Fatal Elephant Endotheliotropic Herpesvirus Infection of Two Young Asian Elephants

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
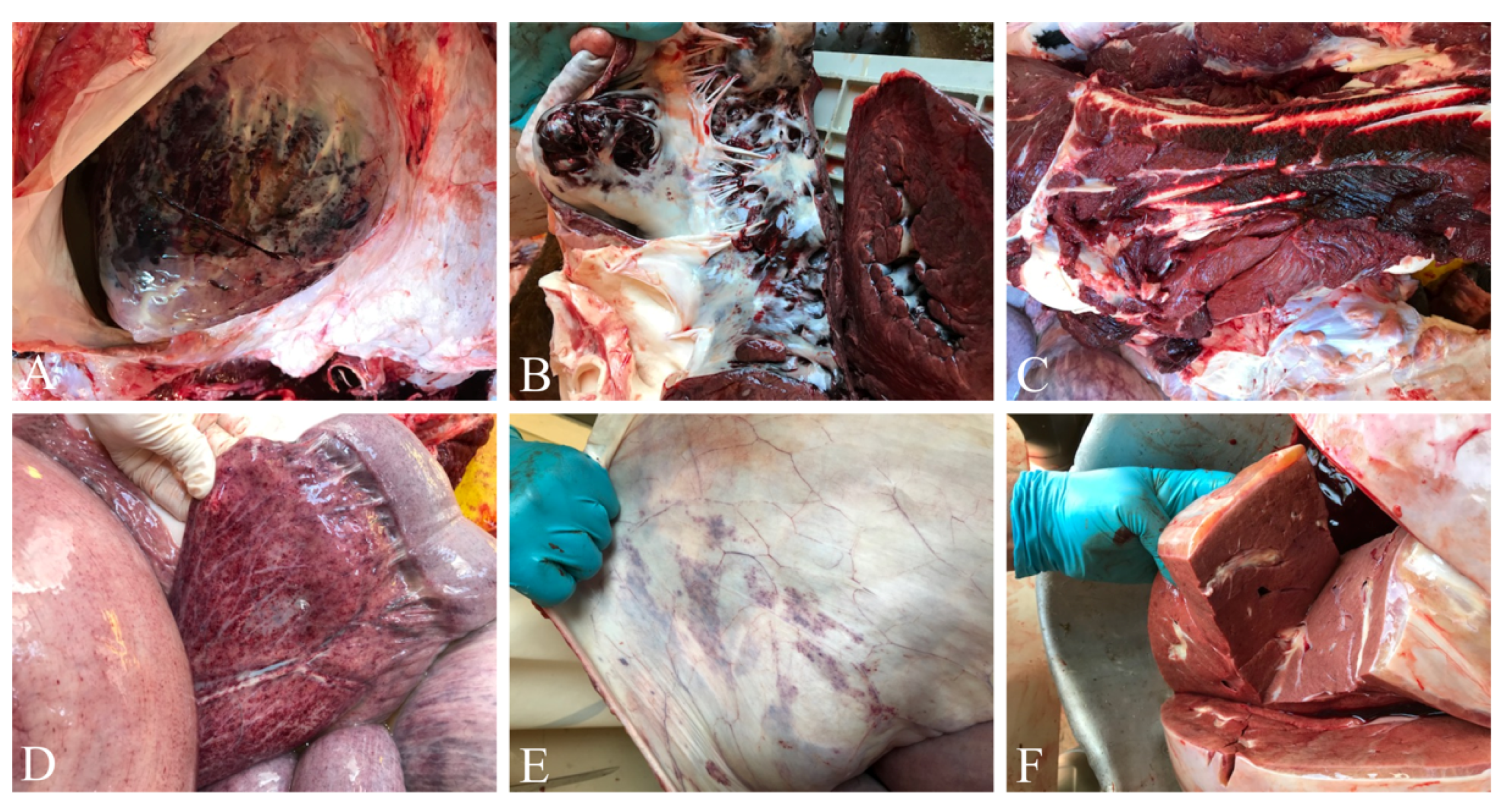
3.1. Hemorrhagic Lesions in All Organs
3.2. Extensive Distribution of Virus in All Organs
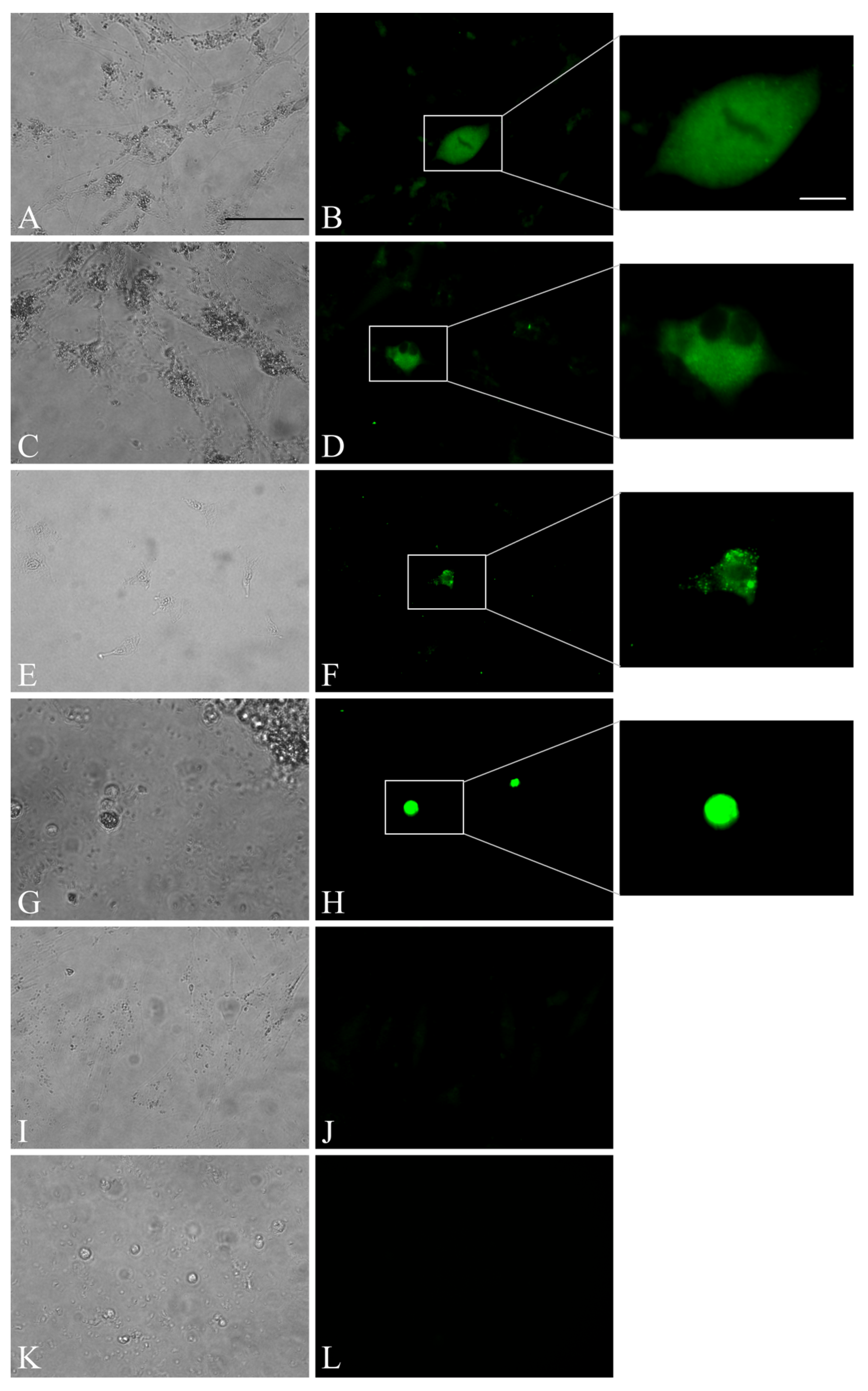
3.3. Trials of Virus Isolation in Cell Culture Revealed Limited Virus Replication
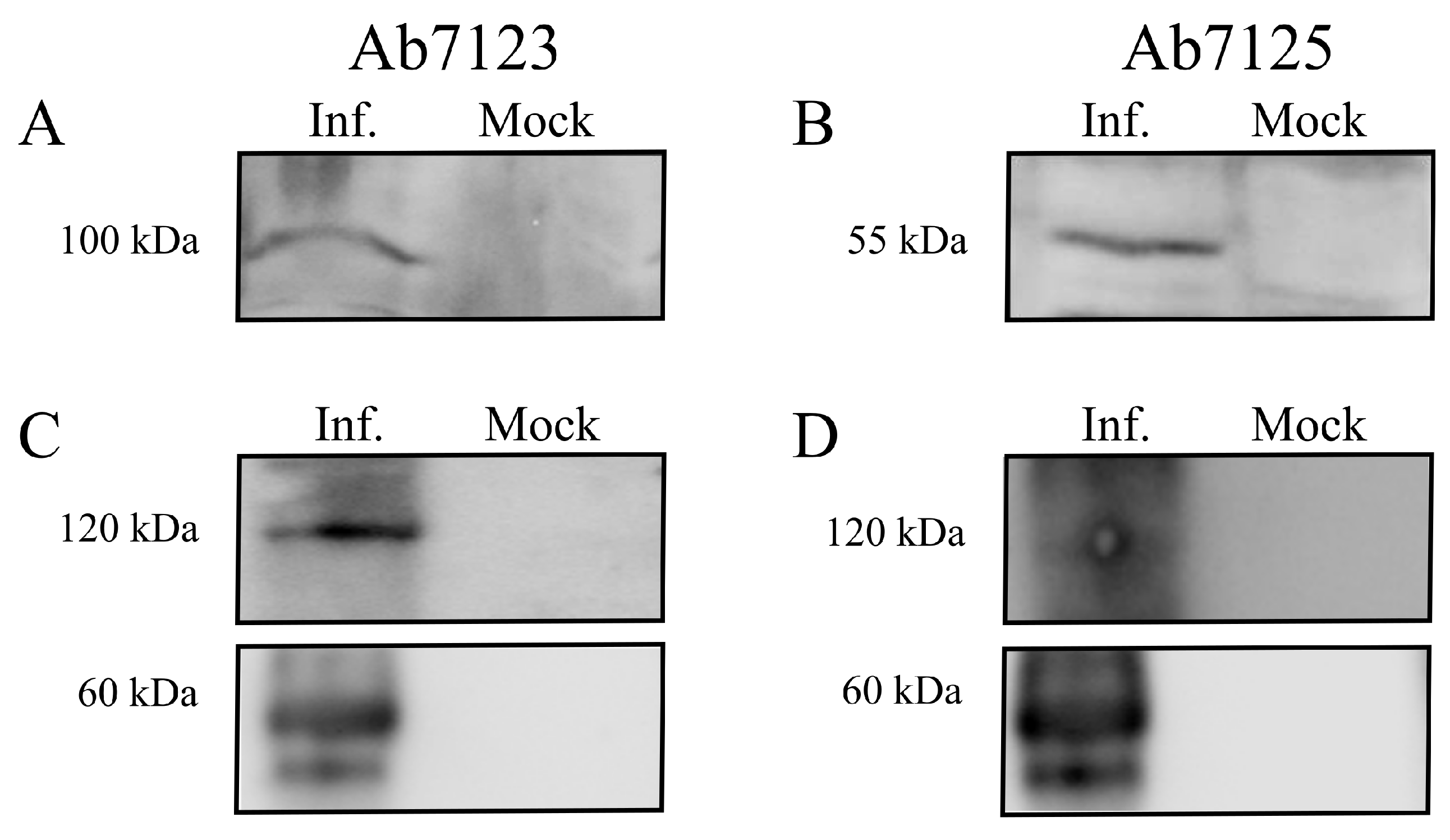
3.4. Expression and Cleavage of gB
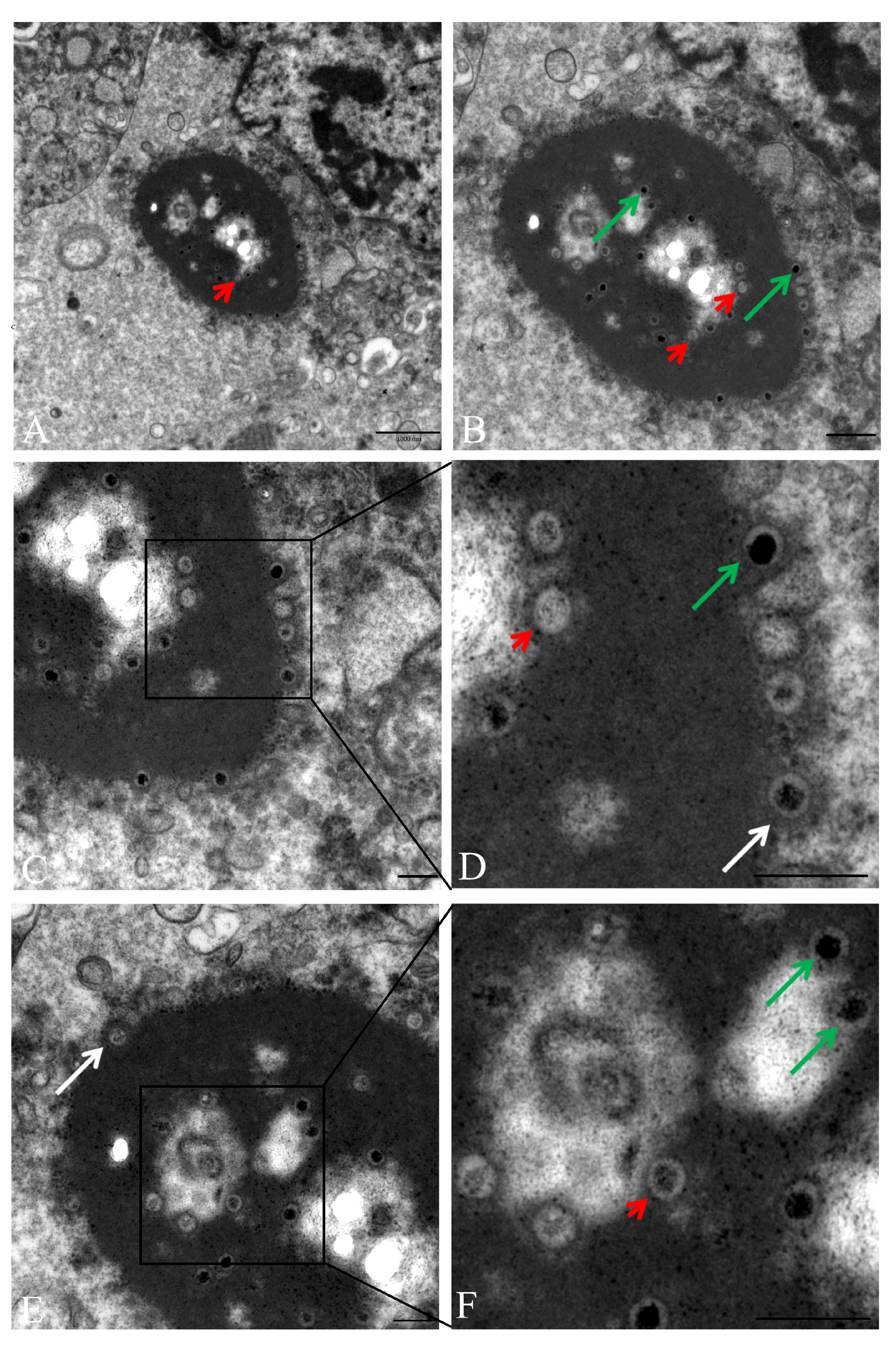
3.5. Transmission Electron Microscopy (TEM) Revealed Intranuclear and Intracytoplasmic Viral Particles
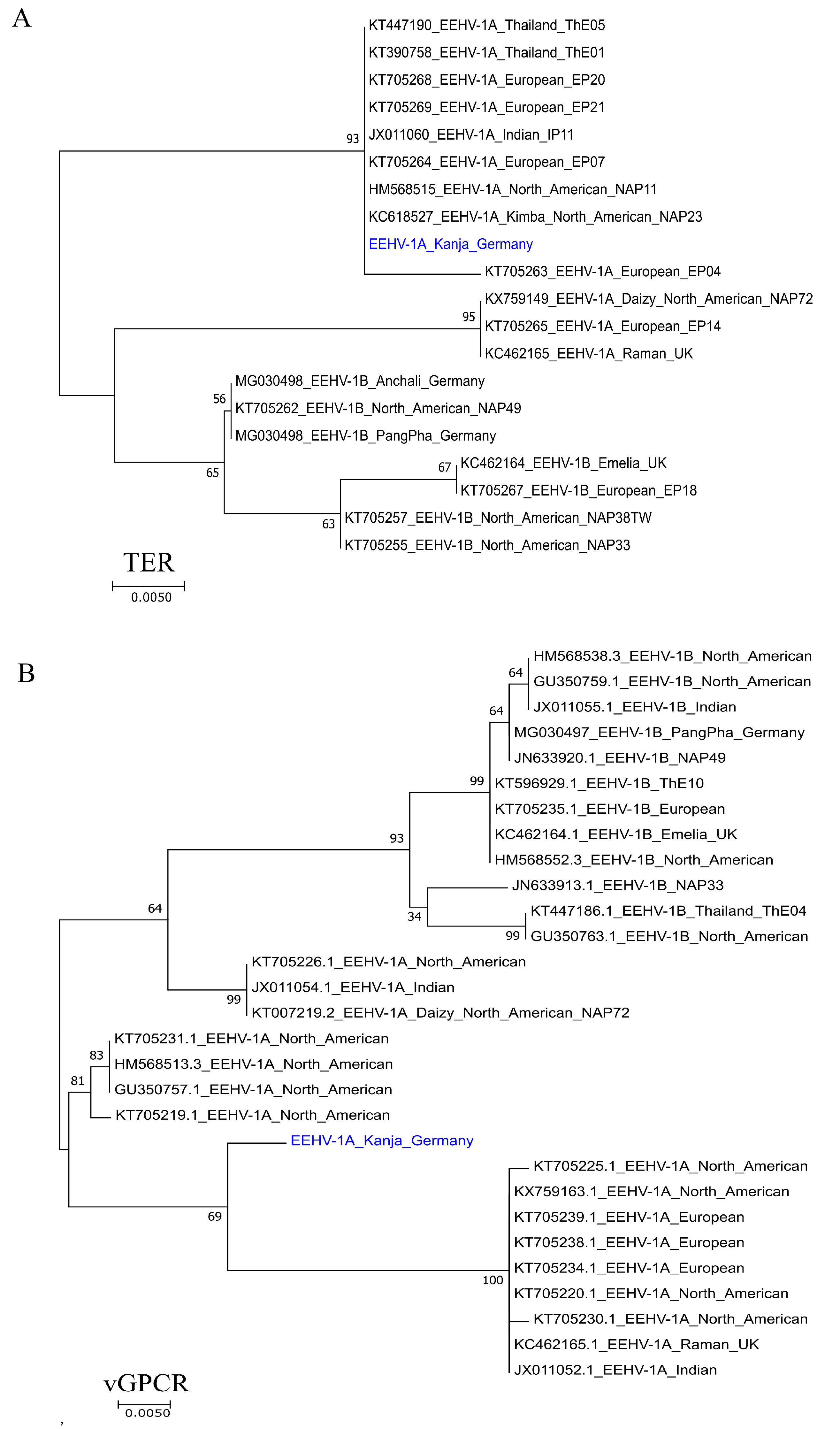
3.6. Whole Genome Sequencing of EEHV-1A
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Richman, L.K.; Montali, R.J.; Garber, R.L.; Kennedy, M.A.; Lehnhardt, J.; Hildebrandt, T.; Schmitt, D.; Hardy, D.; Alcendor, D.J.; Hayward, G.S. Novel endotheliotropic herpesviruses fatal for Asian and African elephants. Science 1999, 283, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.M.; Helmick, K.; Ochsenreiter, J.; Richman, L.K.; Latimer, E.; Wise, A.G.; Maes, R.K.; Kiupel, M.; Nordhausen, R.W.; Zong, J.C.; et al. Clinico-pathologic features of fatal disease attributed to new variants of endotheliotropic herpesviruses in two Asian elephants (Elephas maximus). Vet. Pathol. 2009, 46, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Ossent, P.; Guscetti, F.; Metzler, A.E.; Lang, E.M.; Rubel, A.; Hauser, B. Acute and fatal herpesvirus infection in a young Asian elephant (Elephas maximus). Vet. Pathol. 1990, 27, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Long, S.Y.; Latimer, E.M.; Hayward, G.S. Review of Elephant Endotheliotropic Herpesviruses and Acute Hemorrhagic Disease. ILAR J. 2016, 56, 283–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fickel, J.; Richman, L.K.; Montali, R.; Schaftenaar, W.; Goritz, F.; Hildebrandt, T.B.; Pitra, C. A variant of the endotheliotropic herpesvirus in Asian elephants (Elephas maximus) in European zoos. Vet. Microbiol. 2001, 82, 103–109. [Google Scholar] [CrossRef]
