Effect of Nano-Montmorillonite on Osteoblast Differentiation, Mineral Density, and Osteoclast Differentiation in Bone Formation


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Preparation of NM
2.2. Powder X-Ray Diffraction (XRD)
2.3. Scanning Electron Microscopy (Sem) and Energy Dispersive X-Ray Spectroscopy (EDS)
2.4. Particle Size Analysis
2.5. Cell Culture
2.6. Cell Viability Assay
2.7. Analysis of Alkaline Phosphatase (ALP) Activity
2.8. Bone Mineralization Analysis Using Alizarin Red S Staining
2.9. Tartrate-Resistant Acid Phosphatase (Trap) Activity and Staining
2.10. Animals and Diet
2.11. Biochemical Analyses
2.12. Bone Strength Test
2.13. Measurement of Tibial Bone Ca Content
2.14. Micro-Computerized Tomography (Micro-CT)
2.15. Histological Evaluation
2.16. Von Kossa Staining
2.17. Reverse Transcriptase PCR (RT-PCR)
2.18. Immunohistochemical Analysis
2.19. Western Blot Analyses
2.20. Statistical Analyses
3. Results
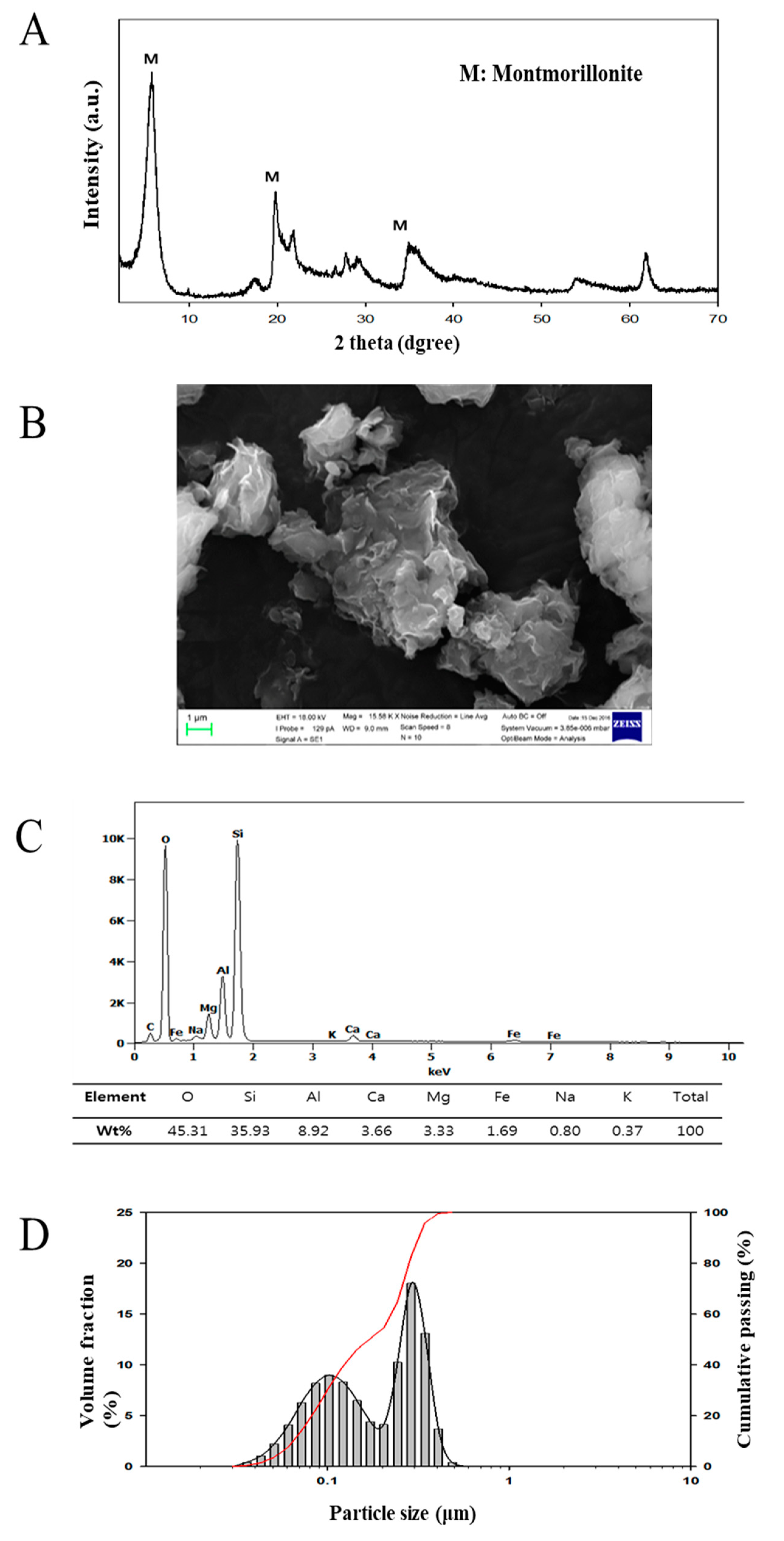
3.1. Characteristics of Montmorillonite
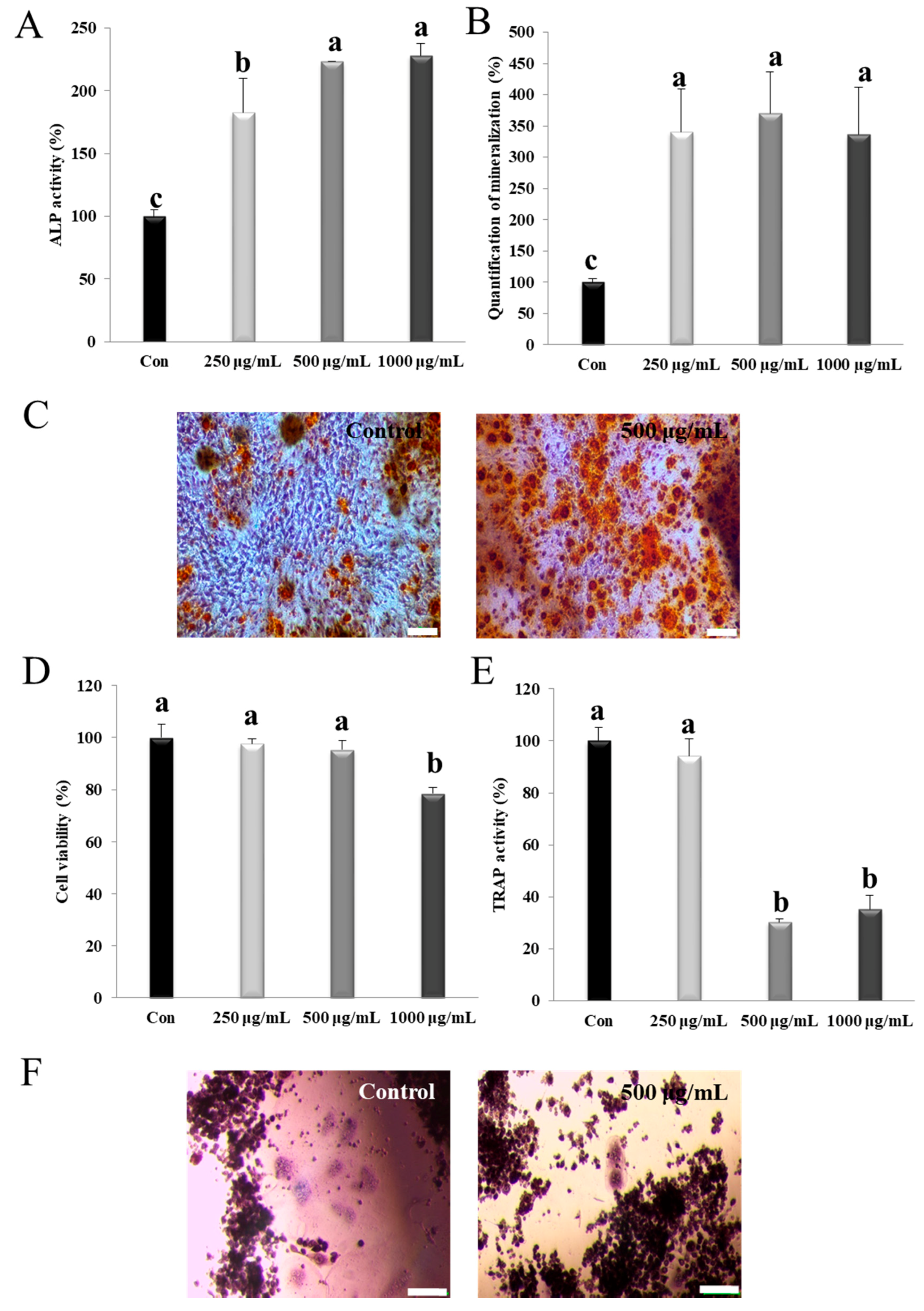
3.2. NM Activates Alp and Mineralization in Osteoblasts
3.3. NM Suppresses Osteoclast Differentiation in Osteoclast Precursor Cells
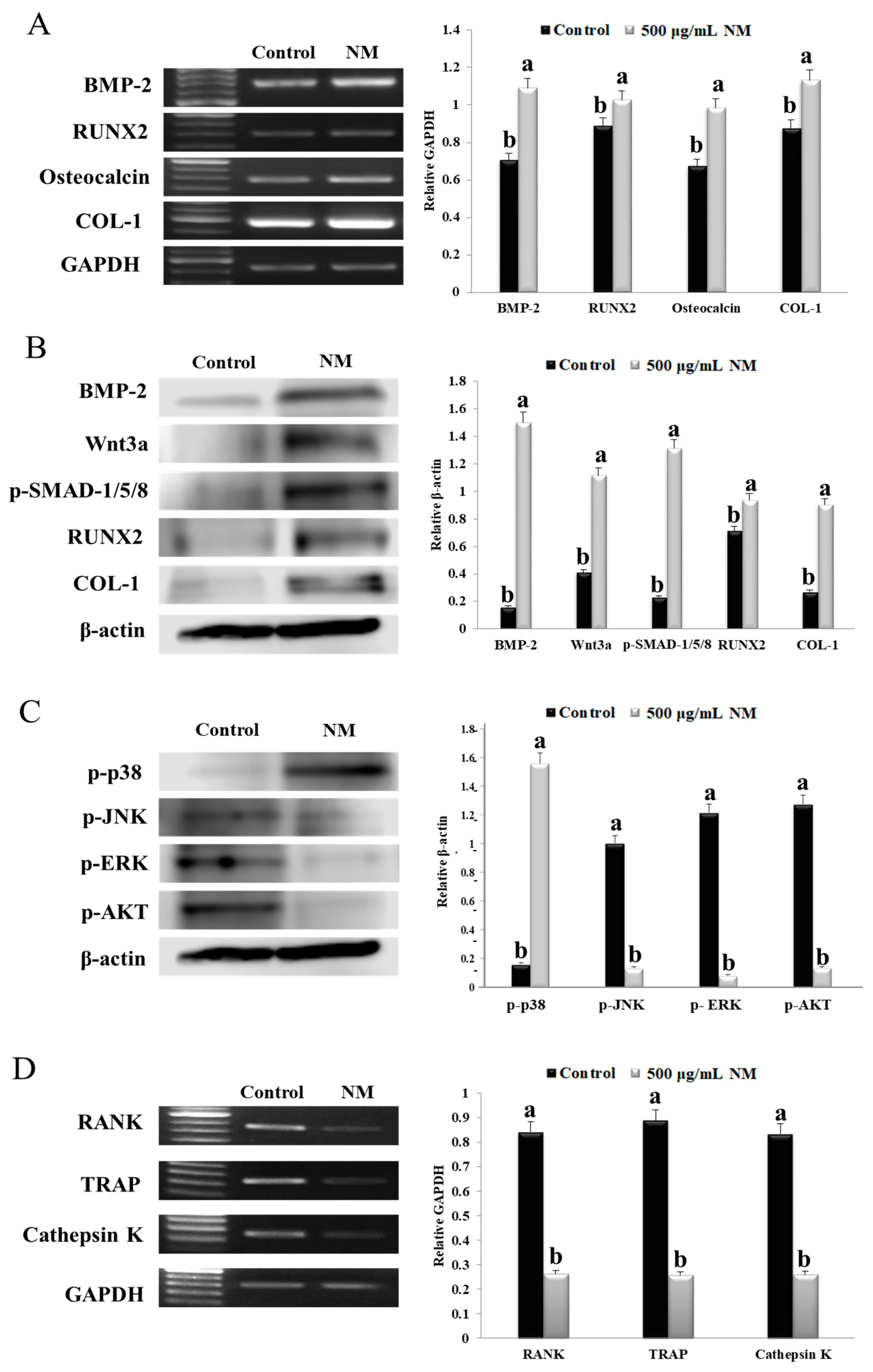
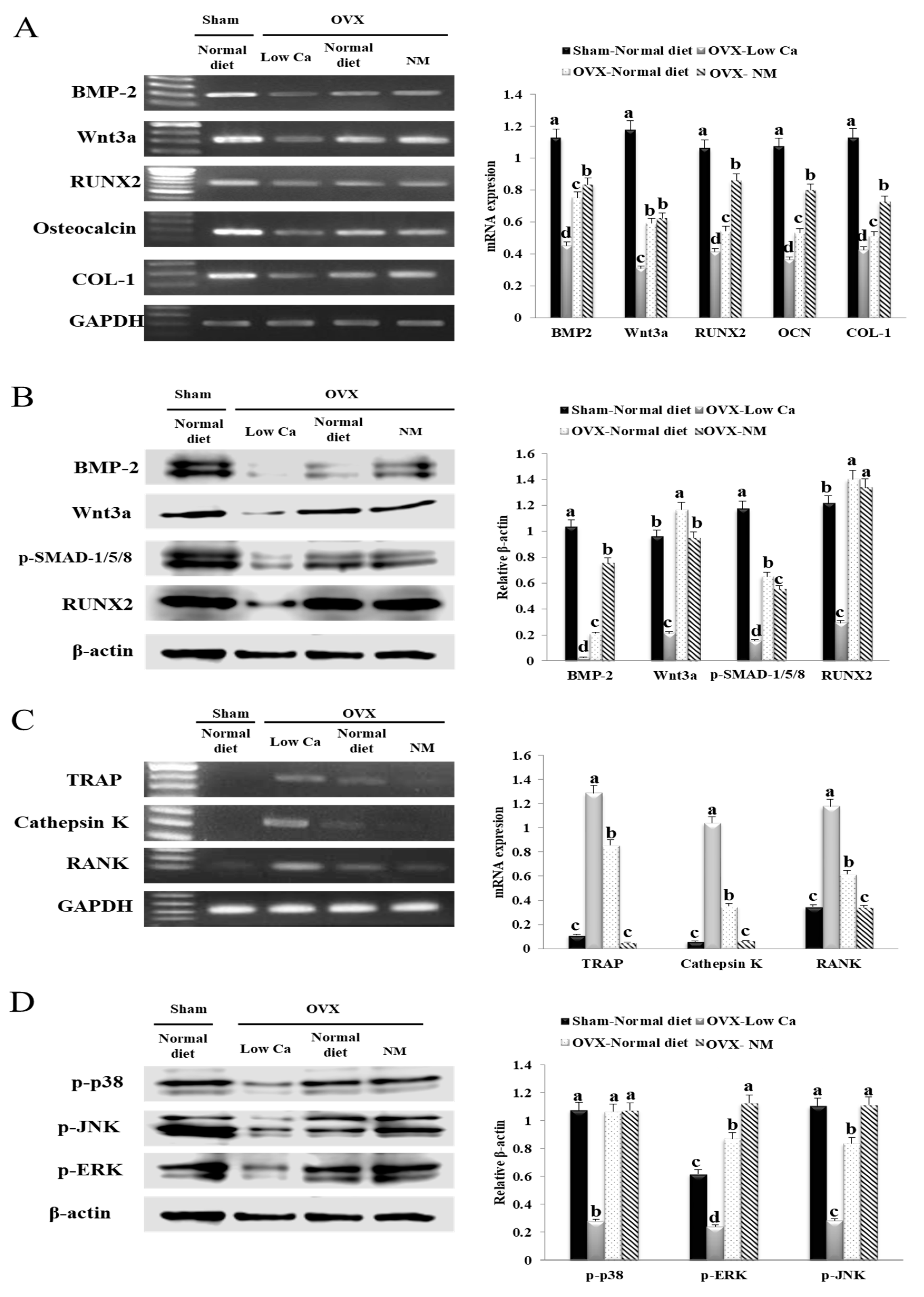
3.4. NM Enhances Expression of Osteoblast Differentiation Marker Genes via P38
3.5. NM Inhibits Osteoclast Formation Genes
3.6. Food Efficiency Ratio
3.7. NM Improves High-Density Lipoprotein (HDL)-Cholesterol Lipid Metabolism and Prevents Atherosclerosis
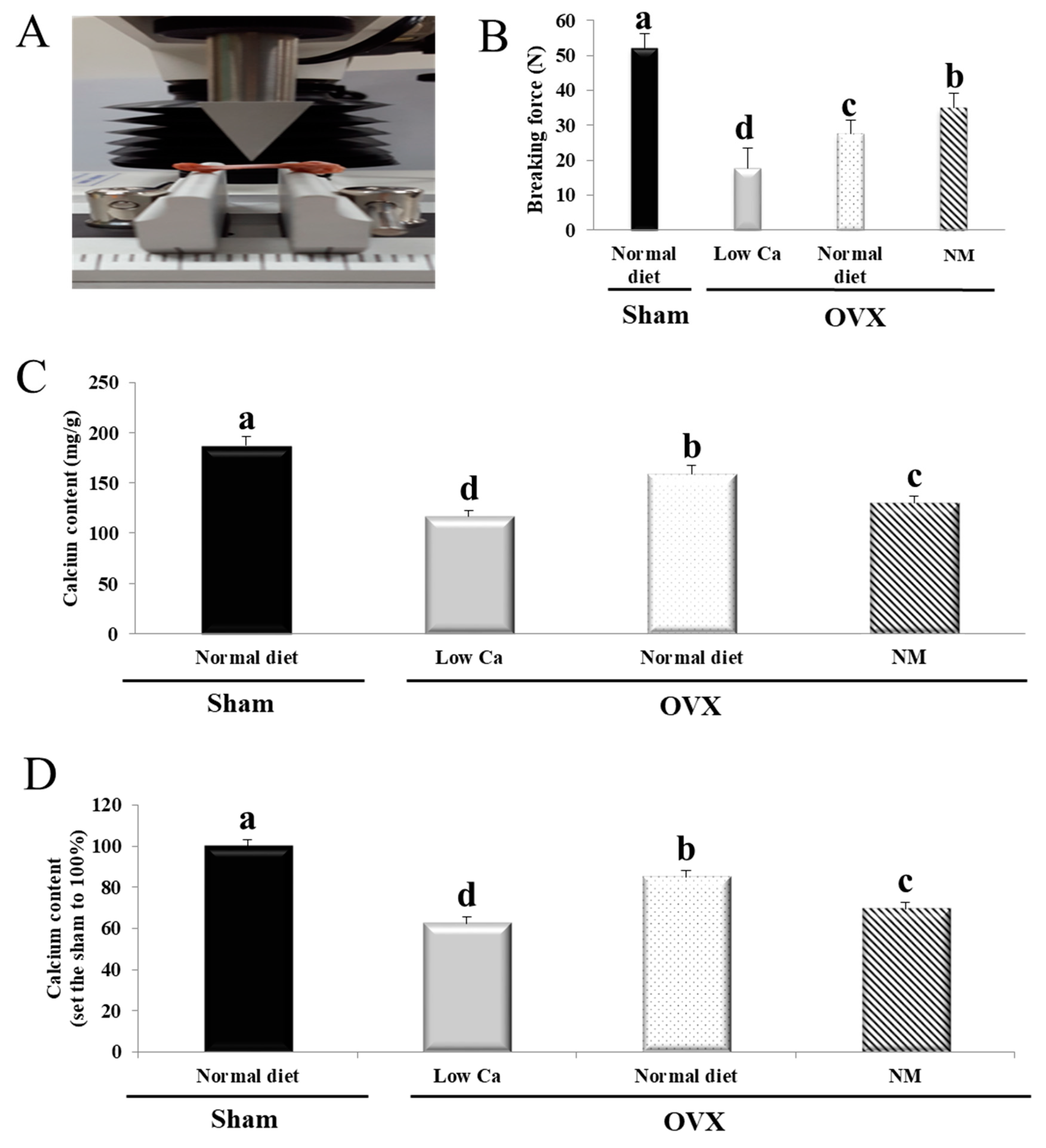
3.8. NM Increases Tibial Ca Content and Bone Strength
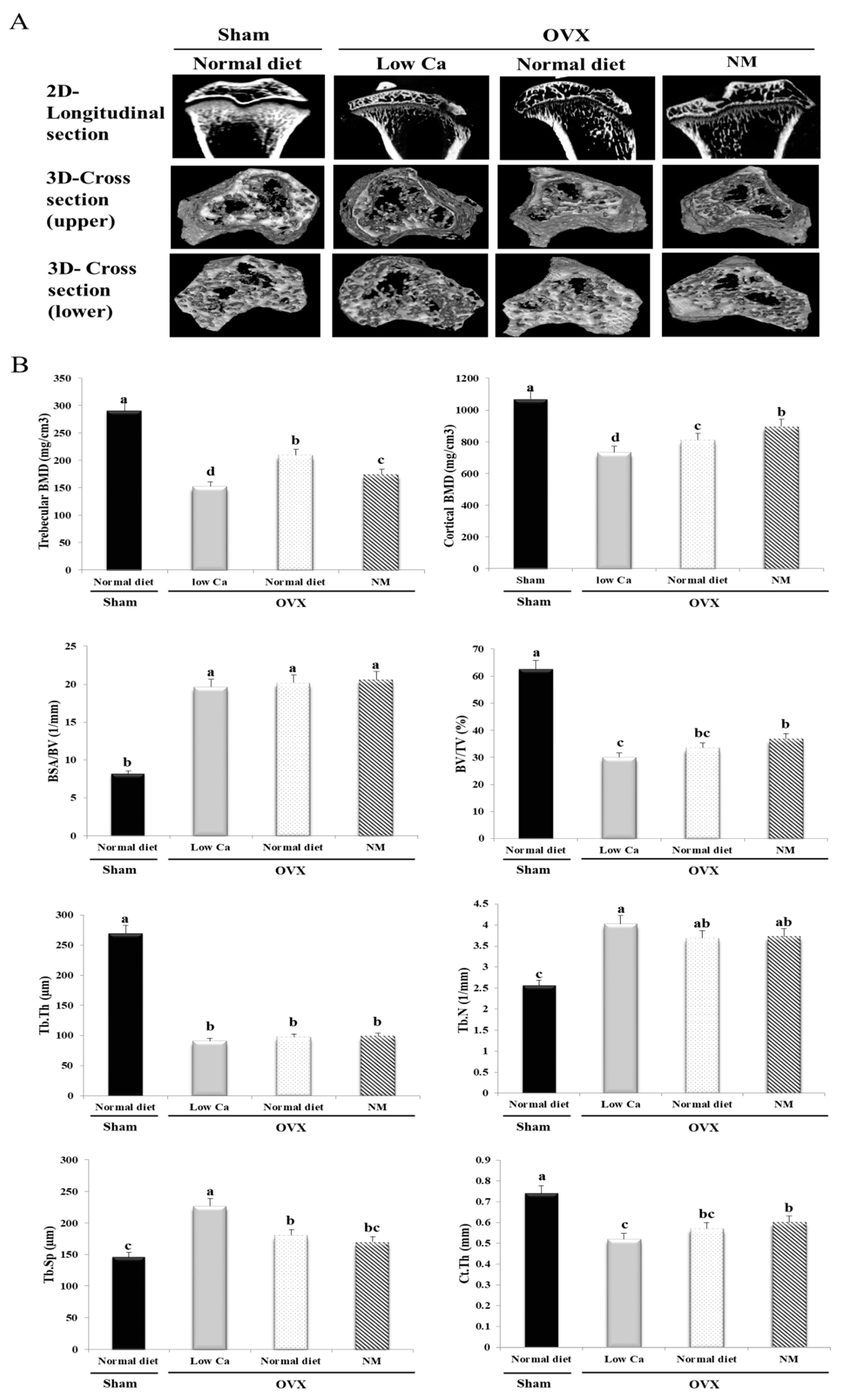
3.9. NM Improves BMD and Bone Microarchitecture
3.10. NM Increases Trabecular Connections
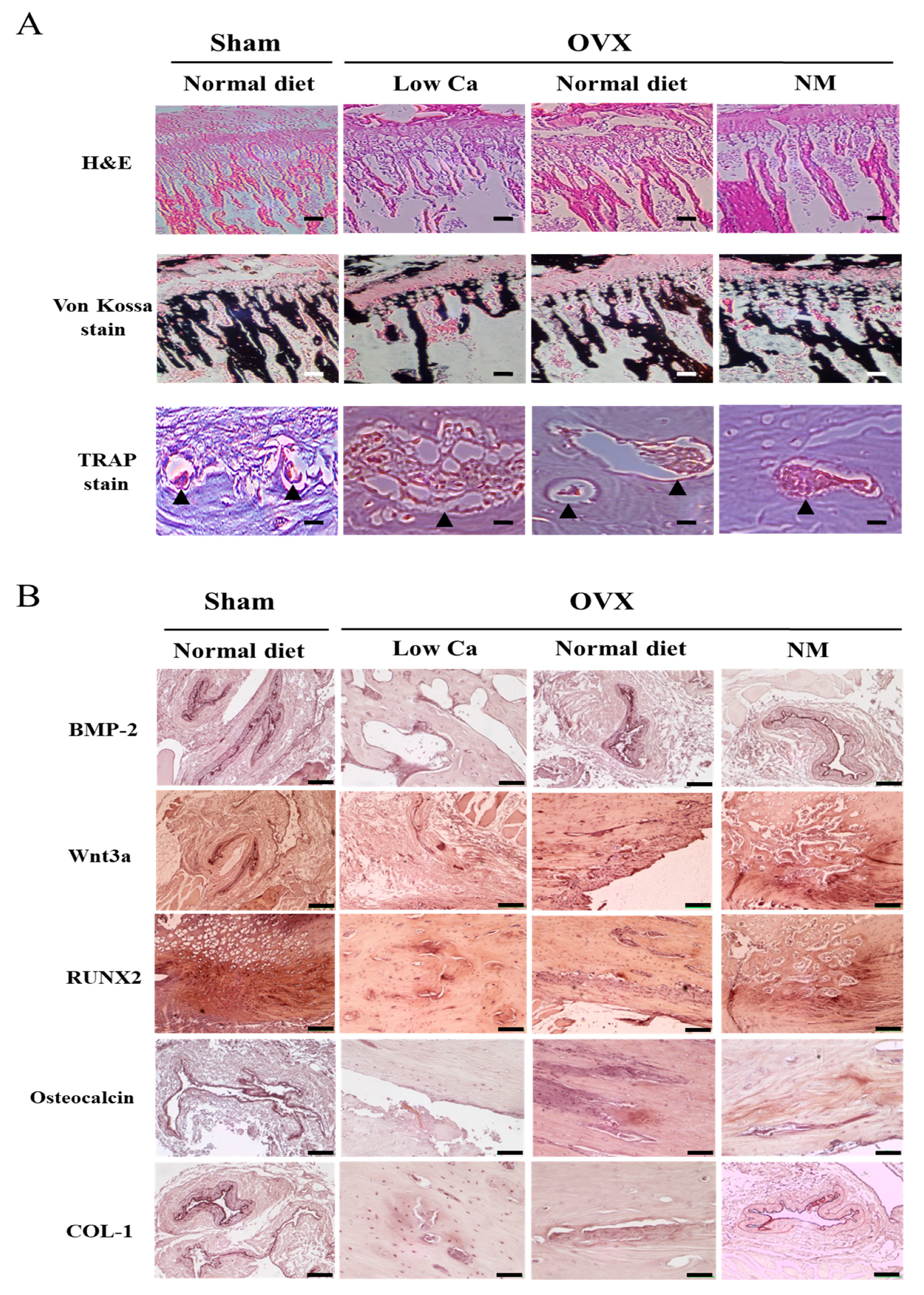
3.11. NM Promotes in Vivo mRNA and Protein Expression of Genes Involved in Osteoblastogenesis
3.12. NM Promotes in Vivo Osteoclastogenesis Related with the mRNA Expression of Genes
3.13. NM Promotes in Vivo Expression of Phosphorylated Mitogen-Activated Protein Kinases (MAPKs) JNK, ERK, and P38
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sawhney, B.L.; Jackson, M.L. Soil montmorillonite formulas. Proc. Soil Sci. Soc. Am. 1958, 22, 115–118. [Google Scholar] [CrossRef]
