Water-Soluble Extract from Actinidia arguta (Siebold & Zucc.) Planch. ex Miq. and Perilla frutescens (L.) Britton, ACTPER, Ameliorates a Dry Skin-Induced Itch in a Mice Model and Promotes Filaggrin Expression by Activating the AhR Signaling in HaCaT Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Water-Soluble Extracts from Plants
2.2. High-Performance Liquid Chromatography (HPLC) Analysis
2.3. Experimental Animals
2.4. Acetone-Induced Dry Skin Mice Model
2.5. Measurement of Skin Properties
2.6. Immunofluorescence Assay
2.7. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR) Analysis
2.8. Cell Culture and Reagents
2.9. Western Blot Analysis
2.10. Luciferase Reporter Plasmid Assay
2.11. Molecular Docking Simulation
2.12. Calculation of Binding Energy
2.13. Statistical Analysis
3. Results
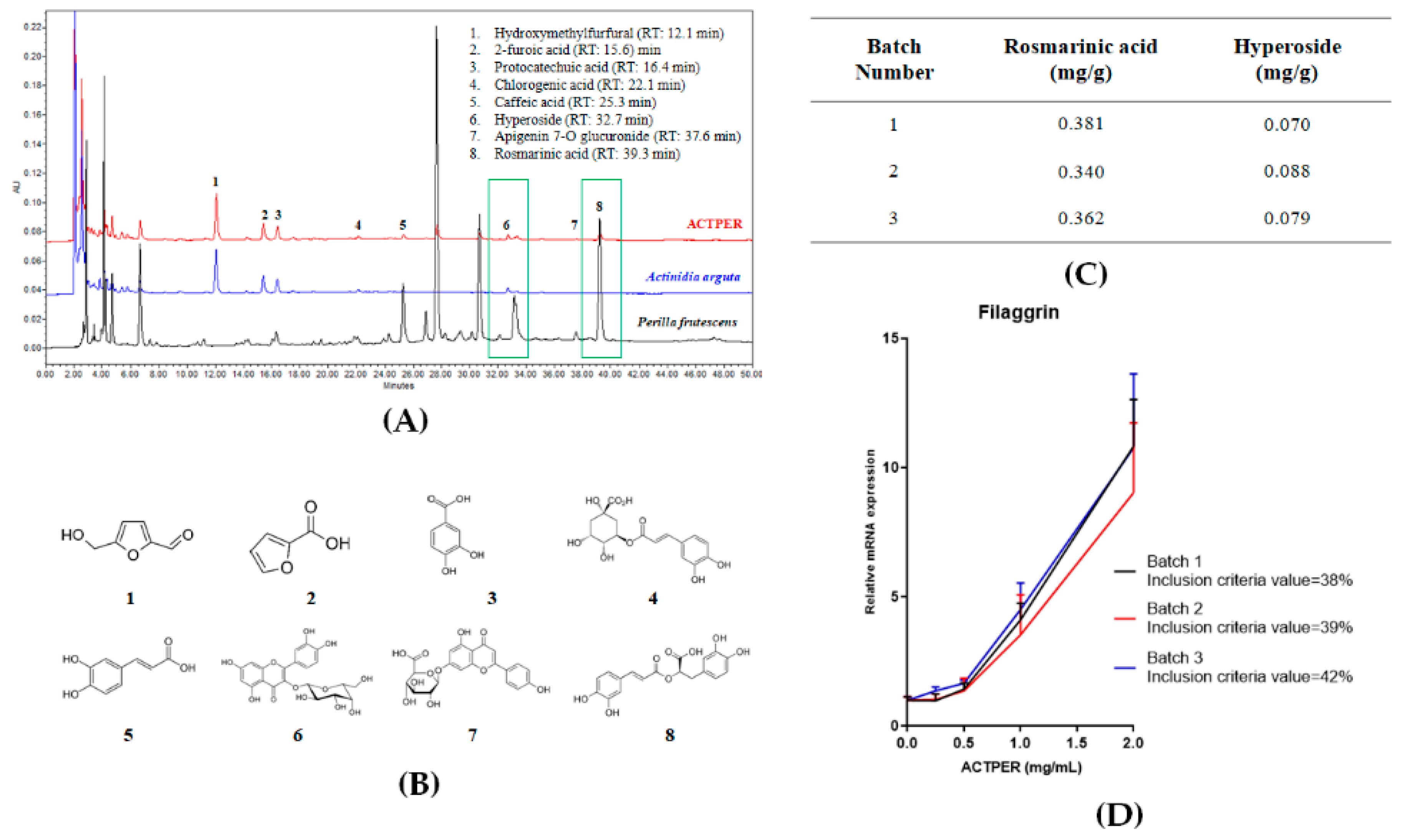
3.1. Quality of ACTPER is Measured by HPLC and Cell-Based Bioassay
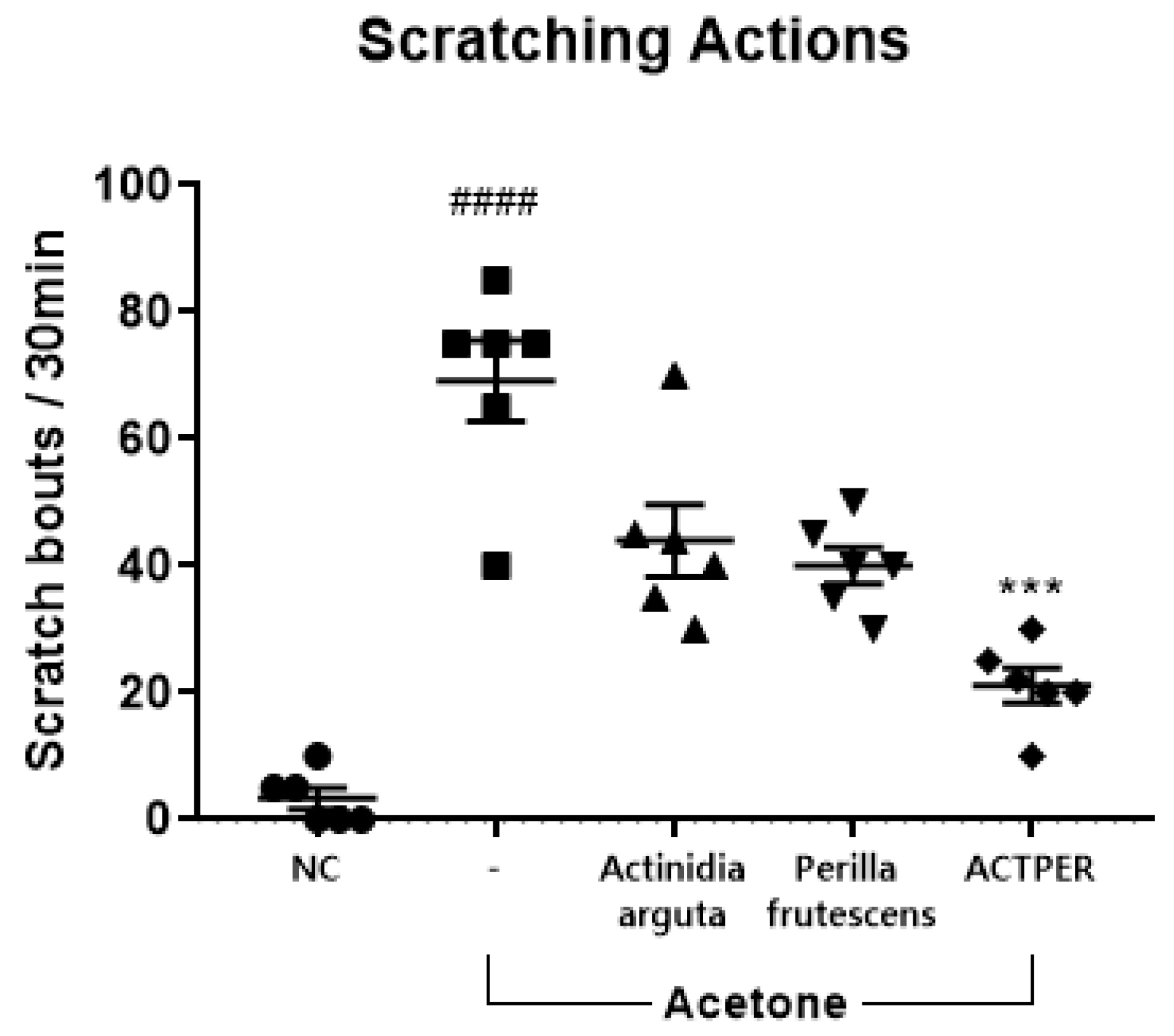
3.2. The Combined Use of Actinidia arguta and Perilla frutescens Synergistically Improves Acetone-Induced Itch in Mice
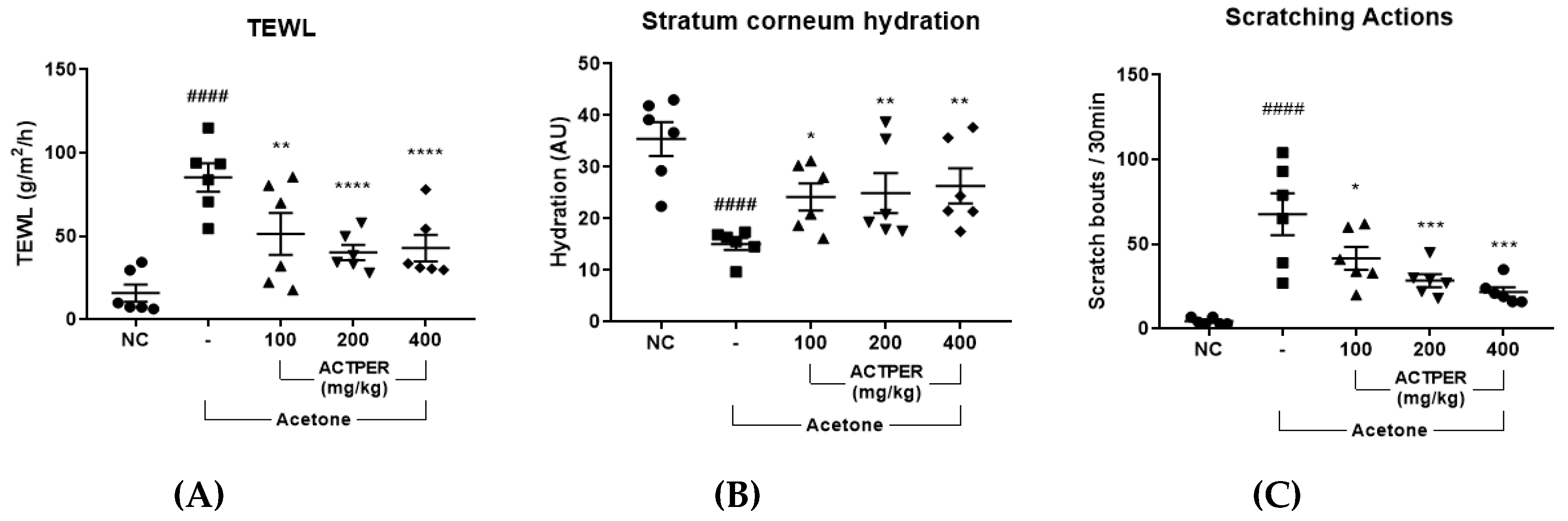
3.3. ACTPER Ameliorates Dry Skin-Induced Itch in Acetone-Treated Mice
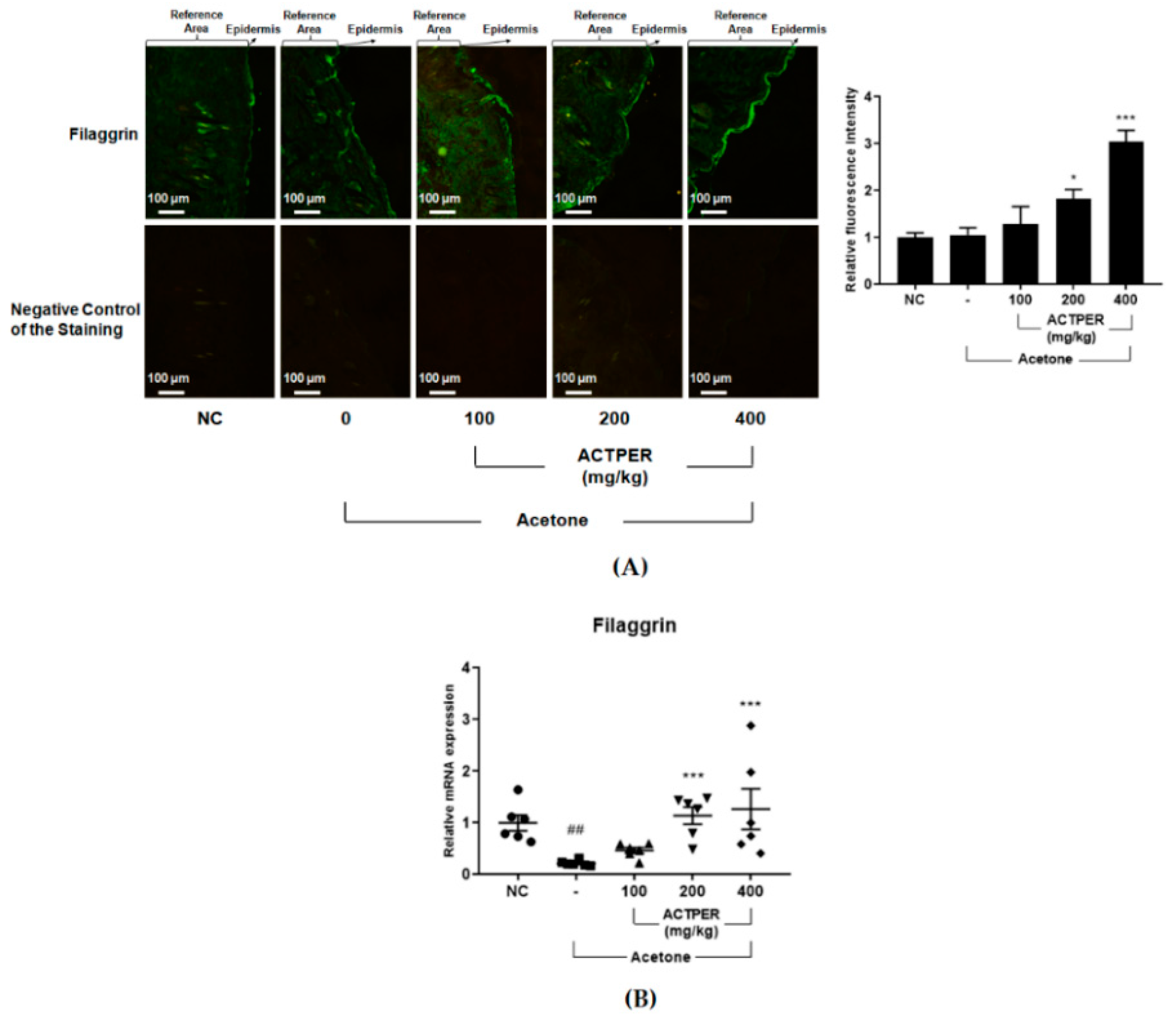
3.4. ACTPER Promotes the Expression Level of Filaggrin in Acetone-Treated Mice Back Skin
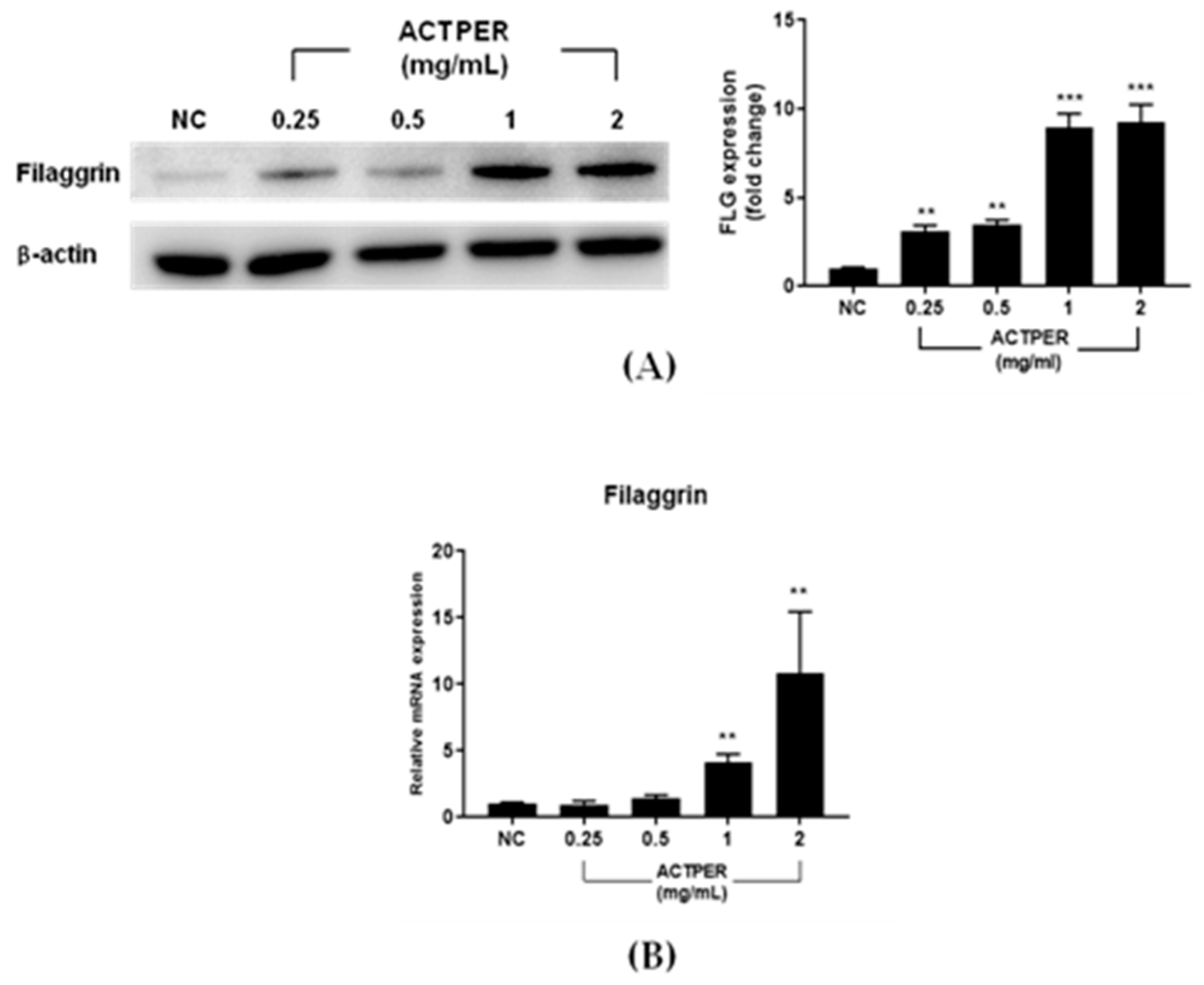
3.5. ACTPER Promotes the Expression of Filaggrin in HaCaT cells
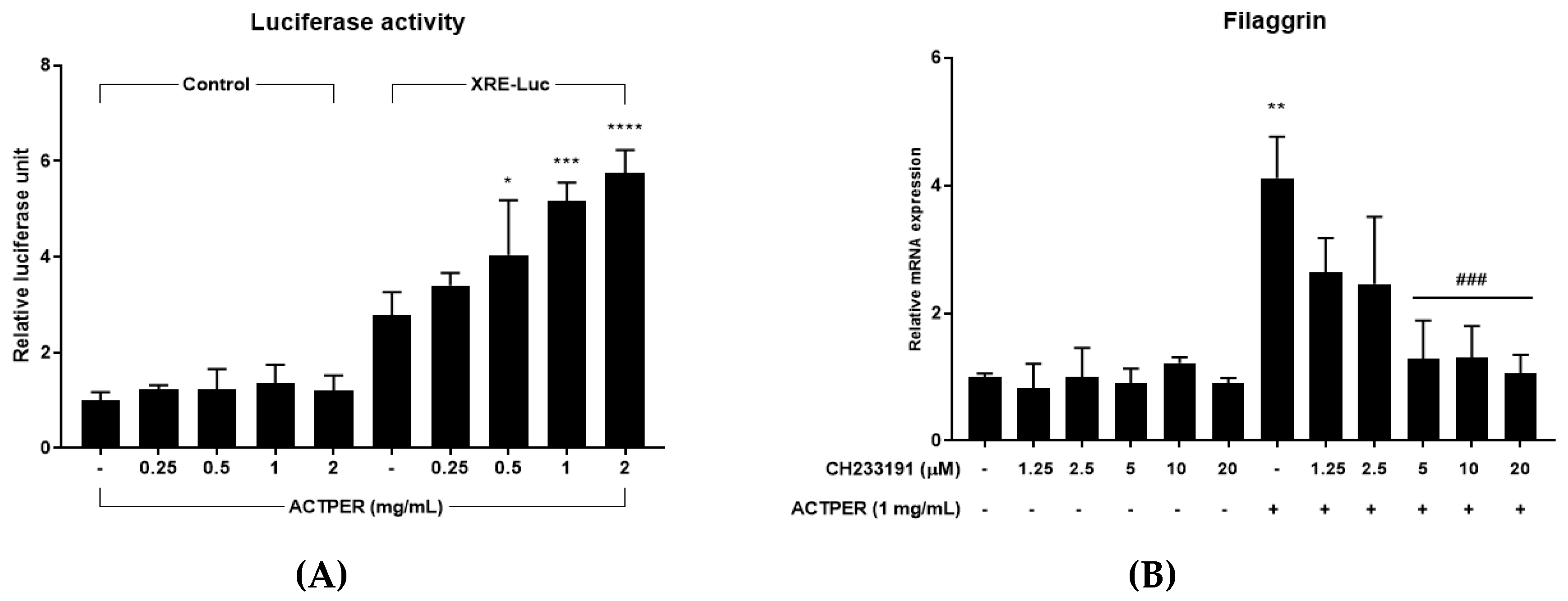
3.6. Effect of ACTPER on Filaggrin Expression was Mediated by the AhR Signaling Pathway
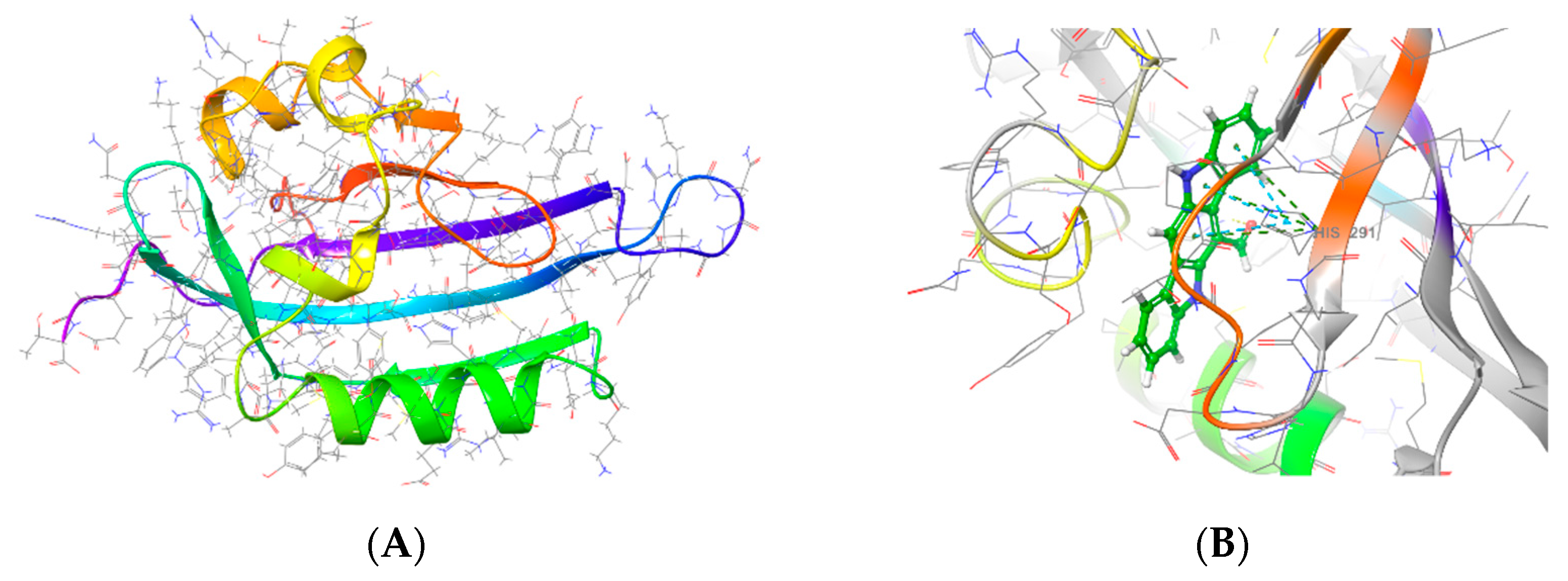
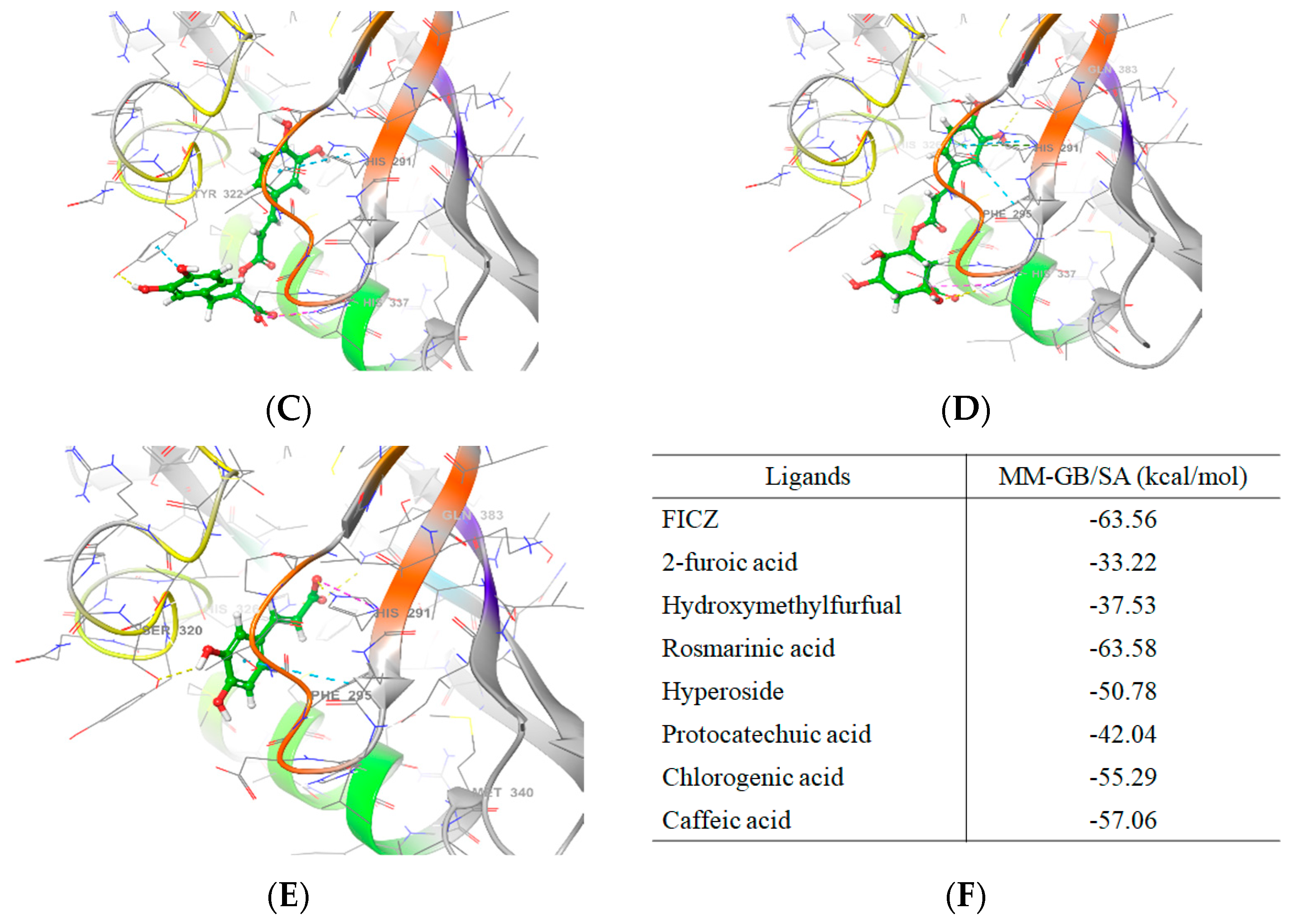
