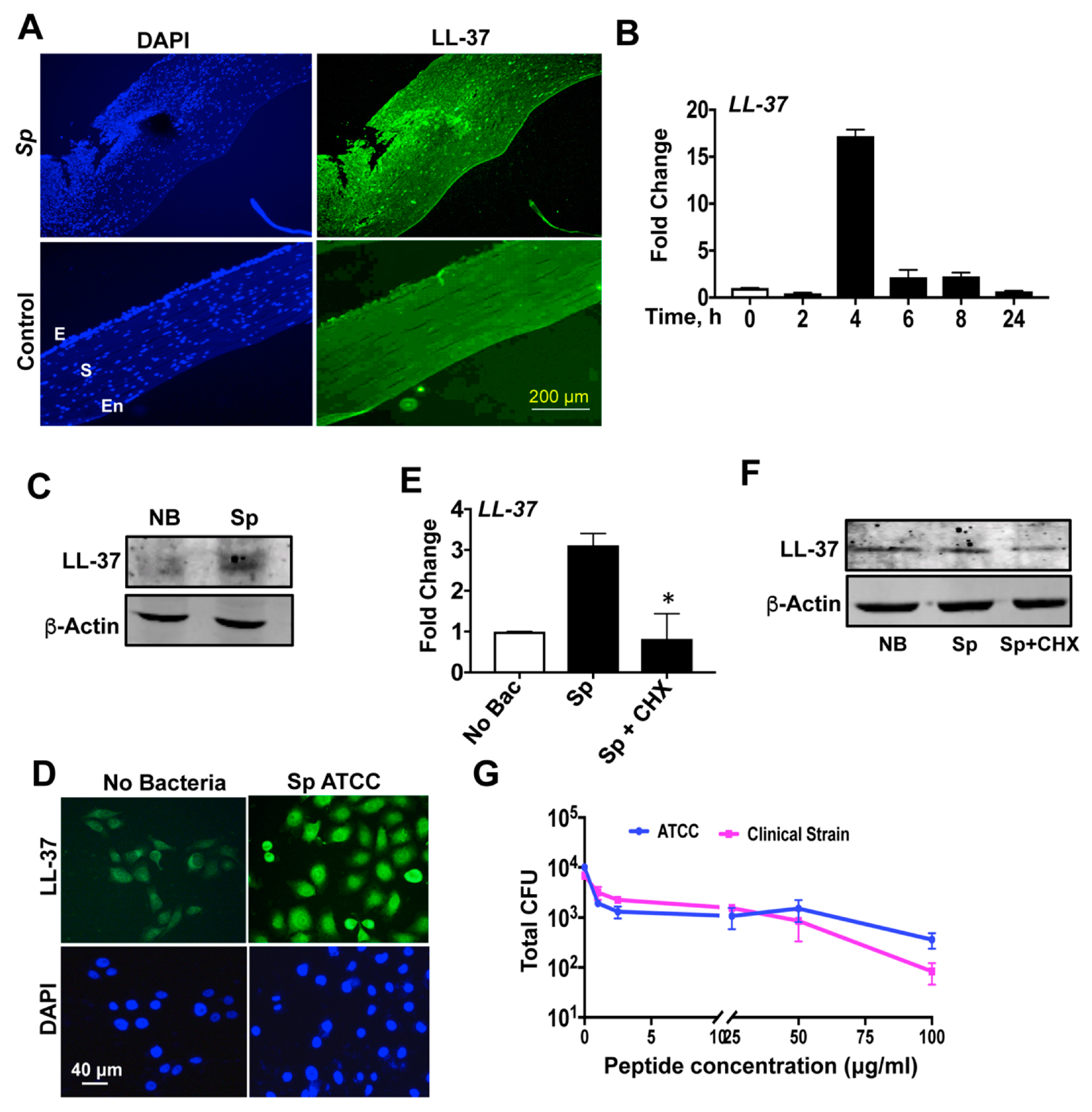
3.1. Differential Expressions of Antimicrobial Peptides during S. pneumoniae Keratitis
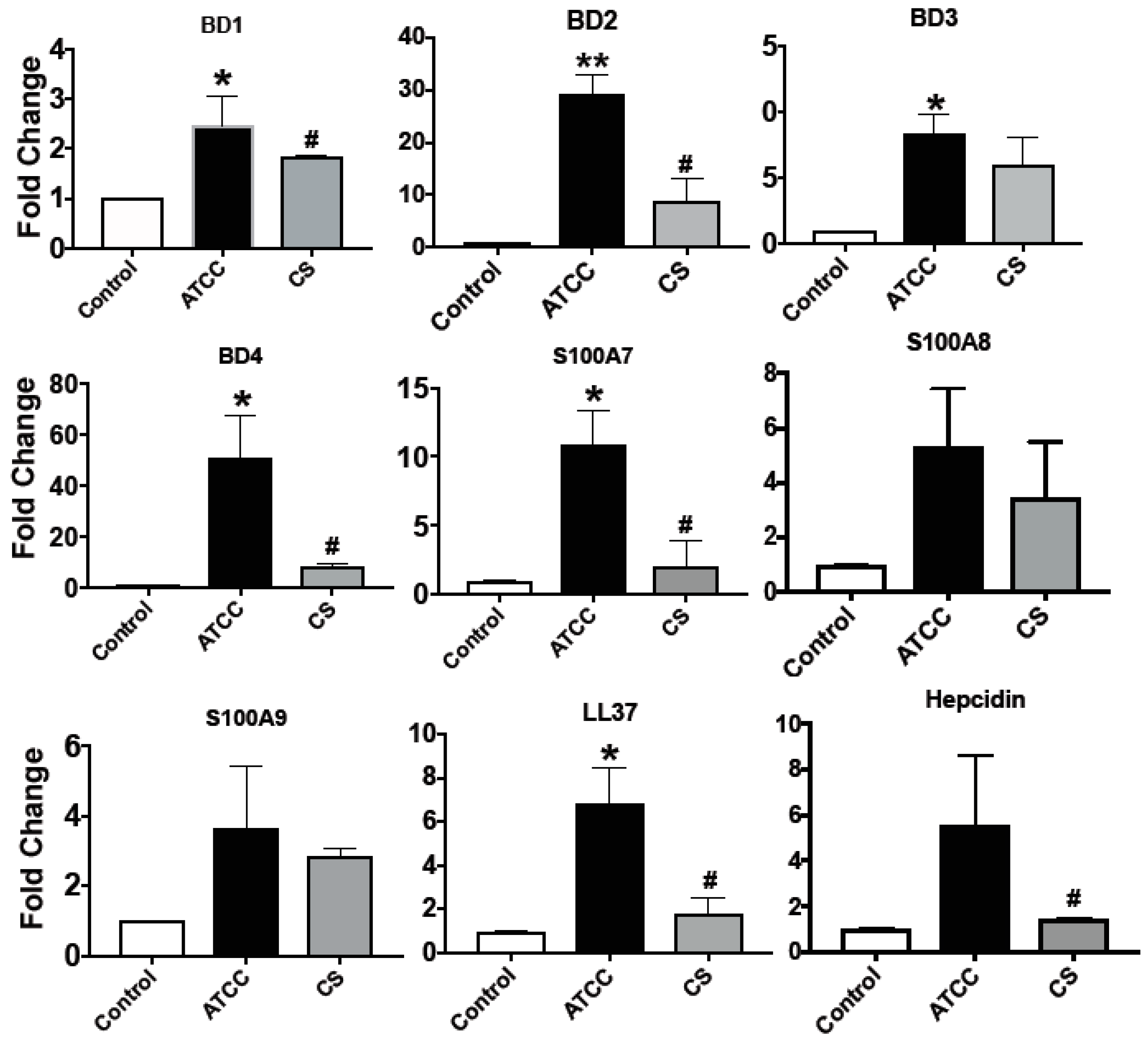
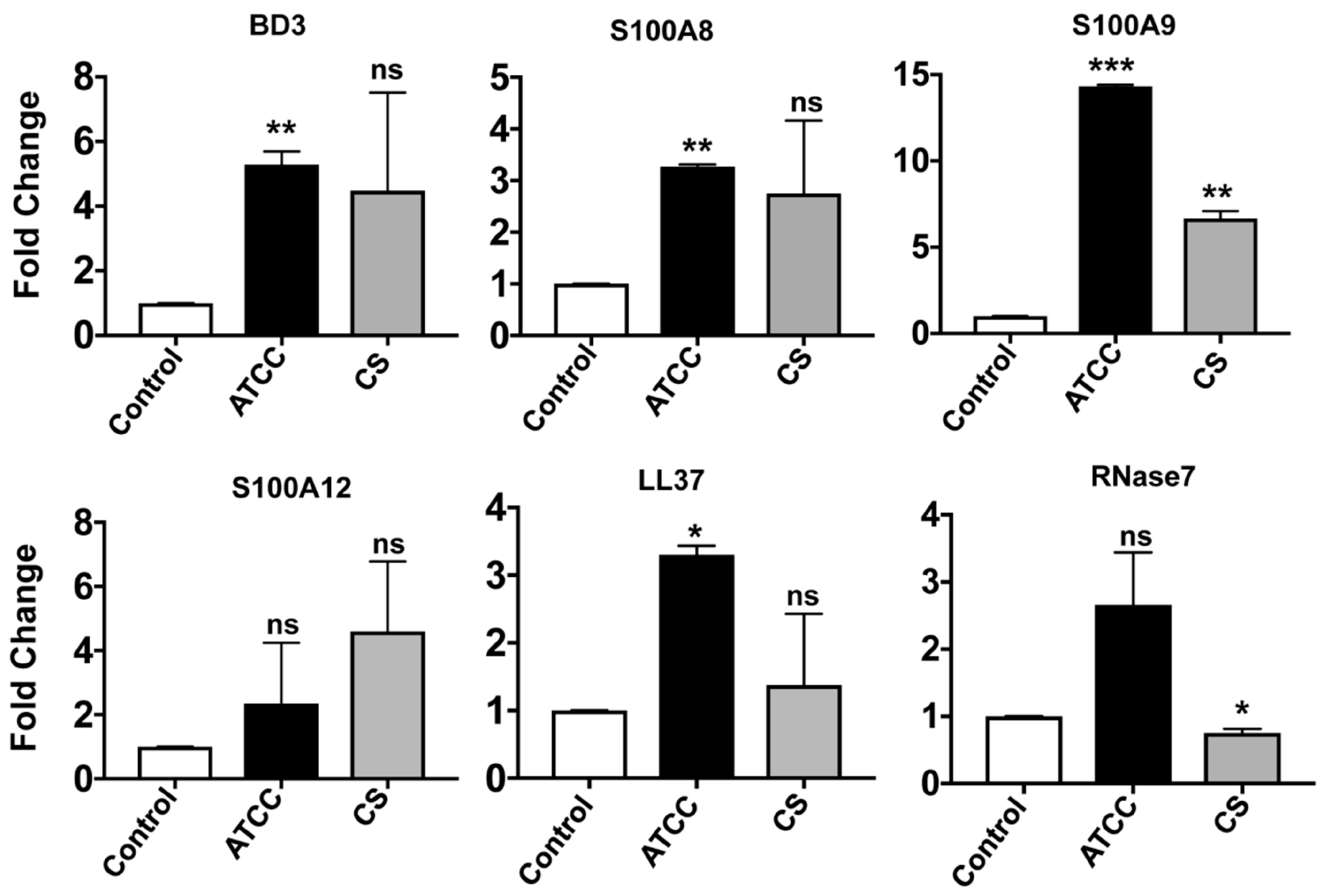
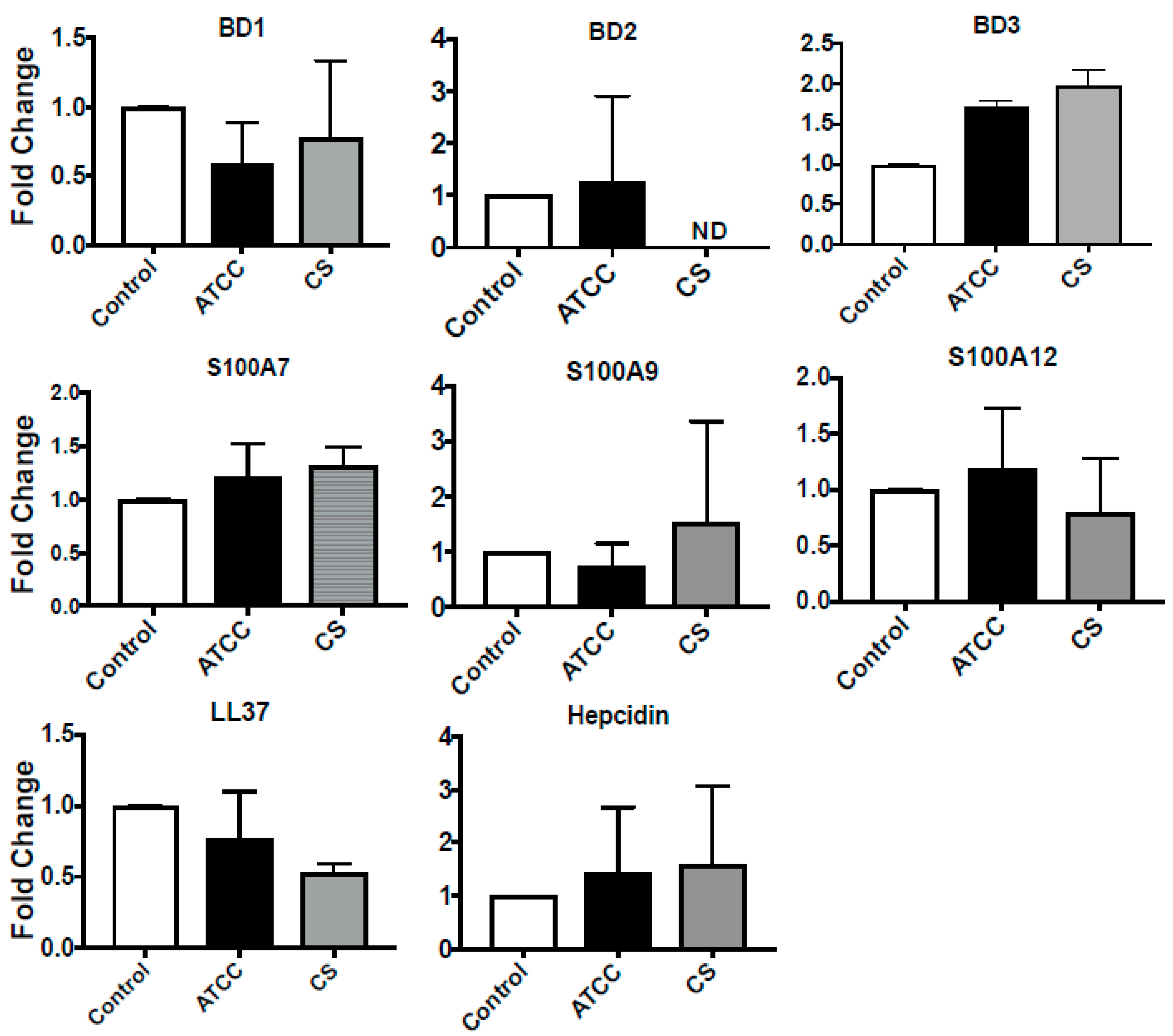
3.2. Expression of Antimicrobial Peptides in Immortalized and Primary HCECs and U937 in Response to S. pneumoniae
3.3. S. pneumoniae Induces LL-37 Expression in Human Corneal Epithelial Cells
3.4. LL-37 Exhibits Antimicrobial Activity against S. pneumoniae
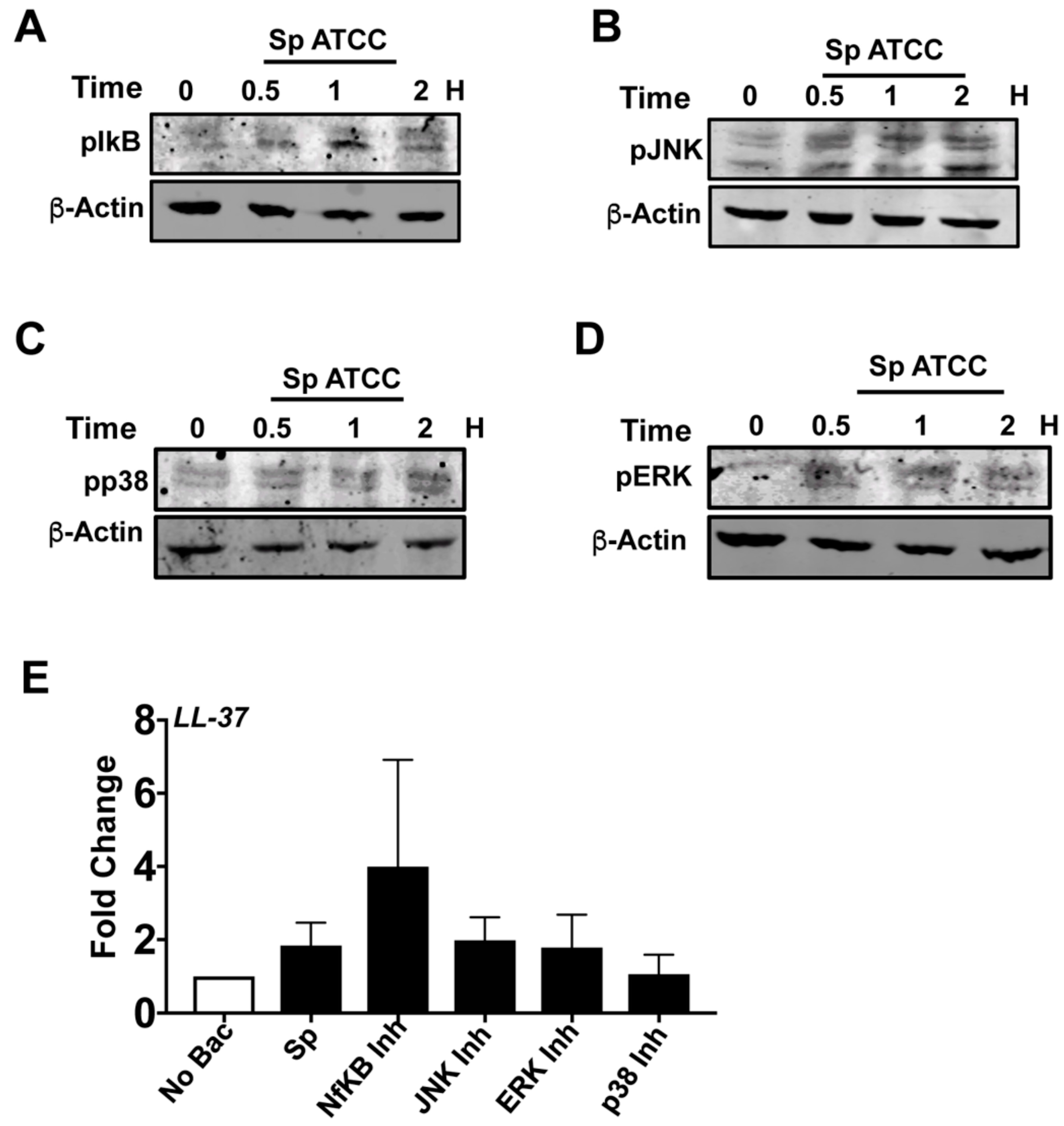
3.5. NF-κB and MAPK Pathways Are Not Responsible for LL-37 Expression by S. pneumoniaein HCECs
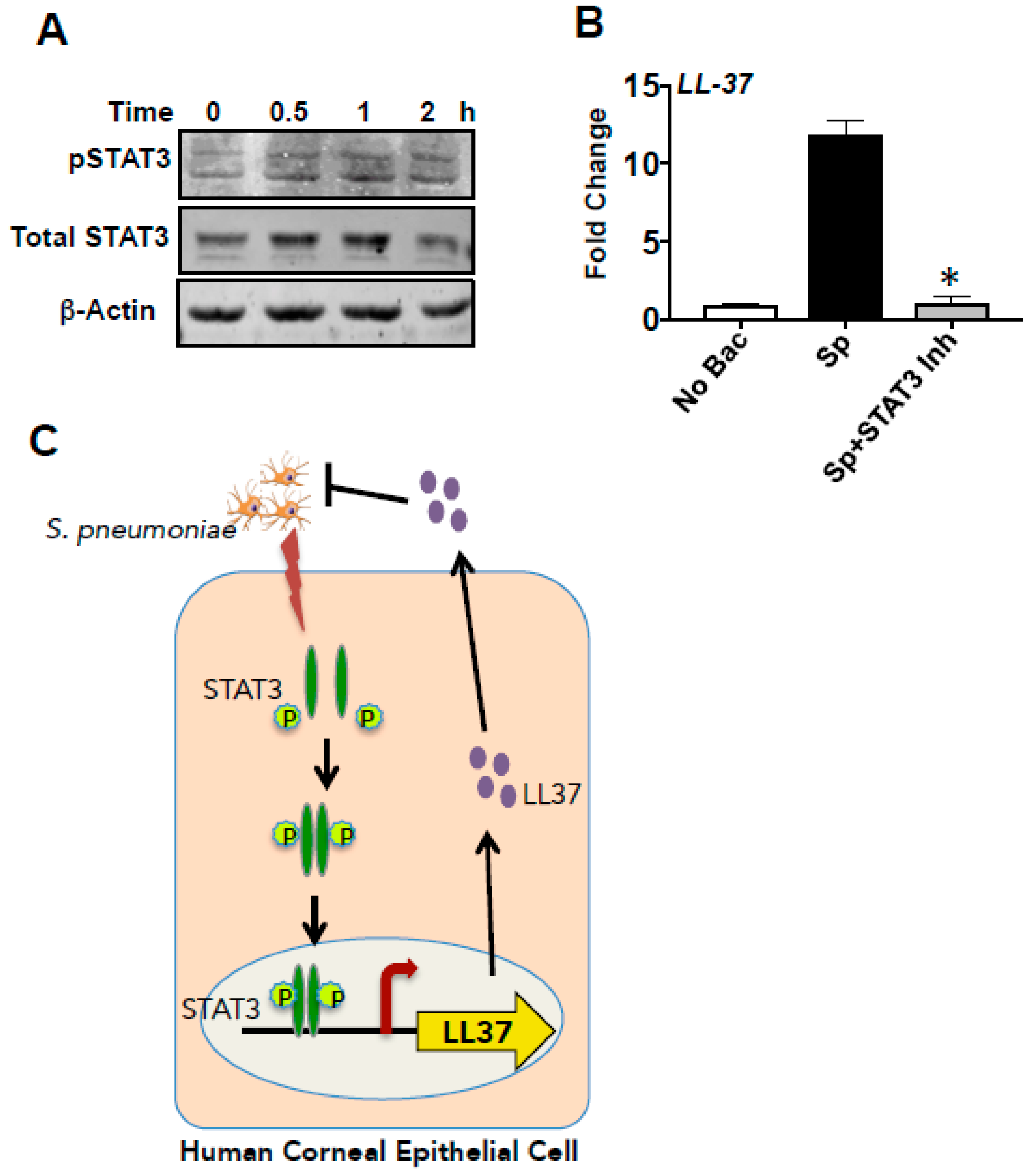
3.6. LL-37 Expression by S. pneumoniaein HCECs Is Mediated by STAT3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gupta, N.; Tandon, R.; Gupta, S.K.; Sreenivas, V.; Vashist, P. Burden of corneal blindness in India. Indian J. Community Med. 2013, 38, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.; Lalitha, P.; Srinivasan, M.; Prajna, N.V.; McLeod, S.D.; Acharya, N.R.; Lietman, T.M.; Porco, T.C. Seasonal trends of microbial keratitis in South India. Cornea 2012, 31, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Short, W.R.; Tunkel, A.R. Changing Epidemiology of Bacterial Meningitis in the United States. Curr. Infect. Dis. Rep. 2000, 2, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Tomasz, A. Antibiotic resistance in Streptococcus pneumoniae. Clin. Infect. Dis. 1997, 24 (Suppl. 1), S85–S88. [Google Scholar] [CrossRef] [PubMed]
- Heikkinen, T.; Chonmaitree, T. Importance of respiratory viruses in acute otitis media. Clin. Microbiol. Rev. 2003, 16, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Hirst, R.A.; Kadioglu, A.; O’Callaghan, C.; Andrew, P.W. The role of pneumolysin in pneumococcal pneumonia and meningitis. Clin. Exp. Immunol. 2004, 138, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Tweten, R.K. Cholesterol-dependent cytolysins, a family of versatile pore-forming toxins. Infect. Immun. 2005, 73, 6199–6209. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.M.; Lock, R.A.; Hansman, D.; Paton, J.C. Contribution of autolysin to virulence of Streptococcus pneumoniae. Infect. Immun. 1989, 57, 2324–2330. [Google Scholar] [PubMed]