- Röhl, W.; von Rybinski, W.; Schwuger, M.J. Adsorption of surfactants on low-charged layer silicates. Part I: Adsorption of cationic surfactants. Prog. Colloid Polym. Sci. 1991, 84, 206–214. [Google Scholar]
- Kaufhold, S.; Dohrmann, R.; Ufer, K.; Meyer, F.M. Comparison of methods for the bentonites. Appl. Clay Sci. 2002, 22, 145–151. [Google Scholar] [CrossRef]
- Abdou, M.I.; Al-Sabagh, A.M.; Dardir, M.M. Evaluation of Egyptian bentonite and nano-bentonite as drilling mud. Egyptian J. Petrol. 2013, 22, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Mahto, V.; Sharma, V.P. Rheological study of a water based oil well drilling fluid. J. Pet. Sci. Eng. 2004, 45, 123–128. [Google Scholar] [CrossRef]
- Miles, R.D.; Henry, P.R. Safety of improved Milbond-TX® when fed in broiler diets at greater than recommended levels. Anim. Feed Sci. Technol. 2007, 138, 309–317. [Google Scholar] [CrossRef]
- Peng, X.; Luan, Z.; Zhang, H. Montmorillonite-Cu(II)/Fe(III) oxides magnetic material as adsorbent for removal of humic acid and its thermal regeneration. Chemosphere 2006, 63, 300–306. [Google Scholar] [CrossRef]
- Moraes, J.D.D.; Bertolino, S.R.A.; Cuffini, S.L.; Ducart, D.F.; Bretzke, P.E.; Leonardi, G.R. Clay minerals: Properties and applications to dermocosmetic products and perspectives of natural raw materials for therapeutic purposes-A review. Int. J. Pharm. 2017, 534, 213–219. [Google Scholar] [CrossRef]
- Elmore, A.R. Cosmetic ingredient review expert panel. Final report on the safety assessment of aluminum silicate, calcium silicate, magnesium aluminum silicate, magnesium silicate, magnesium trisilicate, sodium magnesium silicate, zirconium silicate, attapulgite, bentonite, Fuller’s earth, hectorite, kaolin, lithium magnesium silicate, lithium magnesium sodium silicate, montmorillonite, pyrophyllite, and zeolite. Int. J. Toxicol. 2003, 22, 37–102. [Google Scholar]
- Fowler, J.F., Jr. A skin moisturizing cream containing Quaternium-18-Bentonite effectively improves chronic hand dermatitis. J. Cutan. Med. Surg. 2001, 5, 201–205. [Google Scholar] [CrossRef]
