Supplementation of Juçara Berry (Euterpe edulis Mart.) Modulates Epigenetic Markers in Monocytes from Obese Adults: A Double-Blind Randomized Trial

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Participants
2.3. Anthropometric Measurements
2.4. Supplementation
2.5. Dietary Intake
2.6. Sample Collection
2.7. Serum Fatty Acids Analyses by GC/FID
2.8. Total DNA Methylation and HDAC Enzymatic Activity
2.9. RNA Extraction and RT-PCR
2.10. Western Blotting
2.11. Statistical Analyzes
3. Results
3.1. Dietary Intake and Anthropometric Measurements
3.2. Serum Fatty Acids Profile
3.3. Epigenetic Markers Gene Expression
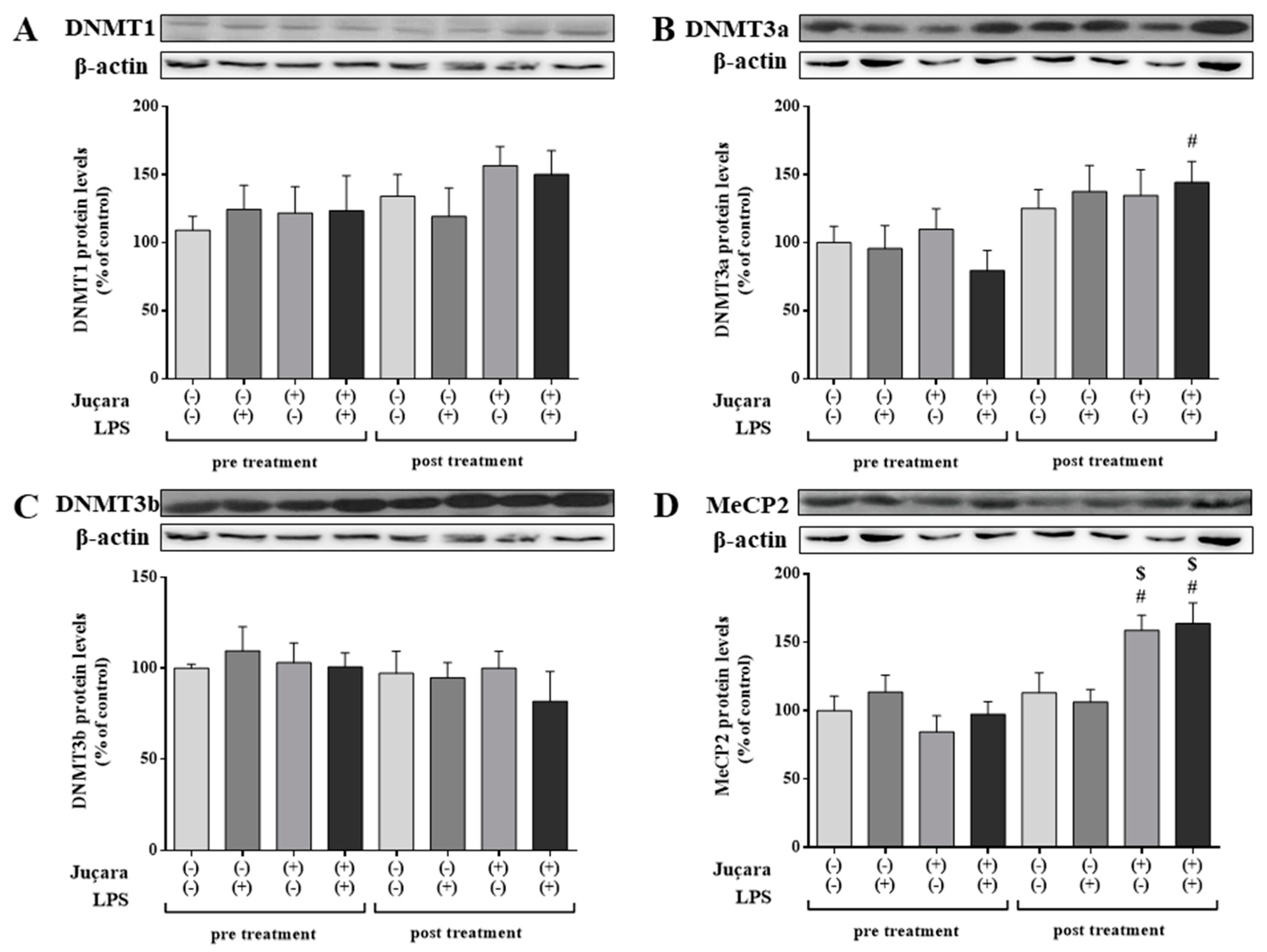
3.4. Epigenetic Markers Protein Expression
3.5. HDAC Activity and Total DNA Methylation
3.6. Serum Fatty Acids Profile Predict Epigenetic Markers Gene Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darnton-Hill, I.; Nishida, C.; James, W. A life course approach to diet, nutrition and the prevention of chronic diseases. Public Health Nutr. 2007, 7, 101–121. [Google Scholar] [CrossRef]
- Delarue, J.; Magnan, C. Free fatty acids and insulin resistance. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Seeley, R.J.; Rushing, P.A.; D’Alessio, D.; Tso, P. A controlled high-fat diet induces an obese syndrome in rats. J. Nutr. 2003, 133, 1081–1087. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Lillycrop, K.A. Fatty acids and epigenetics. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Alfaradhi, M.Z.; Ozanne, S.E. Developmental programming in response to maternal overnutrition. Front. Genet. 2011, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Sloboda, D.M.; Vickers, M.H. Maternal obesity and developmental programming of metabolic disorders in offspring: Evidence from animal models. Exp. Diabetes Res. 2011, 2011. [Google Scholar] [CrossRef]
- Seki, Y.; Williams, L.; Vuguin, P.M.; Charron, M.J. Minireview: Epigenetic Programming of Diabetes and Obesity: Animal Models. Endocrinology 2012, 153, 1031–1038. [Google Scholar] [CrossRef] [Green Version]
- Burdge, G.C.; Hanson, M.A.; Slater-Jefferies, J.L.; Lillycrop, K.A. Epigenetic regulation of transcription: A mechanism for inducing variations in phenotype (fetal programming) by differences in nutrition during early life? Br. J. Nutr. 2007, 97, 1036–1046. [Google Scholar] [CrossRef]
- Lee, W.; Lee, S.Y.; Son, Y.-J.; Yun, J.-M. Gallic Acid Decreases Inflammatory Cytokine Secretion Through Histone Acetyltransferase/Histone Deacetylase Regulation in High Glucose-Induced Human Monocytes. J. Med. Food 2015, 18, 793–801. [Google Scholar] [CrossRef]