3.7. ACTPER was Predicted to Contain Chemical Compounds Expected to Bind to AhR by Molecular Docking Simulation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Han, L.; Dong, X. Itch mechanisms and circuits. Annu. Rev. Biophys. 2014, 43, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Garibyan, L.; Rheingold, C.G.; Lerner, E.A. Understanding the pathophysiology of itch. Dermatol. Ther. 2013, 26, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Caccavale, S.; Bove, D.; Bove, R.M.; LA Montagna, M. Skin and brain: Itch and psychiatric disorders. G. Ital. Dermatol. Venereol. 2016, 151, 525–529. [Google Scholar]
- Potenzieri, C.; Undem, B.J. Basic mechanisms of itch. Clin. Exp. Allergy 2012, 42, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Hu, H. TRP Channels as Drug Targets to Relieve Itch. Pharmaceuticals 2018, 11, 100. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Gupta, R.; Jordt, S.E.; Chen, Y.; Liedtke, W.B. Regulation of Pain and Itch by TRP Channels. Neurosci. Bull. 2018, 34, 120–142. [Google Scholar] [CrossRef] [PubMed]
- Kittaka, H.; Tominaga, M. The molecular and cellular mechanisms of itch and the involvement of TRP channels in the peripheral sensory nervous system and skin. Allergol. Int. 2017, 66, 22–30. [Google Scholar] [CrossRef]
- Rossbach, K.; Nassenstein, C.; Gschwandtner, M.; Schnell, D.; Sander, K.; Seifert, R.; Stark, H.; Kietzmann, M.; Baumer, W. Histamine H1, H3 and H4 receptors are involved in pruritus. Neuroscience 2011, 190, 89–102. [Google Scholar] [CrossRef]
- Manenti, L.; Tansinda, P.; Vaglio, A. Uraemic pruritus: Clinical characteristics, pathophysiology and treatment. Drugs 2009, 69, 251–263. [Google Scholar] [CrossRef]
- Yosipovitch, G. Dry skin and impairment of barrier function associated with itch—New insights. Int. J. Cosmet. Sci. 2004, 26, 1–7. [Google Scholar] [CrossRef]
- Rawlings, A.V.; Harding, C.R. Moisturization and skin barrier function. Dermatol. Ther. 2004, 17 (Suppl. 1), 43–48. [Google Scholar] [CrossRef] [PubMed]