- Dardir, F.M.; Mohamed, A.S.; Abukhadra, M.R.; Ahmed, E.A.; Soliman, M.F. Cosmetic and pharmaceutical qualifications of Egyptian bentonite and its suitability as drug carrier for Praziquantel drug. Eur. J. Pharm. Sci. 2018, 115, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Galambos, M.; Suchanek, P.; Rosskopfova, O. Sorption of anthropogenic radionuclides on natural and synthetic inorganic sorbents. J. Radioanal. Nucl. Chem. 2012, 293, 613–633. [Google Scholar] [CrossRef]
- Kahr, G.; Madsen, F.T. Determination of the cation exchange capacity and the surface area of bentonite, illite and kaolinite by methylene blue adsorption. Appl. Clay Sci. 1995, 9, 327–336. [Google Scholar] [CrossRef]
- Lee, J.O.; Lim, J.G.; Kang, I.M.; Kwon, S. Swelling pressures of compacted Ca-bentonite. Eng. Geol. 2012, 129–130, 20–26. [Google Scholar] [CrossRef]
- Babel, S.; Kurniawan, T.A. Low-cost adsorbents for heavy metals uptake from contaminated water: a review. J. Hazard. Mater. 2003, 97, 219–243. [Google Scholar] [CrossRef]
- Vicente-Hernandez, M.A.; Sanchez-Camazano, M.; Sanchez-Martin, M.J. Adsorption and desorption of n-methyl-8-hydroxy quinoline methyl sulfate on smectite and the potential use of the clay-organic product as an ultraviolet radiation collector. Clay Min. 1989, 37, 157–163. [Google Scholar] [CrossRef]
- Song, J.; Li, Y.L.; Hu, C.H. Effects of copper-exchanged montmorillonite, as alternative to antibiotic, on diarrhea, intestinal permeability and proinflammatory cytokine of weanling pigs. Appl. Clay Sci. 2013, 77–78, 52–55. [Google Scholar] [CrossRef]
- Phillips, T.D.; Afriyie-Gyawu, E.; Williams, J.; Huebner, H.; Ankrah, N.A.; Ofori-Adjei, D.; Jolly, P.; Johnson, N.; Taylor, J.; Marroquin-Cardona, A.; et al. Reducing human exposure to aflatoxin through the use of clay: A review. Food Addit. Conta. 2007, 25, 134–145. [Google Scholar] [CrossRef]
- Linares, C.F.; Torres, A. Modified bentonite with Na2CO3 as possible antacid medicament. Stud. Surf. Sci. Catal. 2005, 158, 2073–2080. [Google Scholar]
- Lee, E.S.; Song, E.J.; Lee, S.Y.; Park, S.L.; Kim, D.; Kim, D.; Kim, D.; Kim, J.H.; Lim, S.I.; Nam, Y.D. Effects of bentonite Bgp35b-p on the gut microbiota of mice fed a high-fat diet. J. Sci. Food Agric. 2018, 98, 4369–4373. [Google Scholar] [CrossRef]