- Simar, D.; Versteyhe, S.; Donkin, I.; Liu, J.; Hesson, L.; Nylander, V.; Fossum, A.; Barrès, R. DNA methylation is altered in B and NK lymphocytes in obese and type 2 diabetic human. Metab. Clin. Exp. 2014, 63, 1188–1197. [Google Scholar] [CrossRef]
- Zhang, F.F.; Cardarelli, R.; Carroll, J.; Fulda, K.G.; Kaur, M.; Gonzalez, K.; Vishwanatha, J.K.; Santella, R.M.; Morabia, A. Significant differences in global genomic DNA methylation by gender and race/ethnicity in peripheral blood. Epigenetics 2011, 6, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Santa, F.; Narang, V.; Yap, Z.H.; Tusi, B.K.; Burgold, T.; Austenaa, L.; Bucci, G.; Caganova, M.; Notarbartolo, S.; Casola, S.; et al. Jmjd3 contributes to the control of gene expression in LPS-activated macrophages. EMBO J. 2009, 28, 3341–3352. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Sun, L.; Zhu, Z.; Wang, L.; Li, S.; Ye, R.D. Jmjd3-mediated epigenetic regulation of inflammatory cytokine gene expression in serum amyloid A-stimulated macrophages. Cell. Signal. 2014, 26, 1783–1791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rupasinghe, H.P.V.; Sekhon-Loodu, S.; Mantso, T.; Panayiotidis, M.I. Phytochemicals in regulating fatty acid β-oxidation: Potential underlying mechanisms and their involvement in obesity and weight loss. Pharmacol. Ther. 2016, 165, 153–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Martínez, G.A.; Rodríguez-Ríos, D.; Alvarado-Caudillo, Y.; Vaquero, A.; Esteller, M.; Carmona, F.J.; Moran, S.; Nielsen, F.C.; Wickström-Lindholm, M.; Wrobel, K.; et al. Arachidonic and oleic acid exert distinct effects on the DNA methylome. Epigenetics 2016, 11, 321–334. [Google Scholar] [CrossRef]
- Schulz, M.; da Silva Campelo Borges, G.; Gonzaga, L.V.; Oliveira Costa, A.C.; Fett, R. Juçara fruit (Euterpe edulis Mart.): Sustainable exploitation of a source of bioactive compounds. Food Res. Int. 2016, 89, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.; Carmo, L.; Silva, G.; Silveira-diniz, M.; Casemiro, R.; Spoto, M. Physical, Chemical, and Lipid Composition of Juçara (Euterpe Edulis Mart.) Pulp. Braz. J. Food Nutr. 2013, 24, 7–13. [Google Scholar]
- Silva, N.A.D.; Rodrigues, E.; Mercadante, A.Z.; De Rosso, V.V. Phenolic compounds and carotenoids from four fruits native from the Brazilian Atlantic forest. J. Agric. Food Chem. 2014, 62, 5072–5084. [Google Scholar] [CrossRef]
- Oyama, L.M.; Silva, F.P.; Carnier, J.; De Miranda, D.A.; Santamarina, A.B.; Ribeiro, E.B.; Oller Do Nascimento, C.M.; De Rosso, V.V. Jucąra pulp supplementation improves glucose tolerance in mice. Diabetol. Metab. Syndr. 2016, 8, 1–8. [Google Scholar] [CrossRef]
