Conferring the Metabolic Self-Sufficiency of the CAM Plasmid of Pseudomonas putida ATCC 17453: The Key Role of Putidaredoxin Reductase


Abstract
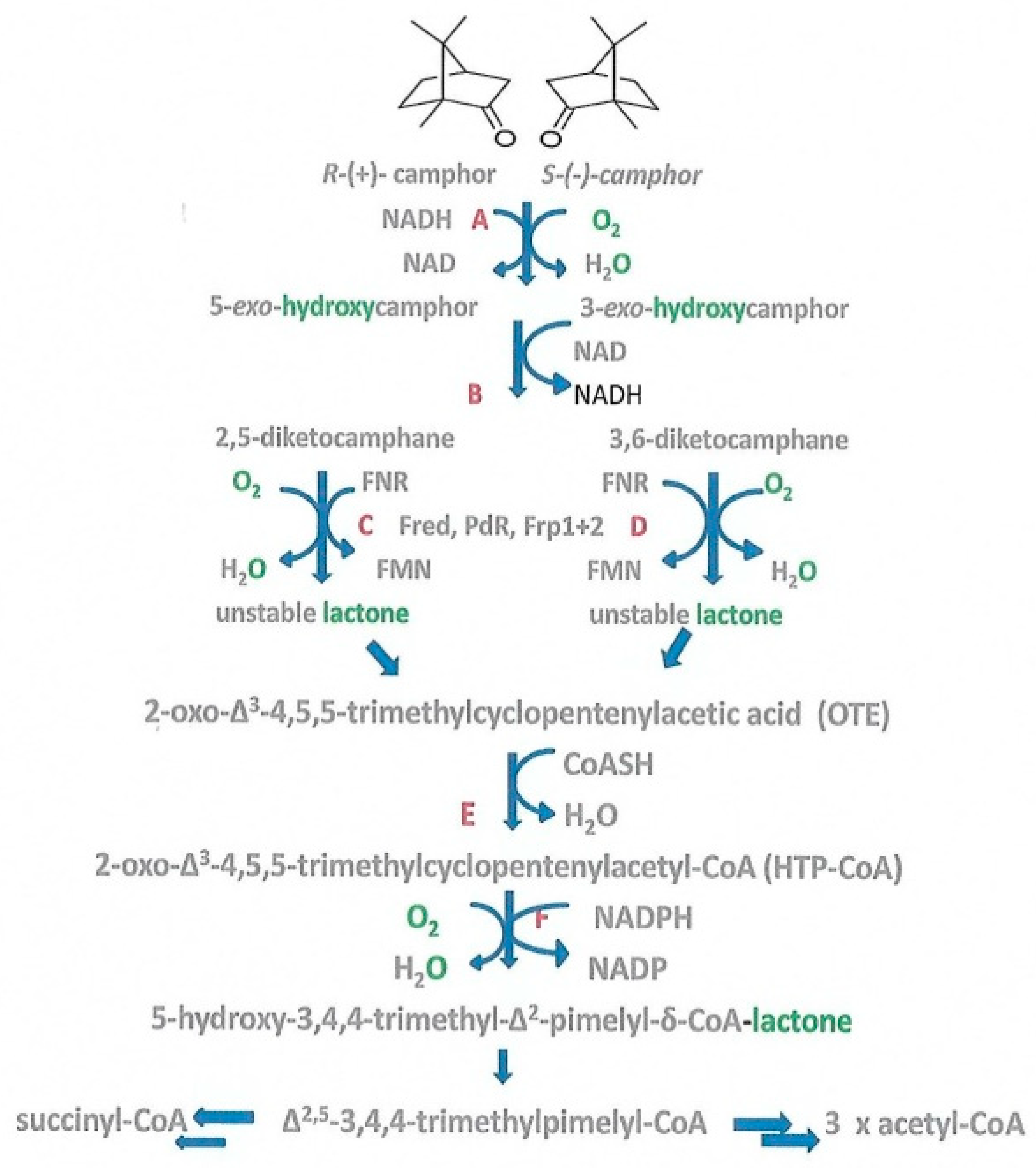
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Maintenance and Growth Conditions
2.2. Culture Samples and Extract Preparation to Determine the Effects of Zn2+ Addition to the Diauxic Growth Medium
2.3. Purification of FMN-Reductase Activities
2.4. Purification of PdR
2.5. Assays of Enzyme Activities in the Crude Cell-Free Extracts and the Semi-Purified High and Low MW Ultrafiltrates
2.6. Kinetic Studies of Highly Purified Frp1, Frp2, Fred and PdR
2.7. Reproducibility
2.8. Chemicals and General Procedures
3. Results and Discussion
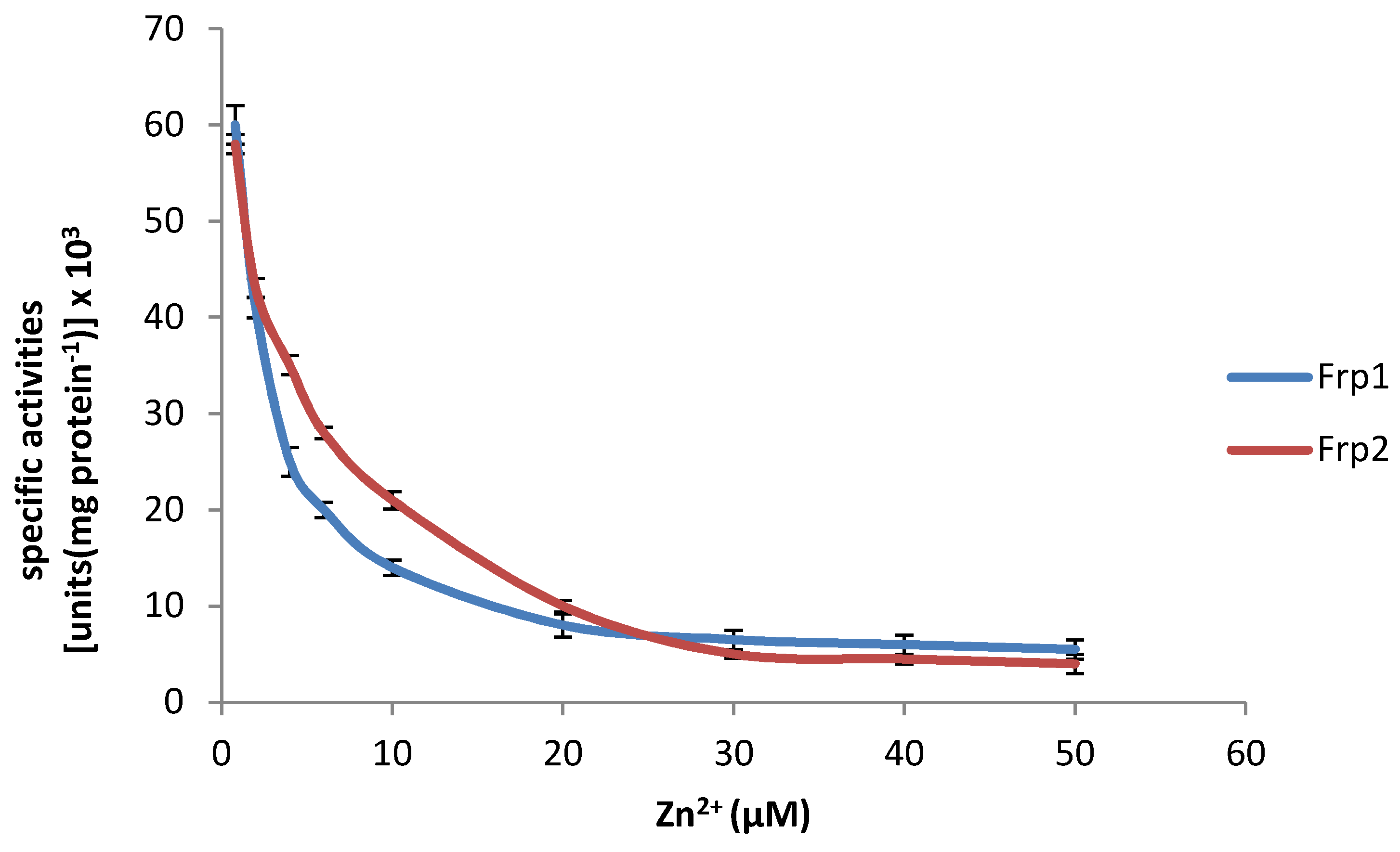
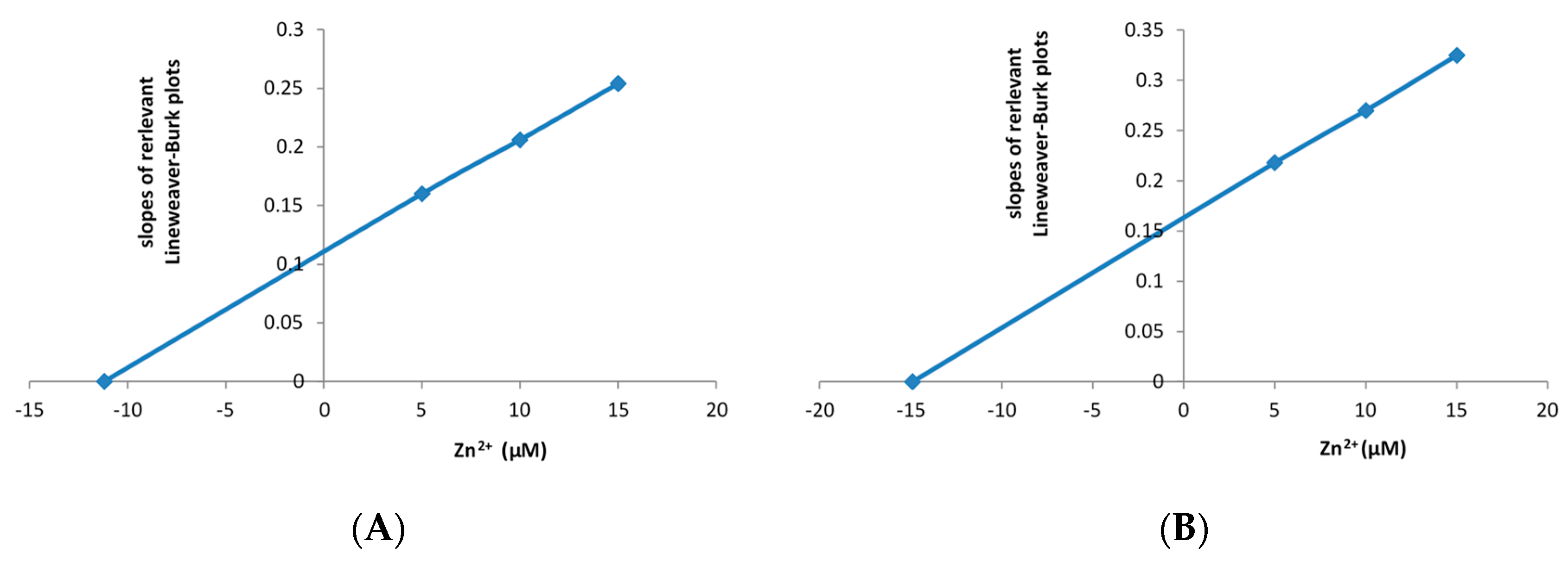
3.1. Effect of Zn2+ on Purified Preparations of the Frp1 and Frp2 of P. putida ATCC 17453
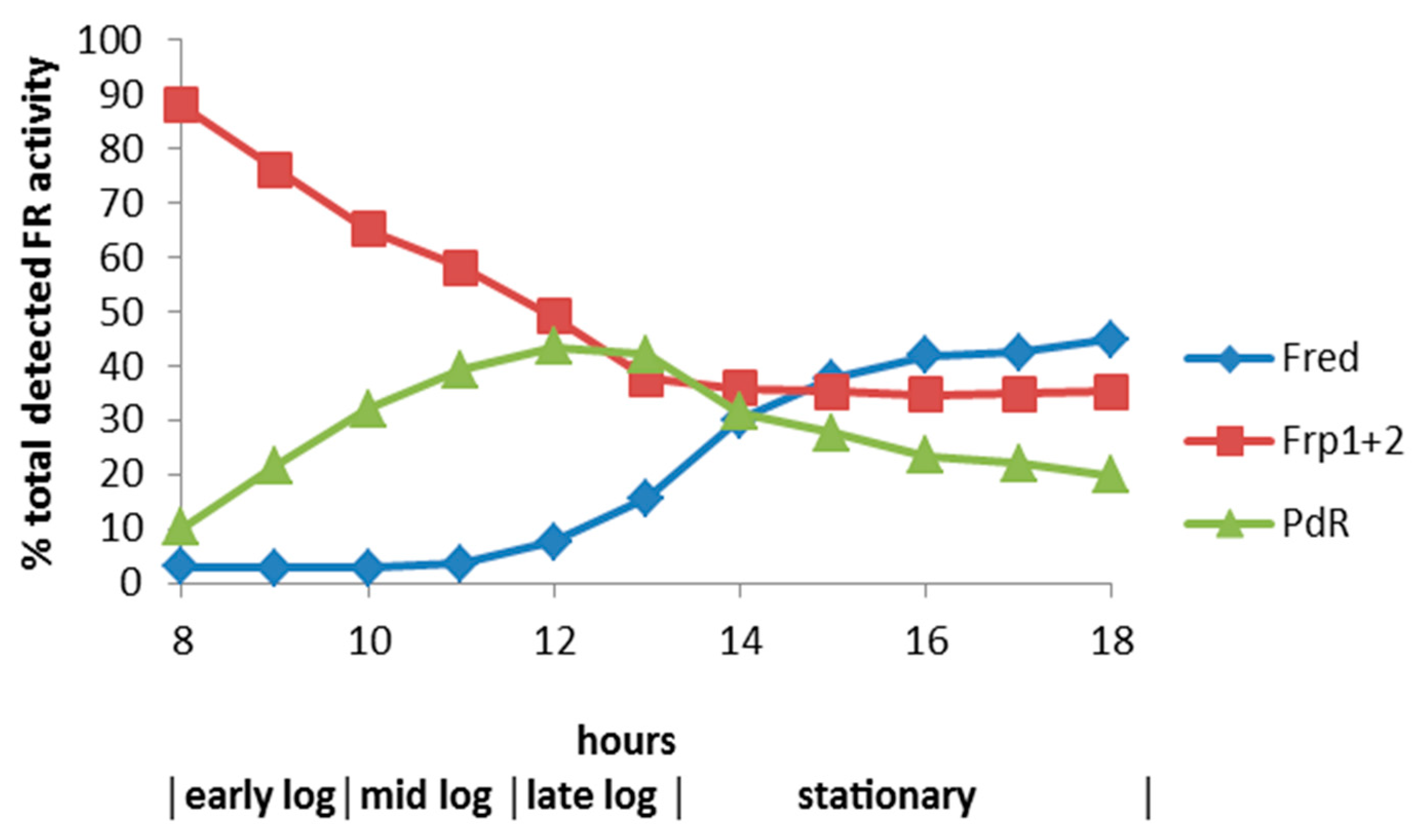
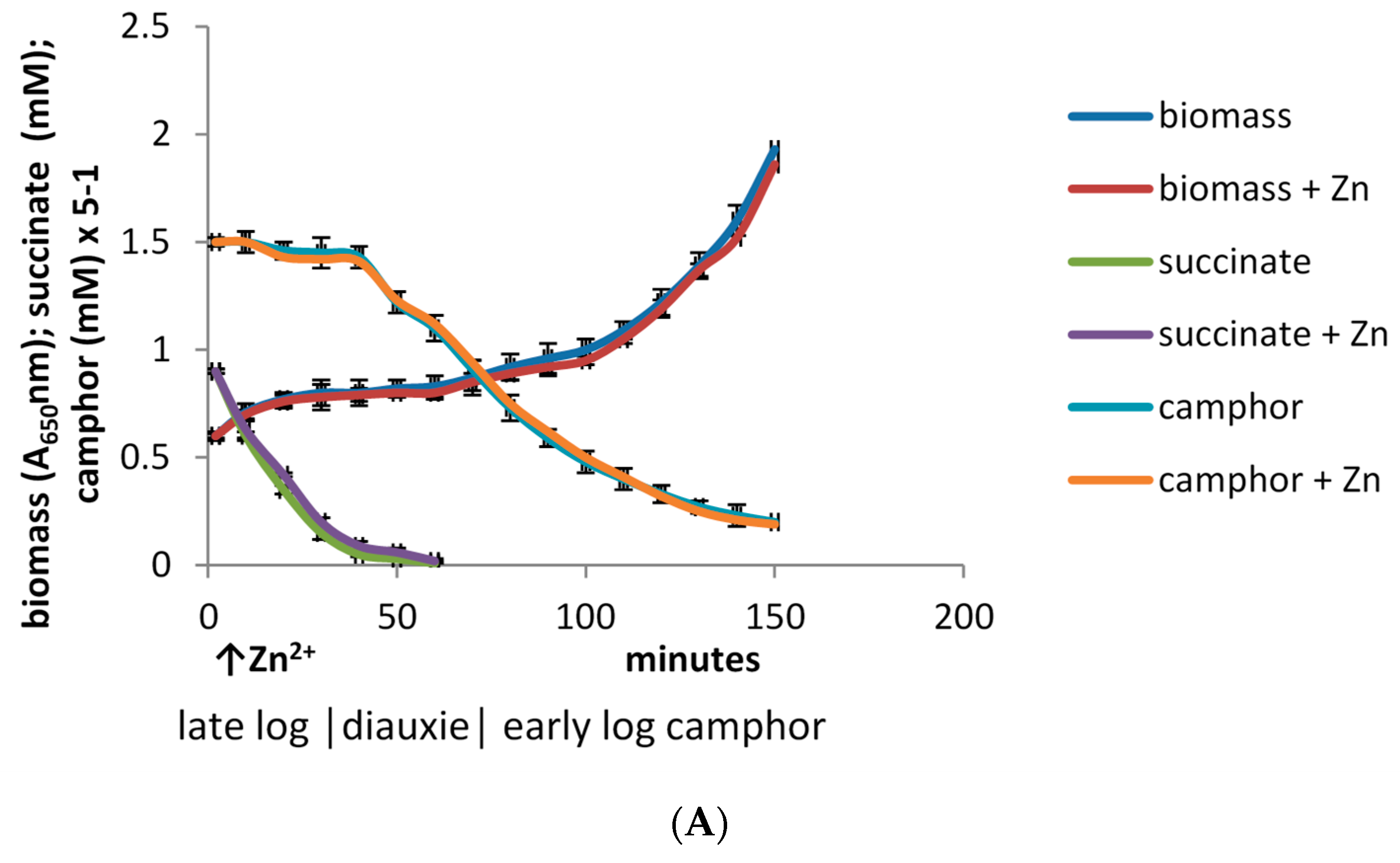
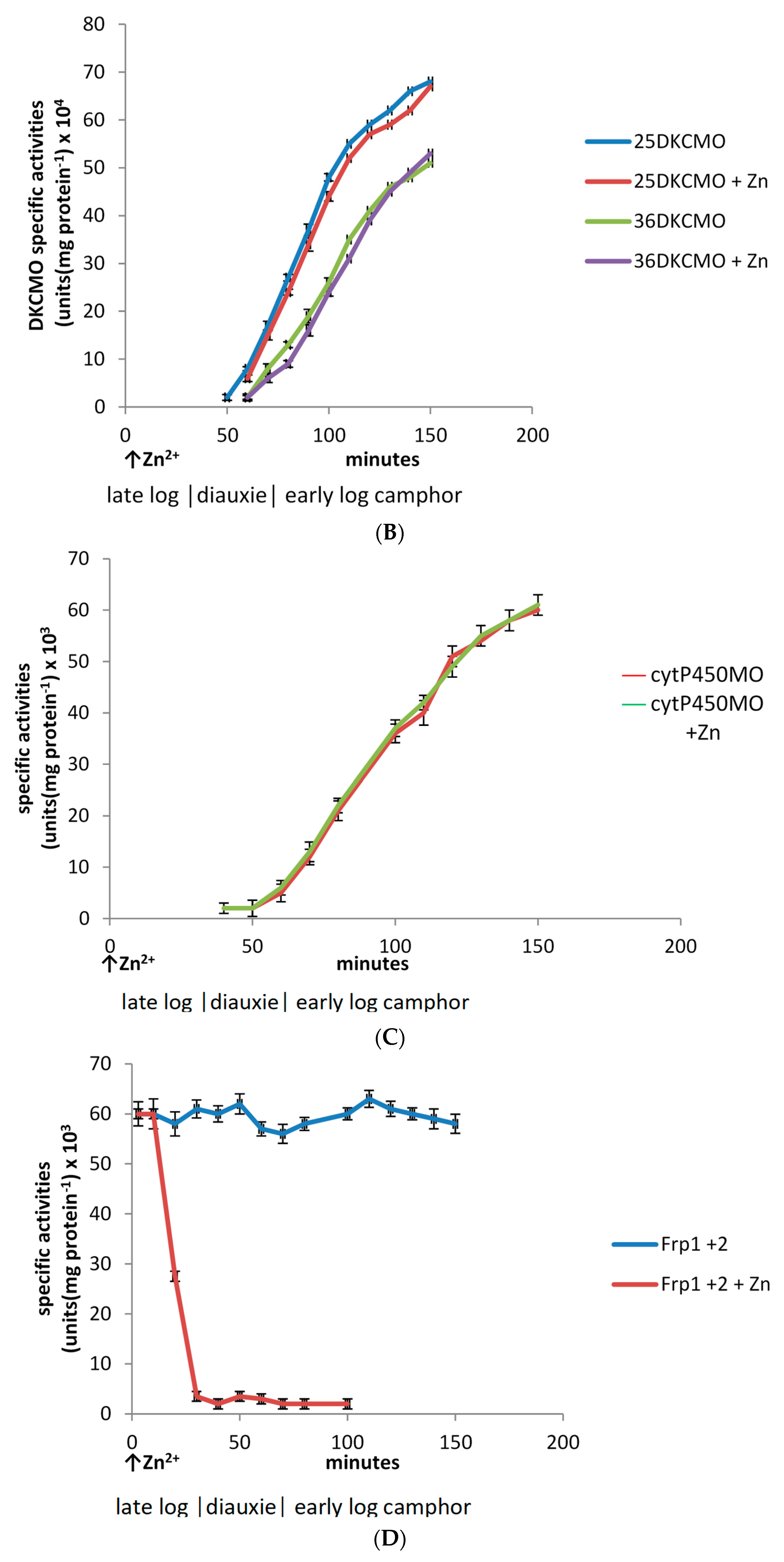