- Xu, P.; Dai, S.; Wang, J.; Zhang, J.; Liu, J.; Wang, F.; Zhai, Y. Preventive obesity agent montmorillonite adsorbs dietary lipids and enhances lipid excretion from the digestive tract. Sci. Rep. 2016, 6, 19459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mieszawska, A.J.; Llamas, J.G.; Vaiana, C.A.; Kadakia, M.P.; Naik, R.R.; Kaplan, D.L. Clay enriched silk biomaterials for bone formation. Acta Biomater. 2011, 7, 3036–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katti, K.S.; Katti, D.R.; Dash, R. Synthesis and characterization of a novel chitosan/montmorillonite/hydroxyapatite nanocomposite for bone tissue engineering. Biomed. Mater. 2008, 3, 034122. [Google Scholar] [CrossRef] [PubMed]
- Haroun, A.A.; Gamal-Eldeen, A.; Harding, D.R. Preparation, characterization and in vitro biological study of biomimetic three-dimensional gelatin–montmorillonite/cellulose scaffold for tissue engineering. J. Mater. Sci. Mater. Med. 2009, 20, 2527–2540. [Google Scholar] [CrossRef]
- Khajuria, D.K.; Razdan, R.; Mahapatra, D.R. Additive effects of zoledronic acid and propranolol on bone density and biochemical markers of bone turnover in osteopenic ovariectomized rats. Rev. Bras. Reumatol. 2015, 55, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Weitzmann, M.N.; Pacifici, R. Estrogen deficiency and bone loss: An inflammatory tale. J. Clin. Investig. 2006, 116, 1186–1194. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Mukwaya, E.; Pan, H.; Li, X.M.; Yang, J.L.; Ge, J.; Wang, H.Y. Combination therapy of Chinese herbal medicine Fructus Ligustri Lucidi with high calcium diet on calcium imbalance induced by ovariectomy in mice. Pharm. Biol. 2015, 53, 1082–1085. [Google Scholar] [CrossRef]
- Han, N.R.; Kim, H.Y.; Yang, W.M.; Jeong, H.J.; Kim, H.M. Glutamic acid ameliorates estrogen deficiency–induced menopausal-like symptoms in ovariectomized mice. Nutr. Res. 2015, 35, 774–783. [Google Scholar] [CrossRef]
- Pravina, P.; Sayaji, D.; Avinash, M. Calcium and its role in human body. Int. J. Res. Pharm. Biomed. Sci. 2013, 4, 659–668. [Google Scholar]
- Divittorio, G.; Jackson, K.L.; Chindalore, V.L.; Welker, W.; Walker, J.B. Examining the relationship between bone mineral density and fracture risk reduction during pharmacologic treatment of osteoporosis. Pharmacotherapy 2006, 26, 104–114. [Google Scholar] [CrossRef]