- Jedrzejas, M.J. Pneumococcal virulence factors: Structure and function. Microbiol. Mol. Biol. Rev. 2001, 65, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, E.; Baba, T.; Shinohara, T.; Kitamura, H.; Sekine, A.; Komatsu, S.; Ogura, T. Ten-Year Trends and Clinical Relevance of the Antimicrobial Resistance Genotype in Respiratory Isolates of Streptococcus pneumoniae. Chemotherapy 2017, 62, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Cherazard, R.; Epstein, M.; Doan, T.L.; Salim, T.; Bharti, S.; Smith, M.A. Antimicrobial Resistant Streptococcus pneumoniae: Prevalence, Mechanisms, and Clinical Implications. Am. J. Ther. 2017, 24, e361–e369. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, M.H.; Kowalski, R.P.; Gordon, Y.J. Emerging fluoroquinolone resistance in bacterial keratitis: A 5-year review. Ophthalmology 1999, 106, 1313–1318. [Google Scholar] [CrossRef]
- Snyder, M.E.; Katz, H.R. Ciprofloxacin-resistant bacterial keratitis. Am. J. Ophthalmol. 1992, 114, 336–338. [Google Scholar] [CrossRef]
- Lalitha, P.; Manoharan, G.; Karpagam, R.; Prajna, N.V.; Srinivasan, M.; Mascarenhas, J.; Das, M.; Porco, T.C.; Lietman, T.M.; Cevallos, V.; et al. Trends in antibiotic resistance in bacterial keratitis isolates from South India. Br. J. Ophthalmol. 2017, 101, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.J.; Fleiszig, S.M. Why does the healthy cornea resist Pseudomonas aeruginosa infection? Am. J. Ophthalmol. 2013, 155, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, G.; Sun, Y.; Pearlman, E. C-Jun NH2 terminal kinase (JNK) is an essential mediator of Toll-like receptor 2-induced corneal inflammation. J. Leukoc. Biol. 2008, 83, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Karmakar, M.; Pearlman, E. CD14 mediates Toll-like receptor 4 (TLR4) endocytosis and spleen tyrosine kinase (Syk) and interferon regulatory transcription factor 3 (IRF3) activation in epithelial cells and impairs neutrophil infiltration and Pseudomonas aeruginosa killing in vivo. J. Biol. Chem. 2014, 289, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.M.; Shirtliff, M.E.; Jabra-Rizk, M.A. Antimicrobial peptides: Primeval molecules or future drugs? PLoS Pathog. 2010, 6, e1001067. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Hancock, R.E. Synergistic interactions between mammalian antimicrobial defense peptides. Antimicrob. Agents Chemother. 2001, 45, 1558–1560. [Google Scholar] [CrossRef] [PubMed]