3.2. Effect of Zn2+ on Culture Density and the Specific Activities of Key Enzymes throughout Growth of P. putida ATCC 17453 on (rac)-Camphor
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rheinwald, J.G.; Chakrabarty, A.M.; Gunsalus, I.C. A transmissible plasmid controlling camphor oxidation in Pseudomonas putida. Proc. Natl. Acad. Sci. USA 1973, 70, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, J.A.; Owen, D.J.; Kok, M.; Eggink, G. Pseudomonas hydrocarbon oxidation. In Genetic control of environmental pollution; Omenn, G.S., Hollnender, A., Eds.; Plenum: New York, NY, USA, 1984; pp. 229–238. [Google Scholar]
- Chakrabarty, A.M. Genetic basis of the biodegradation of salicylate in Pseudomonas. J. Bacteriol. 1972, 112, 815–823. [Google Scholar] [PubMed]
- Yen, K.M.; Gunsalus, I.C. Plasmid gene organization: Naphthalene/salicylate oxidation. Proc. Natl. Acad. Sci. USA 1982, 79, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Duggleby, C.J.; Bayley, S.A.; Worsey, M.J.; Williams, P.S.; Broda, P. Molecular sizes and relationships of TOL plasmids in Pseudfomonas. J. Bacteriol. 1977, 130, 1274–1280. [Google Scholar] [PubMed]