- Zong, J.C.; Latimer, E.M.; Long, S.Y.; Richman, L.K.; Heaggans, S.Y.; Hayward, G.S. Comparative genome analysis of four elephant endotheliotropic herpesviruses, EEHV3, EEHV4, EEHV5, and EEHV6, from cases of hemorrhagic disease or viremia. J. Virol. 2014, 88, 13547–13569. [Google Scholar] [CrossRef] [PubMed]
- Kochagul, V.; Srivorakul, S.; Boonsri, K.; Somgird, C.; Sthitmatee, N.; Thitaram, C.; Pringproa, K. Production of antibody against elephant endotheliotropic herpesvirus (EEHV) unveils tissue tropisms and routes of viral transmission in EEHV-infected Asian elephants. Sci. Rep. 2018, 8, 4675. [Google Scholar] [CrossRef]
- Ackermann, M.; Hatt, J.M.; Schetle, N.; Steinmetz, H. Identification of shedders of elephant endotheliotropic herpesviruses among Asian elephants (Elephas maximus) in Switzerland. PLoS ONE 2017, 12, e0176891. [Google Scholar] [CrossRef]
- Richman, L.K.; Zong, J.C.; Latimer, E.M.; Lock, J.; Fleischer, R.C.; Heaggans, S.Y.; Hayward, G.S. Elephant endotheliotropic herpesviruses EEHV1A, EEHV1B, and EEHV2 from cases of hemorrhagic disease are highly diverged from other mammalian herpesviruses and may form a new subfamily. J. Virol. 2014, 88, 13523–13546. [Google Scholar] [CrossRef]
- Schmitt, D.L.; Hardy, D.A.; Montali, R.J.; Richman, L.K.; Lindsay, W.A.; Isaza, R.; West, G. Use of famciclovir for the treatment of endotheliotrophic herpesvirus infections in Asian elephants (Elephas maximus). J. Zoo Wildl. Med. 2000, 31, 518–522. [Google Scholar] [CrossRef]
- Hardman, K.; Dastjerdi, A.; Gurrala, R.; Routh, A.; Banks, M.; Steinbach, F.; Bouts, T. Detection of elephant endotheliotropic herpesvirus type 1 in asymptomatic elephants using TaqMan real-time PCR. Vet. Rec. 2012, 170, 205. [Google Scholar] [CrossRef] [PubMed]