- Jamar, G.; Santamarina, A.B.; Mennitti, L.V.; Cesar, H.d.C.; Oyama, L.M.; de Rosso, V.V.; Pisani, L.P. Bifidobacterium spp. reshaping in the gut microbiota by low dose of juçara supplementation and hypothalamic insulin resistance in Wistar rats. J. Funct. Foods 2018, 46, 212–219. [Google Scholar] [CrossRef]
- Santamarina, A.; Jamar, G.; Mennitti, L.; de Rosso, V.; Cesar, H.; Oyama, L.; Pisani, L. The Use of Juçara (Euterpe edulis Mart.) Supplementation for Suppression of NF-κB Pathway in the Hypothalamus after High-Fat Diet in Wistar Rats. Molecules 2018, 23, 1814. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campión, J.; Milagro, F.I.; Goyenechea, E.; Martínez, J.A. TNF-α promoter methylation as a predictive biomarker for weight-loss response. Obesity 2009, 17, 1293–1297. [Google Scholar] [CrossRef] [PubMed]
- Caris, A.V.; Lira, F.S.; de Mello, M.T.; Oyama, L.M.; dos Santos, R.V.T. Carbohydrate and glutamine supplementation modulates the Th1/Th2 balance after exercise performed at a simulated altitude of 4500 m. Nutrition 2014, 30, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Hardy, O.T.; Kim, A.; Ciccarelli, C.; Hayman, L.L.; Wiecha, J. Monocytes is a Feature of Metabolic Syndrome in Adolescents. Pediatr. Obes. 2013, 8, 1–7. [Google Scholar] [CrossRef]
- Jialal, I.; Kaur, H.; Devaraj, S. Toll-like receptor status in obesity and metabolic syndrome: A translational perspective. J. Clin. Endocrinol. Metab. 2014, 99, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Durrer, C.; Mah, D.; Simtchouk, S.; Little, J.P. One-week high-fat diet leads to reduced toll-like receptor 2 expression and function in young healthy men. Nutr. Res. 2014, 34, 1045–1051. [Google Scholar] [CrossRef]
- Weiterer, S.; Uhle, F.; Lichtenstern, C.; Siegler, B.H.; Bhuju, S.; Jarek, M.; Bartkuhn, M.; Weigand, M.A. Sepsis induces specific changes in histone modification patterns in human monocytes. PLoS ONE 2015, 10, e0124718. [Google Scholar] [CrossRef]
- WHO. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; WHO: Geneva, Switzerland, 2000; Volume 894. [Google Scholar]
- Kamimura, M.; Baxmann, A.; Sampaio, L.; Cuppari, L. Avaliação Nutricional. Nutrição Clínica no Adulto. In Guias de Medicina Ambulatorial e Hospitalar; Ed. Manole: São Paulo, Brazil, 2007; pp. 89–115. [Google Scholar]
- Alberti, K.; Zimmet, P.; Shaw, J. IDF Epidemiology Task Force Consensus Group. The metabolic syndrome: A new worldwide definition. Lancet 2005, 366, 1059–1062. [Google Scholar] [CrossRef]