- Proksch, E.; Folster-Holst, R.; Jensen, J.M. Skin barrier function, epidermal proliferation and differentiation in eczema. J. Dermatol. Sci. 2006, 43, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Moniaga, C.S.; Kabashima, K. Filaggrin in atopic dermatitis: Flaky tail mice as a novel model for developing drug targets in atopic dermatitis. Inflamm. Allergy Drug Targets 2011, 10, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Floudas, A.; Saunders, S.P.; Moran, T.; Schwartz, C.; Hams, E.; Fitzgerald, D.C.; Johnston, J.A.; Ogg, G.S.; McKenzie, A.N.; Walsh, P.T.; et al. IL-17 Receptor A Maintains and Protects the Skin Barrier to Prevent Allergic Skin Inflammation. J. Immunol. 2017, 199, 707–717. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Q.; Liu, H.; Lu, C.; Liang, C.L.; Qiu, F.; Han, L.; Dai, Z. Esculetin Ameliorates Psoriasis-Like Skin Disease in Mice by Inducing CD4(+)Foxp3(+) Regulatory T Cells. Front. Immunol. 2018, 9, 2092. [Google Scholar] [CrossRef]
- Oyoshi, M.K.; Murphy, G.F.; Geha, R.S. Filaggrin-deficient mice exhibit TH17-dominated skin inflammation and permissiveness to epicutaneous sensitization with protein antigen. J. Allergy Clin. Immunol. 2009, 124, 485–493.e1. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C. Antioxidant Functions of the Aryl Hydrocarbon Receptor. Stem. Cells Int. 2016, 2016, 7943495. [Google Scholar] [CrossRef]
- de Araujo, E.F.; Feriotti, C.; Galdino, N.A.L.; Preite, N.W.; Calich, V.L.G.; Loures, F.V. The IDO-AhR Axis Controls Th17/Treg Immunity in a Pulmonary Model of Fungal Infection. Front. Immunol. 2017, 8, 880. [Google Scholar] [CrossRef]
- Napolitano, M.; Patruno, C. Aryl hydrocarbon receptor (AhR) a possible target for the treatment of skin disease. Med. Hypotheses 2018, 116, 96–100. [Google Scholar] [CrossRef]