- Azab, W.; Damiani, A.M.; Ochs, A.; Osterrieder, N. Subclinical infection of a young captive Asian elephant with elephant endotheliotropic herpesvirus 1. Arch. Virol. 2018, 163, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.; Wallaschek, N.; Glass, M.; Flamand, L.; Wight, D.J.; Kaufer, B.B. The ND10 Complex Represses Lytic Human Herpesvirus 6A Replication and Promotes Silencing of the Viral Genome. Viruses 2018, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Spiesschaert, B.; Goldenbogen, B.; Taferner, S.; Schade, M.; Mahmoud, M.; Klipp, E.; Osterrieder, N.; Azab, W. Role of gB and pUS3 in Equine Herpesvirus 1 Transfer between Peripheral Blood Mononuclear Cells and Endothelial Cells: A Dynamic In Vitro Model. J. Virol. 2015, 89, 11899–11908. [Google Scholar] [CrossRef] [PubMed]
- Black, W.; Troyer, R.M.; Coutu, J.; Wong, K.; Wolff, P.; Gilbert, M.; Yuan, J.; Wise, A.G.; Wang, S.; Xu, D.; et al. Identification of gammaherpesvirus infection in free-ranging black bears (Ursus americanus). Virus Res. 2019, 259, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Dayaram, A.; Tsangaras, K.; Pavulraj, S.; Azab, W.; Groenke, N.; Wibbelt, G.; Sicks, F.; Osterrieder, N.; Greenwood, A.D. Novel Divergent Polar Bear-Associated Mastadenovirus Recovered from a Deceased Juvenile Polar Bear. mSphere 2018, 3, e00171. [Google Scholar] [CrossRef] [PubMed]
- Spiesschaert, B.; Stephanowitz, H.; Krause, E.; Osterrieder, N.; Azab, W. Glycoprotein B of equine herpesvirus type 1 has two recognition sites for subtilisin-like proteases that are cleaved by furin. J. Gen. Virol. 2016, 97, 1218–1228. [Google Scholar] [CrossRef]
- Fickel, J.; Lieckfeldt, D.; Richman, L.K.; Streich, W.J.; Hildebrandt, T.B.; Pitra, C. Comparison of glycoprotein B (gB) variants of the elephant endotheliotropic herpesvirus (EEHV) isolated from Asian elephants (Elephas maximus). Vet. Microbiol. 2003, 91, 11–21. [Google Scholar] [CrossRef]