- Haun, D.R.; Pitanga, F.J.G.; Lessa, I. Razão cintura/estatura comparado a outros indicadores antropométricos de obesidade como preditor de risco coronariano elevado. Revista da Associação Médica Brasileira 2009, 55, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Argentato, P.P.; Morais, C.A.; Santamarina, A.B.; de Cássia César, H.; Estadella, D.; de Rosso, V.V.; Pisani, L.P. Jussara (Euterpe edulis Mart.) supplementation during pregnancy and lactation modulates UCP-1 and inflammation biomarkers induced by trans-fatty acids in the brown adipose tissue of offspring. Clin. Nutr. Exp. 2017, 12, 50–65. [Google Scholar] [CrossRef]
- Morais, C.A.; Oyama, L.M.; de Moura Conrado, R.; de Rosso, V.V.; do Nascimento, C.O.; Pisani, L.P. Polyphenols-rich fruit in maternal diet modulates inflammatory markers and the gut microbiota and improves colonic expression of ZO-1 in offspring. Food Res. Int. 2015, 77, 186–193. [Google Scholar] [CrossRef]
- Christie, W. Chromatpgraphy and Lipids: A Practical Guide; The Oily Press: Bridgwater, Scotlad, 1989; pp. 11–27. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Jamar, G.; Pisani, L.P.; Oyama, L.M.; Belote, C.; Masquio, D.C.L.; Furuya, V.A.; Carvalho-Ferreira, J.P.; Andrade-Silva, S.G.; Dâmaso, A.R.; Caranti, D.A. Is the neck circumference an emergent predictor for inflammatory status in obese adults? Int. J. Clin. Pract. 2013, 67, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Masquio, D.C.L.; de Piano, A.; Campos, R.M.S.; Sanches, P.L.; Corgosinho, F.C.; Carnier, J.; Oyama, L.M.; do Nascimento, C.M.P.O.; de Mello, M.T.; Tufik, S.; Dâmaso, A.R. Saturated fatty acid intake can influence increase in plasminogen activator inhibitor-1 in obese adolescents. Horm. Metab. Res. 2014, 46, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; Gagliardi, A.; Xavier, H.; Magnoni, C.; Cassani, R.; Lottenberg, A.; Casella Filho, A.; Araújo, D.; Cesena, F.; Alves, R.; et al. I Diretriz sobre o consumo de Gorduras e Saúde Cardiovascular. Arquivos Brasileiros de Cardiologia 2013, 100, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Liou, Y.A.; King, D.J.; Zibrik, D.; Innis, S.M. Decreasing linoleic acid with constant α-linolenic acid in dietary fats increases (n-3) eicosapentaenoic acid in plasma phospholipids in healthy men. J. Nutr. 2007, 137, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Babio, N.; Bulló, M.; Salas-Salvadó, J. Mediterranean diet and metabolic syndrome: The evidence. Public Health Nutr. 2009, 12, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, M.C.; Bermudez, B.; Garcia, I.; Lopez, S.; Abia, R.; Muriana, F.J.G.; Montserrat-De La Paz, S. Dietary fatty acids on aortic root calcification in mice with metabolic syndrome. Food Funct. 2017, 8, 1468–1474. [Google Scholar] [CrossRef] [PubMed]