- Haas, K.; Weighardt, H.; Deenen, R.; Kohrer, K.; Clausen, B.; Zahner, S.; Boukamp, P.; Bloch, W.; Krutmann, J.; Esser, C. Aryl Hydrocarbon Receptor in Keratinocytes Is Essential for Murine Skin Barrier Integrity. J. Investig. Dermatol. 2016, 136, 2260–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, G.; Hashimoto-Hachiya, A.; Kiyomatsu-Oda, M.; Takemura, M.; Ohno, F.; Ito, T.; Morino-Koga, S.; Mitoma, C.; Nakahara, T.; Uchi, H.; et al. Aryl hydrocarbon receptor activation restores filaggrin expression via OVOL1 in atopic dermatitis. Cell Death Dis. 2017, 8, e2931. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Z.; Zhong, W.L.; Hu, H.; Chen, X.F.; Zhang, W.; Huang, H.Y.; Yu, B.; Dou, X. Aryl hydrocarbon receptor polymorphisms are associated with dry skin phenotypes in Chinese patients with atopic dermatitis. Clin. Exp. Dermatol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hirano, A.; Goto, M.; Mitsui, T.; Hashimoto-Hachiya, A.; Tsuji, G.; Furue, M. Antioxidant Artemisia princeps Extract Enhances the Expression of Filaggrin and Loricrin via the AHR/OVOL1 Pathway. Int. J. Mol. Sci. 2017, 18, 1948. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto-Hachiya, A.; Tsuji, G.; Murai, M.; Yan, X.; Furue, M. Upregulation of FLG, LOR, and IVL Expression by Rhodiola crenulata Root Extract via Aryl Hydrocarbon Receptor: Differential Involvement of OVOL1. Int. J. Mol. Sci. 2018, 19, 1654. [Google Scholar] [CrossRef] [PubMed]
- Bae, M.J.; Lim, S.; Lee, D.S.; Ko, K.R.; Lee, W.; Kim, S. Water soluble extracts from Actinidia arguta, PG102, attenuates house dust mite-induced murine atopic dermatitis by inhibiting the mTOR pathway with Treg generation. J. Ethnopharmacol. 