- Friello, D.A.; Mylroie, J.R.; Gibson, D.T.; Rogers, J.E.; Chakrabarty, A.M. XYL, a conjugative xylene degradative plasmid in Pseudomonas Pxy. J. Bacteriol. 1976, 127, 1217–1224. [Google Scholar] [PubMed]
- Weinberger, M.; Kolenbrander, P.E. Plasmid-determined 2-hydroxyproline utilization in Arthrobacter crystallopoietes. Can. J. Microbiol. 1979, 25, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Thacker, R.; Gunsalus, I.C. Dissociation of the NIC plasmid aggregate in Pseudomonas putida. J. Bacteriol. 1979, 137, 697–699. [Google Scholar] [PubMed]
- Fisher, P.R.; Appleton, J.; Pemberton, J.M. Isolation and characterization of a-degradative plasmid from Alcaligenes paradoxus. J. Bacteriol. 1978, 135, 798–804. [Google Scholar]
- Don, R.H.; Pemberton, J.M. Properties of six pesticide degrading plasmids isolated from Alcaligenes paradoxus and Alcaligenes eutrophus. J. Bacteriol. 1981, 145, 684–686. [Google Scholar]
- Chakrabarty, A.M.; Friello, D.A.; Bopp, L.H. Transposition of DNA segments specifying hydrocarbon degradation and their expression in various microorganisms. Proc. Natl. Acad. Sci. USA 1978, 75, 3109–3112. [Google Scholar] [CrossRef]