- Razquin, C.; Martinez, J.A.; Martinez-Gonzalez, M.A.; Mitjavila, M.T.; Estruch, R.; Marti, A. A 3 years follow-up of a Mediterranean diet rich in virgin olive oil is associated with high plasma antioxidant capacity and reduced body weight gain. Eur. J. Clin. Nutr. 2009, 63, 1387–1393. [Google Scholar] [CrossRef] [Green Version]
- Riccardi, G.; Vaccaro, O.; Costabile, G.; Rivellese, A.A. How Well Can We Control Dyslipidemias Through Lifestyle Modifications? Curr. Cardiol. Rep. 2016, 18, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine of the National Academies. Dietary Reference Intakes for Energy, Carbohydrates, Fiber, Fat, Protein and Amino Acids; The National Academies Press: Washington, DC, USA, 2002; Chapter 11; pp. 769–845. ISBN 978-0-309-08525-0. [Google Scholar]
- Cintra, D.E.; Ropelle, E.R.; Moraes, J.C.; Pauli, J.R.; Morari, J.; de Souza, C.T.; Grimaldi, R.; Stahl, M.; Carvalheira, J.B.; Saad, M.J.; et al. Unsaturated fatty acids revert diet-induced hypothalamic inflammation in obesity. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Shimpukade, B.; Hudson, B.D.; Hovgaard, C.K.; Milligan, G.; Ulven, T. Discovery of a potent and selective GPR120 agonist. J. Med. Chem. 2012, 55, 4511–4515. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.-H.; Lai, C.-S.; Wu, J.-C.; Ho, C.-T. Epigenetic and disease targets by polyphenols. Curr. Pharm. Des. 2013, 19, 6156–6185. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, J.; Fujii, H.; Ohnuma, K.; Sato, K.; Ito, Y.; Kaji, M.; Sakamoto, E.; Koganei, M.; Sasaki, H.; Nagashima, Y.; et al. Diet-induced adipose tissue inflammation and liver steatosis are prevented by DPP-4 inhibition in diabetic mice. Diabetes 2011, 60, 1246–1257. [Google Scholar] [CrossRef]
- Oliveira, V.; Marinho, R.; Vitorino, D.; Santos, G.A.; Moraes, J.C.; Dragano, N.; Sartori-Cintra, A.; Pereira, L.; Catharino, R.R.; Da Silva, A.S.R.; et al. Diets containing alfa-linolenic (omega-3) or oleic (omega-9) fatty acids rescues obese mice from insulin resistance. Endocrinology 2015, 156, 4033–4046. [Google Scholar] [CrossRef]
- Ju, J.-H.; Yoon, H.-S.; Park, H.-J.; Kim, M.-Y.; Shin, H.-K.; Park, K.-Y.; Yang, J.-O.; Sohn, M.-S.; Do, M.-S. Anti-Obesity and Antioxidative Effects of Purple Sweet Potato Extract in 3T3-L1 Adipocytes In Vitro. J. Med. Food 2011, 14, 1097–1106. [Google Scholar] [CrossRef]
- Garcia-Diaz, D.F.; Johnson, M.H.; de Mejia, E.G. Anthocyanins from Fermented Berry Beverages Inhibit Inflammation-Related Adiposity Response In Vitro. J. Med. Food 2014, 18, 1–8. [Google Scholar] [CrossRef]