- Durão, S.F.; Gomes, P.S.; Colaço, B.J.; Silva, J.C.; Fonseca, H.M.; Duarte, J.R.; Felino, A.C.; Fernandes, M.H. The biomaterial-mediated healing of critical size bone defects in the ovariectomized rat. Osteoporos. Int. 2014, 25, 1535–1545. [Google Scholar] [CrossRef] [PubMed]
- Heaney, R.P.; Dowell, M.S.; Barger-Lux, M.J. Absorption of calcium as the carbonate and citrate salts, with some observations on method. Osteoporos. Int. 1999, 9, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Li, W.; Lu, C.; Liu, J. Preventive and therapeutic effects of calcium lactate on osteoporosis in aged ovariectomized rats. Wei Sheng Yan Jiu 1998, 27, 402–404. [Google Scholar] [PubMed]
- Yoo, H.S.; Kim, G.J.; Song, D.H.; Chung, K.H.; Lee, K.J.; Kim, D.H.; An, J.H. Calcium supplement derived from Gallus gallus domesticus promotes BMP-2/RUNX2/SMAD5 and suppresses TRAP/RANK expression through MAPK signaling activation. Nutrients 2017, 9, 504. [Google Scholar] [CrossRef] [PubMed]
- Anke, M.; Kramer, K.; Glei, M.; Muller, M. Effects of oral bentonite (Fenamin R) on major and trace element incorporation in various tissues and milk of ruminants. 2. Major elements. Mengen und Spurenelemente 1992, 12Arbeitstagung-Leipzig-12/13-Dezember, 435–442. [Google Scholar]
- Dembinski, Z.; Wieckowski, W.; Kulinska, A. Use of bentonite produced in Poland for gastroinal disease of newborn calves. Medycyna Weterynaryjna 1985, 41, 359–362. [Google Scholar]
- Ferrage, E.; Lanson, B.; Sakharov, B.A.; Drits, V.A. Investigation of smectite hydration properties by modeling experimental X-ray diffraction patterns: Part I. Montmorillonite hydration properties. Am. Miner. 2005, 90, 1358–1374. [Google Scholar] [CrossRef] [Green Version]
- Keren, R.; Shainberg, I. Water vapor isotherms and heat of immersion of Na/Ca-montmorillonite systems—I: homoionic clay. Clay Min. 1975, 23, 193–200. [Google Scholar] [CrossRef]
- Viani, A.; Gualtieri, A.F.; Artioli, G. The nature of disorder in montmorillonite by simulation of X-ray powder patterns. Am. Miner. 2002, 87, 966–975. [Google Scholar] [CrossRef]
- Poli, A.L.; Batista, T.; Schmitt, C.C.; Gessner, F.; Neumann, M.G. Effect of sonication on the particle size of montmorillonite clays. J. Colloid. Interf. Sci. 2008, 325, 386–390. [Google Scholar] [CrossRef]