- Chatterjee, D.K.; Chakrabarty, A.M. Genetic homology between independently isolated chlorobenzoate-degradative plasmids. J. Bacteriol. 1983, 153, 532–534. [Google Scholar] [PubMed]
- Rheinwald, J.G. Extrachromosomal transfer between strains of Pseudomonas putida. M.Sc. Thesis, University of Illinois, Urbana, IL, USA, December 1970. [Google Scholar]
- Massey, L.K.; Sokatch, J.R.; Conrad, R.S. Branched-chain amino acid catabolism in bacteria. Bacteriol. Rev. 1976, 40, 42–54. [Google Scholar]
- Shaham, M.; Chakrabarty, A.M.; Gunsalus, I.C. Camphor plasmid-mediated chromosomal transfer in Pseudomonas putida. J. Bacteriol. 1973, 116, 944–949. [Google Scholar] [PubMed]
- Iwaki, H.; Grosse, S.; Bergeron, H.; Leisch, H.; Morley, K.; Hasegawa, Y.; Lau, P.C. Camphor pathway redox: Functional recombinant expression of 2,5- and 3,6-diketocamphane monooxygenases in Pseudomonas putida ATCC 17453 with their cognate flavin reductase catalysing Baeyer-Villiger reactions. Appl. Environ. Microbiol. 2013, 79, 3282–3293. [Google Scholar] [CrossRef] [PubMed]