- Niepmann, M.; Zheng, J. Discontinuous native protein gel electrophoresis. Electrophoresis 2006, 27, 3949–3951. [Google Scholar] [CrossRef]
- Watanabe, T.; Nakagawa, T.; Ikemizu, J.; Nagahama, M.; Murakami, K.; Nakayama, K. Sequence requirements for precursor cleavage within the constitutive secretory pathway. J. Biol. Chem. 1992, 267, 8270–8274. [Google Scholar]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Raney, B.; Paten, B.; Pham, S. Ragout-a reference-assisted assembly tool for bacterial genomes. Bioinformatics 2014, 30, i302–i309. [Google Scholar] [CrossRef] [PubMed]
- Chevreux, B.; Wetter, T.; Suhai, S. Genome sequence assembly using trace signals and additional sequence information in computer science and biology. In Proceedings of the German Conference on Bioinformatics (GCB), Vienna, Austria, 26–28 September 2018. [Google Scholar]
- Jacobson, E.R.; Sundberg, J.P.; Gaskin, J.M.; Kollias, G.V.; O’Banion, M.K. Cutaneous papillomas associated with a herpesvirus-like infection in a herd of captive African elephants. J. Am. Vet. Med. Assoc. 1986, 189, 1075–1078. [Google Scholar] [PubMed]
- McCully, R.M.; Basson, P.A.; Pienaar, J.G.; Erasmus, B.J.; Young, E. Herpes nodules in the lung of the African elephant (Loxodonta africana (Blumebach, 1792)). Onderstepoort J. Vet. Res. 1971, 38, 225–235. [Google Scholar] [PubMed]
- Richman, L.K.; Montali, R.J.; Cambre, R.C.; Schmitt, D.; Hardy, D.; Hildbrandt, T.; Bengis, R.G.; Hamzeh, F.M.; Shahkolahi, A.; Hayward, G.S. Clinical and pathological findings of a newly recognized disease of elephants caused by endotheliotropic herpesviruses. J. Wildl. Dis. 2000, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Barman, N.N.; Choudhury, B.; Kumar, V.; Koul, M.; Gogoi, S.M.; Khatoon, E.; Chakroborty, A.; Basumatary, P.; Barua, B.; Rahman, T.; et al. Incidence of elephant endotheliotropic herpesvirus in Asian elephants in India. Vet. Microbiol. 