- Llanos, J.H.R.; Tadini, C.C. Preparation and characterization of bio-nanocomposite films based on cassava starch or chitosan, reinforced with montmorillonite or bamboo nanofibers. Int. J. Biol. Macromol. 2018, 107, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Greenblatt, M.B.; Shim, J.H.; Glimcher, L.H. Mitogen-activated protein kinase pathways in osteoblasts. Annu. Rev. Cell Dev. Biol. 2013, 29, 63–79. [Google Scholar] [CrossRef] [PubMed]
- Stein, G.S.; Lian, J.B.; Owen, T.A. Relationship of cell growth to the regulation of tissue-specific gene expression during osteoblast differentiation. FASEB J. 1990, 4, 3111–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, R.T.; Ge, C. Control of the osteoblast lineage by mitogen-activated protein kinase signaling. Curr. Mol. Biol. Rep. 2017, 3, 122–132. [Google Scholar] [CrossRef]
- Gibbs, D.M.R.; Black, C.R.M.; Hulsart-Billstrom, G.; Shi, P.; Scarpa, E.; Oreffo, R.O.C.; Dawson, J.I. Bone induction at physiological doses of BMP through localization by clay nanoparticle gels. Biomaterials 2016, 99, 16–23. [Google Scholar] [CrossRef]
- Nordin, B.E.C.; Need, A.G.; Morris, H.A.; Horowitz, M.; Robertson, W.G. Evidence for a renal calcium leak in postmenopausal women. J. Clin. Endocrinol. Metab. 1991, 72, 401–407. [Google Scholar] [CrossRef]
- O’Brien, F.J.; Brennan, O.; Kennedy, O.D.; Lee, T.C. Microcracks in cortical bone: how do they affect bone biology? Curr. Osteoporos. Rep. 2005, 3, 39–45. [Google Scholar] [CrossRef]
- Hsu, P.Y.; Tsai, M.T.; Wang, S.P.; Chen, Y.J.; Wu, J.; Hsu, J.T. Cortical bone morphological and trabecular bone microarchitectural changes in the mandible and femoral neck of ovariectomized rats. PLoS ONE 2016, 11, e0154367. [Google Scholar] [CrossRef]
- Xia, L.; Yin, Z.; Mao, L.; Wang, X.; Liu, J.; Jiang, X.; Zhang, Z.; Lin, K.; Chang, J.; Fang, B. Akermanite bioceramics promote osteogenesis, angiogenesis and suppress osteoclastogenesis for osteoporotic bone regeneration. Sci. Rep. 2016, 6, 22005. [Google Scholar] [CrossRef]
- Kim, B.S.; Kang, H.J.; Park, J.Y.; Lee, J. Fucoidan promotes osteoblast differentiation via JNK- and ERK-dependent BMP2-Smad 1/5/8 signaling in human mesenchymal stem cells. Exp. Mol. Med. 2015, 47, e128. [Google Scholar] [CrossRef]