- Ellis, H.R. The FMN-dependent two-component monooxygenase systems. Arch. Biochem. Biophys. 2010, 497, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Isupov, M.N.; Schroder, E.; Gibson, R.P.; Beecher, J.; Donadio, G.; Saneei, V.; Dounha, S.A.; McGhie, E.J.; Sayer, C.; Davenport, C.F.; et al. The oxygenating constituent of 3,6-diketocamphane monooxygenase from the CAM plasmid of Pseudomonas putida: The first crystal structure of a type II Baeyer-Villiger monooxygenase. Acta Crystalog. SectD 2015, 71, 2344–2351. [Google Scholar] [CrossRef] [PubMed]
- Isupov, M.N.; Schroder, E.; Gibson, R.P.; Beecher, J.; Donadio, G.; Saneei, V.; Dounha, S.A.; McGhie, E.J.; Sayer, C.; Davenport, C.F.; et al. The oxygenating constituent of 3,6-diketocamphane monooxygenase from the CAM plasmid of Pseudomonas putida: The first crystal structure of a type II Baeyer-Villiger monooxygenase. Corrigendum: Statement withdrawn. Acta Crystalog. SectD 2018, 74, 379. [Google Scholar] [CrossRef]
- Willetts, A. Characterised Flavin-Dependent Two-Component Monooxygenases from the CAM Plasmid of Pseudomonas putida ATCC 17453 (NCIMB 10007): ketolactonases by Another Name. Microorganisms 2019, 7, 1. [Google Scholar] [CrossRef]
- Willetts, A.; Kelly, D.R. Multiple native flavin reductases in camphor-metabolising Pseudomonas putida NCIMB 10007: Functional interaction with two-component diketocamphane monooxygenase isoenzymes. Microbiology 2014, 160, 1784–1794. [Google Scholar] [CrossRef] [PubMed]
- Willetts, A.; Kelly, D.R. Flavin-dependent redox transfers by two-component diketocamphane monooxygenases of camphor-grown Pseudomonas putida NCIMB. Microorganisms 2016, 4, 38. [Google Scholar] [CrossRef]
- Vising, L.C. Roles of secondary metabolism in microbes. Ciba Found. Symp. 1992, 171, 184–194. [Google Scholar]
- Gunsalus, I.C.; Wagner, G.C. Bacterial P-450cam methylene monooxygenase components: Cytochrome m, putidaredoxin and putidaredoxin reductase. Methods Enzymol. 1978, 52, 166–188. [Google Scholar]