2017, 208, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Seilern-Moy, K.; Darpel, K.; Steinbach, F.; Dastjerdi, A. Distribution and load of elephant endotheliotropic herpesviruses in tissues from associated fatalities of Asian elephants. Virus Res. 2016, 220, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, B.; Burkhardt, S.; Goltz, M.; Bergmann, V.; Ochs, A.; Weiler, H.; Hentschke, J. Genetic and ultrastructural characterization of a European isolate of the fatal endotheliotropic elephant herpesvirus. J. Gen. Virol. 2001, 82, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Zachariah, A.; Zong, J.C.; Long, S.Y.; Latimer, E.M.; Heaggans, S.Y.; Richman, L.K.; Hayward, G.S. Fatal herpesvirus hemorrhagic disease in wild and orphan asian elephants in southern India. J. Wildl. Dis. 2013, 49, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Kopp, A.; Blewett, E.; Misra, V.; Mettenleiter, T.C. Proteolytic cleavage of bovine herpesvirus 1 (BHV-1) glycoprotein gB is not necessary for its function in BHV-1 or pseudorabies virus. J. Virol. 1994, 68, 1667–1674. [Google Scholar] [PubMed]
- Oliver, S.L.; Sommer, M.; Zerboni, L.; Rajamani, J.; Grose, C.; Arvin, A.M. Mutagenesis of varicella-zoster virus glycoprotein B: Putative fusion loop residues are essential for viral replication, and the furin cleavage motif contributes to pathogenesis in skin tissue in vivo. J. Virol. 2009, 83, 7495–7506. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.S.; Heldwein, E.E. Herpesvirus gB: A Finely Tuned Fusion Machine. Viruses 2015, 7, 6552–6569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchers, K.; Weigelt, W.; Buhk, H.J.; Ludwig, H.; Mankertz, J. Conserved domains of glycoprotein B (gB) of the monkey virus, simian agent 8, identified by comparison with herpesvirus gBs. J. Gen. Virol. 1991, 72, 2299–2304. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, M.K.; Compton, T. Human cytomegalovirus glycoprotein B is required for virus entry and cell-to-cell spread but not for virion attachment, assembly, or egress. J. Virol. 2009, 83, 3891–3903. [Google Scholar] [CrossRef] [PubMed]