2016, 193, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, S.H.; Park, E.J.; Kang, C.Y.; Cho, S.H.; Kim, S. Anti-allergic effects of PG102, a water-soluble extract prepared from Actinidia arguta, in a murine ovalbumin-induced asthma model. Clin. Exp. Allergy 2009, 39, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Kim, S.H.; Park, E.J.; Kim, J.; Cho, S.H.; Kagawa, J.; Arai, N.; Jun, K.; Kiyono, H.; Kim, S. Suppression of allergic diarrhea in murine ovalbumin-induced allergic diarrhea model by PG102, a water-soluble extract prepared from Actinidia arguta. Int. Arch. Allergy Immunol. 2009, 150, 164–171. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, B.; Eo, H.; Park, K.; Kim, Y.; Lee, H.J.; Son, M.; Chang, Y.S.; Cho, S.H.; Kim, S.; et al. Control of IgE and selective T(H)1 and T(H)2 cytokines by PG102 isolated from Actinidia arguta. J. Allergy Clin. Immunol. 2005, 116, 1151–1157. [Google Scholar] [CrossRef]
- Park, E.J.; Park, K.C.; Eo, H.; Seo, J.; Son, M.; Kim, K.H.; Chang, Y.S.; Cho, S.H.; Min, K.U.; Jin, M.; et al. Suppression of spontaneous dermatitis in NC/Nga murine model by PG102 isolated from Actinidia arguta. J. Investig. Dermatol. 2007, 127, 1154–1160. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, S.; Lee, S.H.; Park, H.W.; Chang, Y.S.; Min, K.U.; Cho, S.H. The effects of PG102, a water-soluble extract from Actinidia arguta, on serum total IgE levels: A double-blind, randomized, placebo-controlled exploratory clinical study. Eur. J. Nutr. 2011, 50, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Jeon, I.H.; Kim, H.S.; Kang, H.J.; Lee, H.S.; Jeong, S.I.; Kim, S.J.; Jang, S.I. Anti-inflammatory and antipruritic effects of luteolin from Perilla (P. frutescens L.) leaves. Molecules 2014, 19, 6941–6951. [Google Scholar] [CrossRef] [PubMed]
- Okawa, T.; Yamaguchi, Y.; Takada, S.; Sakai, Y.; Numata, N.; Nakamura, F.; Nagashima, Y.; Ikezawa, Z.; Aihara, M. Oral administration of collagen tripeptide improves dryness and pruritus in the acetone-induced dry skin model. J. Dermatol. Sci. 2012, 66, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Ko, K.R.; Kim, H.K.; Lee, D.S.; Nam, I.J.; Lim, S.; Kim, S. Dehydrodiconiferyl Alcohol Inhibits Osteoclast Differentiation and Ovariectomy-Induced Bone Loss through Acting as an Estrogen Receptor Agonist. J. Nat. Prod. 2018, 81, 1343–1356. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Ko, K.R.; Kim, H.K.; Lim, S.; Kim, S. Dehydrodiconiferyl alcohol promotes BMP-2-induced osteoblastogenesis through its agonistic effects on estrogen receptor. Biochem. Biophys. Res. Commun. 2018, 495, 2242–2248. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, M.; Ozawa, S.; Tengara, S.; Ogawa, H.; Takamori, K. Intraepidermal nerve fibers increase in dry skin of acetone-treated mice. J. Dermatol. Sci. 2007, 48, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Zaniboni, M.C.; Samorano, L.P.; Orfali, R.L.; Aoki, V. Skin barrier in atopic dermatitis: Beyond filaggrin. An. Bras. Dermatol. 2016, 91, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Stawczyk-Macieja, M.; Szczerkowska-Dobosz, A.; Rebala, K.; Purzycka-Bohdan, D. Genetic background of skin barrier dysfunction in the pathogenesis of psoriasis vulgaris. Postep. Dermatol. Alergol. 2015, 32, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yosipovitch, G.; Duque, M.I.; Patel, T.S.; Ishiuji, Y.; Guzman-Sanchez, D.A.; Dawn, A.G.; Freedman, B.I.; Chan, Y.H.; Crumrine, D.; Elias, P.M. Skin barrier structure and function and their relationship to pruritus in end-stage renal disease. Nephrol. Dial. Transpl. 2007, 22, 3268–3272. [Google Scholar] [CrossRef] [Green Version]
- Lavery, M.J.; Kinney, M.O.; Mochizuki, H.; Craig, J.; Yosipovitch, G. Pruritus: An overview. What drives people to scratch an itch? Ulster Med. J. 2016, 85, 164–173. [Google Scholar]