- Xu, W.; Zhou, Q.; Yao, Y.; Li, X.; Zhang, J.; Su, G.; Deng, A. Inhibitory effect of Gardenblue blueberry (Vaccinium ashei Reade) anthocyanin extracts on lipopolysaccharide-stimulated. J. Zhejiang Univ. Sci. B 2016, 17, 425–436. [Google Scholar] [CrossRef]
- Mena, P.; Domínguez-Perles, R.; Gironés-Vilaplana, A.; Baenas, N.; García-Viguera, C.; Villaño, D. Flavan-3-ols, anthocyanins, and inflammation. IUBMB Life 2014, 66, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Yoon, Y.; Yoon, H.; Park, H.M.; Song, S.; Yeum, K.J. Dietary anthocyanins against obesity and inflammation. Nutrients 2017, 9, 1089. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.M.; Jones, A.D.; Henagan, T.M. A Review of Mitochondrial-derived Fatty Acids in Epigenetic Regulation of Obesity and Type 2 Diabetes. J. Nutr. Health Food Sci. 2014, 2, 1–4. [Google Scholar] [CrossRef]
- Lyko, F. The DNA methyltransferase family: A versatile toolkit for epigenetic regulation. Nat. Rev. Genet. 2018, 19, 81–92. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Juçara Pulp | Concentration in 100 g of Fresh Matter | References | |
|---|---|---|---|
| Mean | S.E.M. | ||
| Carbohydrates (g) | 28.3 | 3.5 | [18] |
| Proteins (g) | 6.0 | 0.3 | [18] |
| Lipids (g) | 29.2 | 0.9 | [18] |
| Palmitic acid (%) | 34.43 | 3.42 | [17] |
| Stearic acid (%) | 3.01 | 0.30 | [17] |
| SAT (%) | 37.44 | [17] | |
| Palmitoleic acid (%) | 2.61 | 0.26 | [17] |
| Oleic acid (%) | 35.96 | 3.08 | [17] |
| MUFA (%) | 38.57 | [17] | |
| Linoleic acid (%) | 19.18 | 1.89 | [17] |
| Linolenic acid (%) | 0.91 | 0.20 | [17] |
| PUFA (%) | 20.08 | [17] | |
| Fiber (g) | 28.3 | 0.3 | [18] |
| Ashes (g) | 8.8 | 0.8 | [18] |
| Energetic values (kcal) | 400.0 | 23.9 | [18] |
| Cyanidin 3-rutinoside (mg) | 191.0 | 6.5 | [34] |
| Cyanidin 3-glucoside (mg) | 71.4 | 2.1 | [34] |
| Total anthocyanins (mg) | 262.4 | 8.6 | [34] |
| Total phenolic compounds (mg) | 415.1 | 22.3 | [18] |
| Target Genes | Sequences |
|---|---|
| DNMT1 | 5′-TCCTACGCCATGCCCAGTTTG-3′ (sense) |
| 5′-GAAGATGGGCGTCTCATCATCG-3′ (antisense) | |
| DNMT3a | 5′-GCCCATTCGATCTGGTGATTG-3′ (sense) |
| 5′-TCGTAAAGTCCCTTGCGGGC-3′ (antisense) | |
| DNMT3b | 5′-TGTGCAGAGTCCATTGCTGTAGGA-3′ (sense) |
| 5′-GCTTCCGCCAATCACCAAGTCAAA-3′ (antisense) | |
| MeCP2 | 5’-CAGCTCCAACAGGATTCCATGGT-3′ (sense) |
| 5′-TGATGTCTCTGCTTTGCCTGCCT-3′ (antisense) | |
| HPRT | 5′-CCCTGGCGTCGTGATTAGTG-3′ (sense) |
| 5′-TCGAGCAAGACGTTCAGTCC-3′ (antisense) |
| Placebo (n = 14) | Juçara (n = 13) | p | |||
|---|---|---|---|---|---|