- Spear, P.G. Herpes simplex virus: Receptors and ligands for cell entry. Cell. Microbiol. 2004, 6, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.; Braun, B.; Brandmuller, C.; Kaaden, O.R.; Osterrieder, N. Analysis of the contributions of the equine herpesvirus 1 glycoprotein gB homolog to virus entry and direct cell-to-cell spread. Virology 1997, 227, 281–294. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EEHV-1_Ter_For | actgcaaaygcattcttaaaagat |
| EEHV-1_Ter_Rev | agaatgggattrgctaagaagct |
| EEHV-1_Ter_Probe | tcaacgaggagatattaggcaccaccaaca |
| EEHV-1_Ter_Oligo | cattgacactggaatctgttagaatgggattggctaagaagctcgtgttggtggtgcctaatatctcctcgttgaacgaatcttttaagaatgcgtttgcagtttttttgatattcaaattaa |
| Ele_TNFα_For | cccatctacctgggaggagtct |
| Ele_TNFα_Rev | tcgagatagtcaggcagattgatc |
| Ele_TNFα_Probe | ccagctagagaagggt |
| Ele_TNFα_Oligo | tgaggccaagccctggtatgagcccatctacctgggaggagtcttccagctagagaagggtgatcgactcagcgctgagatcaatctgcctgactatctcgactttgccgagtctgggcaggtca |
| Tissues | Normalized Viral Genome Copies | Tissues | Normalized Viral Genome Copies |
|---|---|---|---|
| Bone marrow | 3.17 × 108 | Lung | 1.19 × 107 |
| Heart | 2.25 × 108 | Temporal gland | 1.15 × 107 |
| Liver | 1.65 × 108 | Colon | 1.00 × 106 |
| Urinary bladder | 9.49 × 107 | Adrenal gland (left) | 7.00 × 105 |
| Trunk | 6.79 × 107 | Uterus | 5.62 × 105 |
| Axillar LN | 6.71 × 107 | Mesenteric LN | 5.55 × 105 |
| Tongue | 5.36 × 107 | Adrenal gland (right) | 5.25 × 105 |
| Muscle | 5.11 × 107 | Thymus | 3.81 × 105 |
| PBMC | 4.04 × 107 | Thyroid | 3.46 × 105 |
| Cervical LN | 3.98 × 107 | Cerebrum | 3.44 × 105 |
| Tonsils | 3.04 × 107 | Small intestine | 3.17 × 105 |
| Mammary gland | 2.97 × 107 | Kidney | 2.68 × 105 |
| Stomach | 2.89 × 107 | Pancreas | 2.37 × 105 |