- Yuk, J.M.; Shin, D.M.; Lee, H.M.; Yang, C.S.; Jin, H.S.; Kim, K.K.; Lee, Z.W.; Lee, S.H.; Kim, J.M.; Jo, E.K. Vitamin D3 induces autophagy in human monocytes/macrophages via cathelicidin. Cell Host Microbe 2009, 6, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Nijnik, A.; Pistolic, J.; Filewod, N.C.; Hancock, R.E. Signaling pathways mediating chemokine induction in keratinocytes by cathelicidin LL-37 and flagellin. J. Innate Immun. 2012, 4, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.H.; Ye, X.J.; Ng, T.B. Cathelicidins: Peptides with antimicrobial, immunomodulatory, anti-inflammatory, angiogenic, anticancer and procancer activities. Curr. Protein Pept. Sci. 2013, 14, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Dorschner, R.A.; Pestonjamasp, V.K.; Tamakuwala, S.; Ohtake, T.; Rudisill, J.; Nizet, V.; Agerberth, B.; Gudmundsson, G.H.; Gallo, R.L. Cutaneous injury induces the release of cathelicidin anti-microbial peptides active against group A Streptococcus. J. Investig. Dermatol. 2001, 117, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Ishida, W.; Harada, Y.; Fukuda, K.; Fukushima, A. Inhibition by the Antimicrobial Peptide LL37 of Lipopolysaccharide-Induced Innate Immune Responses in Human Corneal Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2016, 57, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.C.; Petkova, T.D.; Reins, R.Y.; Proske, R.J.; McDermott, A.M. Multifunctional roles of human cathelicidin (LL-37) at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2369–2380. [Google Scholar] [CrossRef] [PubMed]
- Strandberg, K.L.; Richards, S.M.; Gunn, J.S. Cathelicidin antimicrobial peptide expression is not induced or required for bacterial clearance during salmonella enterica infection of human monocyte-derived macrophages. Infect. Immun. 2012, 80, 3930–3938. [Google Scholar] [CrossRef] [PubMed]