| Initial | Final | Initial | Final | ||
| Energy (kcal) | 2051.82 ± 191.14 | 1960.98 ± 130.24 | 1997.01 ± 157.57 | 2107.45 ± 206.35 | 0.947 |
| Carbohydrates (g) | 258.58 ± 24.49 | 226.72 ± 16 | 253.93 ± 24.20 | 272.47 ± 28.86 | 0.740 |
| Carbohydrates (%) | 50.50 ± 1.54 | 46.65 ± 1.67 | 51.13 ± 1.60 | 51.65 ± 1.91 | 0.301 |
| Protein (g) | 82.85 ± 7.36 | 86.85 ± 8.48 | 81.17 ± 6.01 | 79.99 ± 9.33 | 0.710 |
| Protein (%) | 17.14 ± 1.03 | 22.15 ± 3.56 | 17.03 ± 1.10 | 15.17 ± 0.82 | 0.172 |
| Lipids (g) | 75.74 ± 8.96 | 73.86 ± 8.20 | 72.96 ± 6.38 | 77.51 ± 9.39 | 0.908 |
| Lipids (%) | 32.37 ± 1.25 | 33.73 ± 1.58 | 31.86 ± 1.01 | 33.09 ± 1.68 | 0.912 |
| SFA | 19.39 ± 3.05 | 22.07 ± 3.26 | 22.76 ± 2.21 | 22.63 ± 2.91 | 0.544 |
| MUFA | 18.75 ± 3.00 | 22.05 ± 3.44 | 18.53 ± 2.05 | 23.13 ± 3.33 | 0.912 |
| PUFA | 15.11 ± 2.07 | 14.21 ± 1.35 | 13.13 ± 1.97 | 14.67 ± 2.31 | 0.337 |
| Placebo (n = 14) | Juçara (n = 13) | p | |||
|---|---|---|---|---|---|
| Initial | Final | Initial | Final | ||
| Male/Female | 06/08 | 05/08 | - | ||
| Age (years) | 45.07 ± 3.42 | 45.76 ± 2.58 | - | ||
| Stature (m) | 1.65 ± 0.03 | 1.66 ± 0.03 | - | ||
| Body mass (kg) | 92.23 ± 3.29 | 92.91 ± 3.57 | 96.42 ± 4.85 | 96.81 ± 5.09 | 0.700 |
| BMI (kg/m2) | 33.82 ± 0.71 | 34.06 ± 0.81 | 34.63 ± 1.20 | 34.76 ± 1.30 | 0.440 |
| WC (cm) | 102.70 ± 2.66 | 101.54 ± 2.38 | 104.59 ± 2.43 | 104.83 ± 2.85 | 0.608 |
| W/H | 0.62 ± 0.02 | 0.61 ± 0.01 | 0.63 ± 0.02 | 0.63 ± 0.01 | 0.809 |
| Fatty Acids (% of Total Identified) | Placebo (n = 14) | Juçara (n = 13) | p | ||
|---|---|---|---|---|---|
| Initial | Final | Initial | Final | ||
| ΣSFA | 31.23 ± 2.08 | 35.00 ± 1.48 | 32.49 ± 1.03 | 30.21 ± 1.06 $ | 0.032 |
| C14:0 | 0.87 ± 0.13 | 1.42 ± 0.38 | 0.78 ± 0.05 | 0.86 ± 0.15 | 0.604 |
| C16:0 | 19.91 ± 1.09 | 20.53 ± 1.43 | 21.20 ± 0.63 | 19.06 ± 0.87 | 0.494 |
| C17:0 | 1.36 ± 0.28 | 1.78 ± 0.64 | 0.99 ± 0.32 | 0.98 ± 0.12 | 0.985 |
| C18:0 | 7.06 ± 0.66 | 7.77 ± 0.44 | 7.71 ± 0.58 | 6.70 ± 0.61 | 0.680 |
| C20:0 | 1.82 ± 0.57 | 2.34 ± 0.96 | 1.26 ± 0.48 | 1.70 ± 0.51 | 0.193 |
| C24:0 | 0.73 ± 0.15 | 1.17 ± 0.22 | 0.63 ± 0.07 | 0.91 ± 0.23 | 0.223 |
| ΣMUFA | 27.91 ± 3.63 | 22.19 ± 1.23 | 26.17 ± 2.63 | 28.87 ± 2.78 $ | 0.034 |
| C16:1n7 | 2.97 ± 0.30 | 2.30 ± 0.16 | 2.76 ± 0.32 | 3.19 ± 0.35 $ | 0.031 |
| C18:1n9 | 23.29 ± 3.18 | 18.18 ± 1.07 | 21.73 ± 2.07 | 23.69 ± 2.29 $ | 0.049 |
| C18:1n7 | 1.80 ± 0.18 | 1.72 ± 0.11 | 1.99 ± 0.23 | 1.99 ± 0.20 | 0.359 |
| ΣPUFA | 40.91 ± 1.78 | 42.80 ± 1.91 | 41.34 ± 1.94 | 40.87 ± 3.40 | 0.834 |
| C18:2n6 | 23.98 ± 0.96 | 24.28 ± 1.79 | 25.93 ± 1.51 | 20.10 ± 2.13 | 0.831 |