| Trunk mucosa | 2.88 × 107 | Spinal cord | 2.50 × 105 |
| Spleen | 2.87 × 107 | Salivary gland | 1.53 × 105 |
| Mandibular LN | 2.34 × 107 | Prescapular LN | 8.72 × 104 |
| Aorta | 1.69 × 107 | Cerebellum | 7.79 × 104 |
| Blood vessel | 1.46 × 107 | Gall bladder | 7.05 × 104 |
| Blood | 1.37 × 107 | Inguinal LN | 5.25 × 103 |
| Tissue | Cell Line | Pass 1 | Pass 2 | Pass 3 | Pass 4 | Pass 5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cell | Sup | Cell | Sup | Cell | Sup | Cell | Sup | Cell | Sup | ||
| Tongue “Kanja” | ENL-2 | 12775 | 136614 | 2018 | 5128 | - | 212 | - | - | - | - |
| PBMC “Kanja” | ENL-2 | 1944 | 4233 | 110 | 301 | - | - | - | 56 | - | - |
| PBMC “Anjuli” | ENL-2 | 36011 | 27753 | - | 604 | 91 | 1279 | - | 538 | - | - |
| PBMC | 19873 | 15322 | 976 | 392 | - | - | - | - | - | - | |
| Tissue | Cell Line | Pass 1 | Pass 2 | Pass 3 | Pass 4 | Pass 5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cell | Sup | Cell | Sup | Cell | Sup | Cell | Sup | Cell | Sup | ||
| Tongue “Kanja” | CrFK | 30 | 25 | 37 | 32 | - | - | - | - | - | - |
| MDCK II | 32 | 25 | - | 37 | - | - | - | - | - | - | |
| Vero | 29 | 23 | - | 34 | - | - | - | - | - | - | |
| BD | 32 | 28 | - | 32 | - | - | - | - | - | - | |
| RK-13 | 31 | 25 | - | 37 | - | - | - | - | - | - | |
| 293T | 31 | 26 | 37 | 33 | - | 38 | - | - | - | - | |
| ED | 30 | 24 | - | 32 | - | - | - | - | - | - | |
| EC | 29 | 24 | 36 | 31 | - | 37 | - | - | - | - | |
| PBMC “Kanja” | CrFK | 33 | 31 | - | 37 | - | - | - | - | - | - |
| MDCK II | 34 | 31 | - | 38 | - | - | - | - | - | - | |
| Vero | 31 | 31 | - | - | - | - | - | - | - | - | |
| BD | 35 | 36 | - | 37 | - | - | - | - | - | - | |
| RK-13 | 34 | 31 | 37 | - | - | - | - | - | - | - | |
| 293T | 34 | 32 | - | - | - | - | - | - | - | - | |
| ED | 36 | 34 | 37 | 36 | - | - | - | - | - | - | |
| EC | 32 | 31 | 35 | 35 | - | 37 | - | - | - | - | |