- LaRock, C.N.; Dohrmann, S.; Todd, J.; Corriden, R.; Olson, J.; Johannssen, T.; Lepenies, B.; Gallo, R.L.; Ghosh, P.; Nizet, V. Group A Streptococcal M1 Protein Sequesters Cathelicidin to Evade Innate Immune Killing. Cell Host Microbe 2015, 18, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal epithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Sharma, P.; Guha, S.; Garg, P.; Roy, S. Differential expression of antimicrobial peptides in corneal infection and regulation of antimicrobial peptides and reactive oxygen species by type III secretion system of Pseudomonas aeruginosa. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Sun, Y.; Pearlman, E. Interferon-gamma-induced MD-2 protein expression and lipopolysaccharide (LPS) responsiveness in corneal epithelial cells is mediated by Janus tyrosine kinase-2 activation and direct binding of STAT1 protein to the MD-2 promoter. J. Biol. Chem. 2011, 286, 23753–23762. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Bonfield, T.; Tartakoff, A.M. Non-apoptotic toxicity of Pseudomonas aeruginosa toward murine cells. PLoS ONE 2013, 8, e54245. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Chaurasia, S.; Ramachandran, C.; Roy, S. SLC4A11 depletion impairs NRF2 mediated antioxidant signaling and increases reactive oxygen species in human corneal endothelial cells during oxidative stress. Sci. Rep. 2017, 7, 4074. [Google Scholar] [CrossRef] [PubMed]
- Mustapha, P.; Paris, I.; Garcia, M.; Tran, C.T.; Cremniter, J.; Garnier, M.; Faure, J.P.; Barthes, T.; Boneca, I.G.; Morel, F.; et al. Chemokines and antimicrobial peptides have a cag-dependent early response to Helicobacter pylori infection in primary human gastric epithelial cells. Infect. Immun. 2014, 82, 2881–2889. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.A.; McClellan, S.A.; Vistisen, K.S.; Hazlett, L.D. HIF-1alpha is essential for effective PMN bacterial killing, antimicrobial peptide production and apoptosis in Pseudomonas aeruginosa keratitis. PLoS Pathog. 2013, 9, e1003457. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Santiago, B.; Hernandez-Pando, R.; Carranza, C.; Juarez, E.; Contreras, J.L.; Aguilar-Leon, D.; Torres, M.; Sada, E. Expression of cathelicidin LL-37 during Mycobacterium tuberculosis infection in human alveolar macrophages, monocytes, neutrophils, and epithelial cells. Infect. Immun. 