- Jeon, E.J.; Lee, D.H.; Kim, Y.J.; Ahn, J.; Kim, M.J.; Hwang, J.T.; Hur, J.; Kim, M.; Jang, Y.J.; Ha, T.Y.; et al. Effects of yuja peel extract and its flavanones on osteopenia in ovariectomized rats and osteoblast differentiation. Mol. Nutr. Food Res. 2016, 60, 2587–2601. [Google Scholar] [CrossRef]
- Das, S.S.; Hussain, K.; Singh, S.; Hussain, A.; Faruk, A.; Tebyetekerwa, M. Laponite-based nanomaterials for biomedical applications: A review. Curr. Pharm. Des. 2019, 25, 424–443. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, J.; Xu, Y.; Qian, S.; Wang, B.; Liu, F.; Liu, X. Biological behavior of osteoblast-like cells on titania and zirconia films deposited by cathodic arc deposition. Biointerphases. 2012, 7, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwaha, P.; Khedgikar, V.; Gautam, J.; Dixit, P.; Chillara, R.; Verma, A.; Thakur, R.; Mishra, D.P.; Singh, D.; Maurya, R.; et al. A novel therapeutic approach with Caviunin-based isoflavonoid that en routes bone marrow cells to bone formation via BMP2/Wnt-β-catenin signaling. Cell Death Dis. 2014, 5, e1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Li, Y.; Zhou, H.; Chen, J.; Chen, M.; Xiao, Z. Genistein promotion of osteogenic differentiation through BMP2/SMAD5/RUNX2 signaling. Int. J. Biol. Sci. 2013, 9, 1089–1098. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [Green Version]
- Gaur, T.; Lengner, C.J.; Hovhannisyan, H.; Bhat, R.A.; Bodine, P.V.; Komm, B.S.; Javed, A.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S.; et al. Canonical WNT signaling promotes osteogenesis by directly stimulating Runx2 gene expression. J. Biol. Chem. 2005, 280, 33132–33140. [Google Scholar] [CrossRef] [Green Version]
- Gori, F.; Hofbauer, L.C.; Dunstan, C.R.; Spelsberg, T.C.; Khosla, S.; Riggs, B.L. The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology 2000, 141, 4768–4776. [Google Scholar] [CrossRef]
- Raggatt, L.J.; Partridge, N.C. Cellular and molecular mechanisms of bone remodeling. J. Biol. Chem. 2010, 285, 25103–25108. [Google Scholar] [CrossRef] [Green Version]
- Zenger, S.; Hollberg, K.; Ljusberg, J.; Norgård, M.; Ek-Rylander, B.; Kiviranta, R.; Andersson, G. Proteolytic processing and polarized secretion of tartrate-resistant acid phosphatase is altered in a subpopulation of metaphyseal osteoclasts in cathepsin K-deficient mice. Bone 2007, 41, 820–832. [Google Scholar] [CrossRef]