- Cabanillas, B.; Novak, N. Atopic dermatitis and filaggrin. Curr. Opin. Immunol. 2016, 42, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Xiong, Z.; Xu, X.; Li, F.; Lu, L.; Li, W.; Su, J.; Liu, Y.; Liu, D.; Xie, Z.; et al. Loss-of-function mutations in filaggrin gene associate with psoriasis vulgaris in Chinese population. Hum. Genet. 2012, 131, 1269–1274. [Google Scholar] [CrossRef] [PubMed]
- Sutter, C.H.; Bodreddigari, S.; Campion, C.; Wible, R.S.; Sutter, T.R. 2,3,7,8-Tetrachlorodibenzo-p-dioxin increases the expression of genes in the human epidermal differentiation complex and accelerates epidermal barrier formation. Toxicol. Sci. 2011, 124, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Wallmeyer, L.; Lehnen, D.; Eger, N.; Sochorova, M.; Opalka, L.; Kovacik, A.; Vavrova, K.; Hedtrich, S. Stimulation of PPARalpha normalizes the skin lipid ratio and improves the skin barrier of normal and filaggrin deficient reconstructed skin. J. Dermatol. Sci. 2015, 80, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Takamori, A.; Nambu, A.; Sato, K.; Yamaguchi, S.; Matsuda, K.; Numata, T.; Sugawara, T.; Yoshizaki, T.; Arae, K.; Morita, H.; et al. IL-31 is crucial for induction of pruritus, but not inflammation, in contact hypersensitivity. Sci. Rep. 2018, 8, 6639. [Google Scholar] [CrossRef]
- Feld, M.; Garcia, R.; Buddenkotte, J.; Katayama, S.; Lewis, K.; Muirhead, G.; Hevezi, P.; Plesser, K.; Schrumpf, H.; Krjutskov, K.; et al. The pruritus- and TH2-associated cytokine IL-31 promotes growth of sensory nerves. J. Allergy Clin. Immunol. 2016, 138, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Egeberg, A.; Thyssen, J.P.; Wu, J.J. Anti-Interleukin-31 Receptor A Antibody for Atopic Dermatitis. N. Engl. J. Med. 2017, 376, 2092–2093. [Google Scholar] [CrossRef]
- Neis, M.M.; Peters, B.; Dreuw, A.; Wenzel, J.; Bieber, T.; Mauch, C.; Krieg, T.; Stanzel, S.; Heinrich, P.C.; Merk, H.F.; et al. Enhanced expression levels of IL-31 correlate with IL-4 and IL-13 in atopic and allergic contact dermatitis. J. Allergy Clin. Immunol. 2006, 118, 930–937. [Google Scholar] [CrossRef]