2008, 76, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Johansson, L.; Thulin, P.; Sendi, P.; Hertzen, E.; Linder, A.; Akesson, P.; Low, D.E.; Agerberth, B.; Norrby-Teglund, A. Cathelicidin LL-37 in severe Streptococcus pyogenes soft tissue infections in humans. Infect. Immun. 2008, 76, 3399–3404. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Marla, S.; Praneetha, D.C. Recognition of Corynebacterium pseudodiphtheriticum by Toll-like receptors and up-regulation of antimicrobial peptides in human corneal epithelial cells. Virulence 2015, 6, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.S.; Priya, J.L.; Leal, S.M., Jr.; Toska, J.; Rietsch, A.; Prajna, V.; Pearlman, E.; Lalitha, P. Host response and bacterial virulence factor expression in Pseudomonas aeruginosa and Streptococcus pneumoniae corneal ulcers. PLoS ONE 2013, 8, e64867. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, M.; Katsnelson, M.; Malak, H.A.; Greene, N.G.; Howell, S.J.; Hise, A.G.; Camilli, A.; Kadioglu, A.; Dubyak, G.R.; Pearlman, E. Neutrophil IL-1beta processing induced by pneumolysin is mediated by the NLRP3/ASC inflammasome and caspase-1 activation and is dependent on K+ efflux. J. Immunol. 2015, 194, 1763–1775. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Karmakar, M.; Roy, S.; Ramadan, R.T.; Williams, S.R.; Howell, S.; Shive, C.L.; Han, Y.; Stopford, C.M.; Rietsch, A.; et al. TLR4 and TLR5 on corneal macrophages regulate Pseudomonas aeruginosa keratitis by signaling through MyD88-dependent and -independent pathways. J. Immunol. 2010, 185, 4272–4283. [Google Scholar] [CrossRef] [PubMed]
- Mendez-Samperio, P.; Miranda, E.; Trejo, A. Expression and secretion of cathelicidin LL-37 in human epithelial cells after infection by Mycobacterium bovis Bacillus Calmette-Guerin. Clin. Vac. Immunol. 2008, 15, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Domenico, J.; Jia, Y.; Lucas, J.J.; Gelfand, E.W. NF-kappaB-dependent induction of cathelicidin-related antimicrobial peptide in murine mast cells by lipopolysaccharide. Int. Arch. Allergy Immunol. 2009, 150, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, E.; Nylen, F.; Johansson, K.; Arner, E.; Cebula, M.; Farmand, S.; Ottosson, H.; Stromberg, R.; Gudmundsson, G.H.; Agerberth, B.; et al. Entinostat up-regulates the CAMP gene encoding LL-37 via activation of STAT3 and HIF-1alpha transcription factors. Sci. Rep. 2016, 6, 33274. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Borregaard, N.; Koeffler, H.P. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D3. FASEB J. 2005, 19, 1067–1077. [Google Scholar] [CrossRef] [PubMed]