| C18:3n6 | 2.10 ± 0.24 | 2.53 ± 0.37 | 1.70 ± 0.26 | 2.27 ± 0.36 | 0.658 |
| C18:3n4 | 0.49 ± 0.06 | 0.50 ± 0.05 | 1.03 ± 0.41 | 0.48 ± 0.08 | 0.652 |
| C18:4n3 | 0.14 ± 0.04 | 0.17 ± 0.03 | 0.31 ± 0.11 | 0.74 ± 0.33 | 0.783 |
| C20:2n6 | 0.26 ± 0.03 | 0.28 ± 0.02 | 0.35 ± 0.11 | 1.32 ± 0.61 | 0.427 |
| C20:3n6 | 1.13 ± 0.19 | 1.20 ± 0.16 | 1.87 ± 0.59 | 1.33 ± 0.23 | 0.466 |
| C20:4n6 | 4.19 ± 0.57 | 5.93 ± 0.75 | 4.91 ± 0.67 | 4.90 ± 1.11 | 0.356 |
| C20:3n3 | 1.06 ± 0.43 | 0.50 ± 0.16 | 0.67 ± 0.35 | 0.48 ± 0.31 | 0.552 |
| C20:4n3 | 2.10 ± 0.54 | 2.50 ± 1.02 | 1.68 ± 0.51 | 3.08 ± 0.78 | 0.982 |
| C20:5n3 | 0.87 ± 0.29 | 0.44 ± 0.09 | 0.45 ± 0.08 | 1.37 ± 0.77 | 0.496 |
| C21:5n3 | 2.88 ± 0.67 | 2.64 ± 0.97 | 2.07 ± 0.61 | 3.07 ± 1.15 | 0.858 |
| C22:4n6 | 0.19 ± 0.04 | 0.19 ± 0.03 | 0.26 ± 0.05 | 0.27 ± 0.11 | 0.259 |
| C22:5n3 | 0.84 ± 0.46 | 0.51 ± 0.25 | 0.29 ± 0.04 | 0.52 ± 0.18 | 0.815 |
| C22:6n3 | 0.69 ± 0.20 | 1.12 ± 0.25 | 0.88 ± 0.50 | 1.89 ± 0.99 | 0.537 |
| ΣPUFA ω-3 | 9.06 ± 2.27 | 8.39 ± 2.02 | 6.71 ± 1.70 | 10.70 ± 1.59 # | 0.044 |
| ΣPUFA ω-6 | 31.85 ± 1.50 | 34.41 ± 2.54 | 34.63 ± 2.18 | 30.16 ± 2.13 | 0.532 |
| ω-6/ω-3 | 3.51 ± 0.04 | 4.10 ± 0.12 | 5.16 ± 0.03 | 2.18 ± 0.07 # | 0.036 |
| MeCP2 mRNA | |||||
|---|---|---|---|---|---|
| R2 | β | p | 95% CL | VIF | |
| With Juçara § | 0.362 | 0.602 | 0.014 | 0.178 to 1.304 | 1.000 |
| Oleic fatty acid (C18:1n9) | 0.410 | 0.640 | 0.008 | 0.013 to 0.071 | 1.000 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santamarina, A.B.; Jamar, G.; Mennitti, L.V.; De Cássia César, H.; De Rosso, V.V.; Vasconcelos, J.R.; Oyama, L.M.; Pisani, L.P. Supplementation of Juçara Berry (Euterpe edulis Mart.) Modulates Epigenetic Markers in Monocytes from Obese Adults: A Double-Blind Randomized Trial. Nutrients 2018, 10, 1899. https://doi.org/10.3390/nu10121899
Santamarina AB, Jamar G, Mennitti LV, De Cássia César H, De Rosso VV, Vasconcelos JR, Oyama LM, Pisani LP. Supplementation of Juçara Berry (Euterpe edulis Mart.) Modulates Epigenetic Markers in Monocytes from Obese Adults: A Double-Blind Randomized Trial. Nutrients. 2018; 10(12):1899. https://doi.org/10.3390/nu10121899
Chicago/Turabian StyleSantamarina, Aline Boveto, Giovana Jamar, Laís Vales Mennitti, Helena De Cássia César, Verdiana Vera De Rosso, José Ronnie Vasconcelos, Lila Missae Oyama, and Luciana Pellegrini Pisani. 2018. "Supplementation of Juçara Berry (Euterpe edulis Mart.) Modulates Epigenetic Markers in Monocytes from Obese Adults: A Double-Blind Randomized Trial" Nutrients 10, no. 12: 1899. https://doi.org/10.3390/nu10121899