- Latocha, P. The Nutritional and Health Benefits of Kiwiberry (Actinidia arguta)—A Review. Plant Foods Hum. Nutr. 2017, 72, 325–334. [Google Scholar] [CrossRef]
- Ahmed, H.M. Ethnomedicinal, Phytochemical and Pharmacological Investigations of Perilla frutescens (L.) Britt. Molecules 2018, 24, 102. [Google Scholar] [CrossRef]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, W.; Jeong, Y.; Park, J.-H.; Lee, C.H.; Yun, N.; Lee, D.S.; Nam, I.-J.; Kim, J.-D.; Yoon, K.D.; Son, M.; et al. Water-Soluble Extract from Actinidia arguta (Siebold & Zucc.) Planch. ex Miq. and Perilla frutescens (L.) Britton, ACTPER, Ameliorates a Dry Skin-Induced Itch in a Mice Model and Promotes Filaggrin Expression by Activating the AhR Signaling in HaCaT Cells. Nutrients 2019, 11, 1366. https://doi.org/10.3390/nu11061366
Lee W, Jeong Y, Park J-H, Lee CH, Yun N, Lee DS, Nam I-J, Kim J-D, Yoon KD, Son M, et al. Water-Soluble Extract from Actinidia arguta (Siebold & Zucc.) Planch. ex Miq. and Perilla frutescens (L.) Britton, ACTPER, Ameliorates a Dry Skin-Induced Itch in a Mice Model and Promotes Filaggrin Expression by Activating the AhR Signaling in HaCaT Cells. Nutrients. 2019; 11(6):1366. https://doi.org/10.3390/nu11061366
Chicago/Turabian StyleLee, Wonwoo, Yoonseon Jeong, Jong-Hyung Park, Chang Hyung Lee, Nayoung Yun, Doo Suk Lee, In-Jeong Nam, Jung-Dong Kim, Kee Dong Yoon, Miwon Son, and et al. 2019. "Water-Soluble Extract from Actinidia arguta (Siebold & Zucc.) Planch. ex Miq. and Perilla frutescens (L.) Britton, ACTPER, Ameliorates a Dry Skin-Induced Itch in a Mice Model and Promotes Filaggrin Expression by Activating the AhR Signaling in HaCaT Cells" Nutrients 11, no. 6: 1366. https://doi.org/10.3390/nu11061366