- Schust, J.; Sperl, B.; Hollis, A.; Mayer, T.U.; Berg, T. Stattic: A small-molecule inhibitor of STAT3 activation and dimerization. Chem. Biol. 2006, 13, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Ramakrishnan, R.; Bharathi, M.J.; Amuthan, M.; Viswanathan, S. Prevalence of bacterial pathogens causing ocular infections in South India. Indian J. Pathol. Microbiol. 2010, 53, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Chawla, K.; Gurung, B.; Mukhopadhyay, C.; Bairy, I. Reporting Emerging Resistance of Streptococcus pneumoniae from India. J. Glob. Infect. Dis. 2010, 2, 10–14. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.S.; Cade, J.E.; Al-Abed, M.; Shanmuganathan, V.; Gupta, R.; Bhan, A.; Tighe, P.J.; Dua, H.S. The spectrum of antimicrobial peptide expression at the ocular surface. Investig. Ophthalmol. Vis. Sci. 2005, 46, 1379–1385. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Zhang, J.; Yu, F.S. Toll-like receptor 2-mediated expression of beta-defensin-2 in human corneal epithelial cells. Microbes Infect. 2006, 8, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Otri, A.M.; Mohammed, I.; Abedin, A.; Cao, Z.; Hopkinson, A.; Panjwani, N.; Dua, H.S. Antimicrobial peptides expression by ocular surface cells in response to Acanthamoeba castellanii: An in vitro study. Br. J. Ophthalmol. 2010, 94, 1523–1527. [Google Scholar] [CrossRef] [PubMed]
- Bergman, P.; Johansson, L.; Asp, V.; Plant, L.; Gudmundsson, G.H.; Jonsson, A.B.; Agerberth, B. Neisseria gonorrhoeae downregulates expression of the human antimicrobial peptide LL-37. Cell. Microbiol. 2005, 7, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, B.; Regnault, B.; Guo, J.; Zhang, Z.; Stanley, S.L., Jr.; Sansonetti, P.J.; Pedron, T. Virulent Shigella flexneri subverts the host innate immune response through manipulation of antimicrobial peptide gene expression. J. Exp. Med. 2008, 205, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Hase, K.; Murakami, M.; Iimura, M.; Cole, S.P.; Horibe, Y.; Ohtake, T.; Obonyo, M.; Gallo, R.L.; Eckmann, L.; Kagnoff, M.F. Expression of LL-37 by human gastric epithelial cells as a potential host defense mechanism against Helicobacter pylori. Gastroenterology 2003, 125, 1613–1625. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhu, M.; Yang, Z.; Pan, X.; Zhang, Y.; Wang, Q.; Xiao, W. LL-37 secreted by epithelium promotes fibroblast collagen production: A potential mechanism of small airway remodeling in chronic obstructive pulmonary disease. Lab. Investig. 2014, 94, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Cruse, G.; Fernandes, V.E.; de Salort, J.; Pankhania, D.; Marinas, M.S.; Brewin, H.; Andrew, P.W.; Bradding, P.; Kadioglu, A. Human lung mast cells mediate pneumococcal cell death in response to activation by pneumolysin. J. Immunol. 2010, 184, 7108–7115. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gao, N.; Standiford, T.J.; Gallo, R.L.; Yu, F.S. Topical flagellin protects the injured corneas from Pseudomonas aeruginosa infection. Microbes Infect. 2010, 12, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Gordon, Y.J.; Huang, L.C.; Romanowski, E.G.; Yates, K.A.; Proske, R.J.; McDermott, A.M. Human cathelicidin (LL-37), a multifunctional peptide, is expressed by ocular surface epithelia and has potent antibacterial and antiviral activity. Curr. Eye Res. 2005, 30, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Sakoulas, G.; Rose, W.; Nonejuie, P.; Olson, J.; Pogliano, J.; Humphries, R.; Nizet, V. Ceftaroline restores daptomycin activity against daptomycin-nonsusceptible vancomycin-resistant Enterococcus faecium. Antimicrob. Agents Chemother. 2014, 58, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Zahner, D.; Zhou, X.; Chancey, S.T.; Pohl, J.; Shafer, W.M.; Stephens, D.S. Human antimicrobial peptide LL-37 induces MefE/Mel-mediated macrolide resistance in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2010, 54, 3516–3519. [Google Scholar] [CrossRef] [PubMed]