| Blood “Kanja” | CrFK | 38 | 29 | - | - | - | - | - | - | - | - |
| MDCK II | - | 30 | - | - | - | - | - | - | - | - | |
| Vero | 38 | 29 | - | - | - | - | - | - | - | - | |
| BD | 38 | 35 | - | 38 | - | - | - | - | - | - | |
| RK-13 | - | 30 | - | - | - | - | - | - | - | - | |
| 293T | 38 | 30 | - | 38 | - | - | - | - | - | - | |
| ED | - | 29 | - | - | - | - | - | - | - | - | |
| EC | 33 | 29 | - | 35 | - | - | - | - | - | - | |
| Spleen “Kanja” | CRFK | 32 | 25 | - | 33 | - | - | - | - | - | - |
| MDCK II | 34 | 26 | - | 35 | - | - | - | - | - | - | |
| Vero | 33 | 26 | - | 36 | - | - | - | - | - | - | |
| BD | 33 | 28 | - | 33 | - | - | - | - | - | - | |
| RK-13 | 31 | 25 | - | 36 | - | - | - | - | - | - | |
| 293T | 32 | 26 | - | 34 | - | - | - | - | - | - | |
| NBL-6 | 34 | 25 | - | 34 | - | - | - | - | - | - | |
| EC | 37 | 25 | - | 36 | - | - | - | - | - | - | |
| PBMC “Anjuli” | HrT-18G | 29 | 29 | 38 | 38 | - | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavulraj, S.; Eschke, K.; Prahl, A.; Flügger, M.; Trimpert, J.; van den Doel, P.B.; Andreotti, S.; Kaessmeyer, S.; Osterrieder, N.; Azab, W. Fatal Elephant Endotheliotropic Herpesvirus Infection of Two Young Asian Elephants. Microorganisms 2019, 7, 396. https://doi.org/10.3390/microorganisms7100396
Pavulraj S, Eschke K, Prahl A, Flügger M, Trimpert J, van den Doel PB, Andreotti S, Kaessmeyer S, Osterrieder N, Azab W. Fatal Elephant Endotheliotropic Herpesvirus Infection of Two Young Asian Elephants. Microorganisms. 2019; 7(10):396. https://doi.org/10.3390/microorganisms7100396
Chicago/Turabian StylePavulraj, Selvaraj, Kathrin Eschke, Adriane Prahl, Michael Flügger, Jakob Trimpert, Petra B. van den Doel, Sandro Andreotti, Sabine Kaessmeyer, Nikolaus Osterrieder, and Walid Azab. 2019. "Fatal Elephant Endotheliotropic Herpesvirus Infection of Two Young Asian Elephants" Microorganisms 7, no. 10: 396. https://doi.org/10.3390/microorganisms7100396