- Bossard, M.J.; Tomaszek, T.A.; Thompson, S.K.; Amegadzie, B.Y.; Hanning, C.R.; Jones, C. Proteolytic activity of human osteoclast cathepsin K. Expression, purification, activation, and substrate identification. J. Biol. Chem. 1996, 271, 12517–12524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Kilb, J.; Wex, H.; Li, Z.; Lipyansky, A.; Breuil, V.; Stein, L.; Palmer, J.T.; Dempster, D.W.; Brömme, D. Localization of rat cathepsin K in osteoclasts and resorption pits: inhibition of bone resorption and cathepsin K-activity by peptidyl vinyl sulfones. Biol. Chem. 1999, 380, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Koperski, K.; Schumacher, M.; Gelinsky, M. Relevance of osteoclast-specific enzyme activities in cell-based in vitro resorption assays. Eur. Cells Mater. 2017, 18, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Rubin, J.; Rubin, C.; Jacobs, C.R. Molecular pathways mediating mechanical signaling in bone. Gene 2006, 367, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Eddie, R.C.; Beatriz, G.; Francesc, V. p38 MAPK signaling in osteoblast differentiation. Front. Cell Dev. Biol. 2016, 4, 1–20. [Google Scholar]
- Lin, K.; Xia, L.; Li, H.; Jiang, X.; Pan, H.; Xu, Y.; Lu, W.W.; Zhang, Z.; Chang, J. Enhanced osteoporotic bone regeneration by strontium-substituted calcium silicate bioactive ceramics. Biomaterials 2013, 34, 10028–10042. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.-J.; Kim, D.; Lee, K.-J.; Kim, D.; Chung, K.-H.; Choi, J.W.; An, J.H. Effect of Nano-Montmorillonite on Osteoblast Differentiation, Mineral Density, and Osteoclast Differentiation in Bone Formation. Nanomaterials 2020, 10, 230. https://doi.org/10.3390/nano10020230
Kim G-J, Kim D, Lee K-J, Kim D, Chung K-H, Choi JW, An JH. Effect of Nano-Montmorillonite on Osteoblast Differentiation, Mineral Density, and Osteoclast Differentiation in Bone Formation. Nanomaterials. 2020; 10(2):230. https://doi.org/10.3390/nano10020230
Chicago/Turabian StyleKim, Gyeong-Ji, Daniel Kim, Kwon-Jai Lee, Daeyoung Kim, Kang-Hyun Chung, Jeong Woo Choi, and Jeung Hee An. 2020. "Effect of Nano-Montmorillonite on Osteoblast Differentiation, Mineral Density, and Osteoclast Differentiation in Bone Formation" Nanomaterials 10, no. 2: 230. https://doi.org/10.3390/nano10020230