- Peric, M.; Koglin, S.; Kim, S.M.; Morizane, S.; Besch, R.; Prinz, J.C.; Ruzicka, T.; Gallo, R.L.; Schauber, J. IL-17A enhances vitamin D3-induced expression of cathelicidin antimicrobial peptide in human keratinocytes. J. Immunol. 2008, 181, 8504–8512. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; McAleer, J.P.; Zheng, M.; Pociask, D.A.; Kaplan, M.H.; Qin, S.; Reinhart, T.A.; Kolls, J.K. Innate Stat3-mediated induction of the antimicrobial protein Reg3gamma is required for host defense against MRSA pneumonia. J. Exp. Med. 2013, 210, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Wrighting, D.M.; Andrews, N.C. Interleukin-6 induces hepcidin expression through STAT3. Blood 2006, 108, 3204–3209. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Cui, X.; Ma, T.; Liu, Y.; Li, A.; Huang, M. Paeoniflorin upregulates beta-defensin-2 expression in human bronchial epithelial cell through the p38 MAPK, ERK, and NF-kappaB signaling pathways. Inflammation 2014, 37, 1468–1475. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Streptococcus Pneumoniae |
|---|---|
| Age | 25 to 90 |
| Mean (SD) | 52.25 (20.176) |
| Sex | |
| Male (%) | 75 |
| Female (%) | 25 |
| Hypopyon | |
| Yes (%) | 25 |
| No (%) | 75 |
| Occupation | |
| Agriculture/Manual Labor (%) Desk Jobs (%) | 75 25 |
| Size of Ulcer | |
| <5 mm (%) 5–15 mm (%) >15 mm (%) | 37.5 25 37.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, P.; Sharma, N.; Mishra, P.; Joseph, J.; Mishra, D.K.; Garg, P.; Roy, S. Differential Expression of Antimicrobial Peptides in Streptococcus pneumoniae Keratitis and STAT3-Dependent Expression of LL-37 by Streptococcus pneumoniae in Human Corneal Epithelial Cells. Pathogens 2019, 8, 31. https://doi.org/10.3390/pathogens8010031
Sharma P, Sharma N, Mishra P, Joseph J, Mishra DK, Garg P, Roy S. Differential Expression of Antimicrobial Peptides in Streptococcus pneumoniae Keratitis and STAT3-Dependent Expression of LL-37 by Streptococcus pneumoniae in Human Corneal Epithelial Cells. Pathogens. 2019; 8(1):31. https://doi.org/10.3390/pathogens8010031
Chicago/Turabian StyleSharma, Prerana, Natalia Sharma, Priyasha Mishra, Joveeta Joseph, Dilip K. Mishra, Prashant Garg, and Sanhita Roy. 2019. "Differential Expression of Antimicrobial Peptides in Streptococcus pneumoniae Keratitis and STAT3-Dependent Expression of LL-37 by Streptococcus pneumoniae in Human Corneal Epithelial Cells" Pathogens 8, no. 1: 31. https://doi.org/10.3390/pathogens8010031