- Lammerhofer, M.; Richter, M.; Wu, J.; Noguera, R.; Lindner, W. Mixed-mode ion-exchangers and their comparative chromoatographic characterization in reverse phase and hydrophilic interaction chromatography elution modes. J. Sep. Sci. 2008, 31, 2572–2588. [Google Scholar] [CrossRef] [PubMed]
- Gagnon, R.; Grogan, G.; Roberts, S.M.; Villa, R.; Willetts, A. Enzymatic Baeyer-Villiger oxidation of some bicyclo[2.2.1]heptan-2-ones using monooxygenases from Pseudomonas putida NCIMB 10007: Enantioselective preparation of a precursor of azadirachtin. J. Chem. Soc. Perkin Trans. 1 1995, 1505–1511. [Google Scholar] [CrossRef]
- Halle, E.; Meyer, J.M. Iron release from ferrisiderophores. A multi-step mechanisms involving a NADH/FMN oxidoreductase and a chemical reduction by FMNH2. Eur. J. Biochem. 1992, 209, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.; Oliveira, D.; Hu, G.; Kronstad, J. Role of ferric reductases in iron acquisition and virulence in the fungal pathogen Cryptococcus neoforams. Infect. Immun. 2014, 82, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Baghdianz, A. Role du zinc sur l’apparition de la composante du ‘pigment’ de Pseudomonas fluorescens (Flugge-Migula). Arch. Sci. 1952, 5, 47–48. [Google Scholar]
- Chakrabarty, A.M.; Roy, S.C. Effect of trace elements on the production of pigments by a pseudomonad. Biochem. J. 1964, 93, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Huyer, M.; Page, W.J. Zn2+ increases siderophore production in Azotobacter vinelandii. Appl. Environ. Microbiol. 1988, 54, 2625–2631. [Google Scholar]
- Huyer, M.; Page, W.J. Ferric reductase activity in Azotobacter vinelandii and its inhibition by Zn2+. J. Bacteriol. 1989, 171, 4031–4037. [Google Scholar] [CrossRef]
- Segel, I. Enzyme Kinetics; Wiley: New York, NY, USA, 1975. [Google Scholar]
- Gunsalus, I.C.; Bertland, A.U.; Jacobson, L.A. Enzyme induction and repression in anabolic and catabolic pathways. Arch. Mikrobiol. 1967, 59, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, L.A. Enzyme induction and repression in the catabolism of (+)-camphor by Pseudomonas putida. Ph.D. Thesis, University of Illinois, Urbana, IL, USA, December 1967. [Google Scholar]
- Hartline, R.A.; Gunsalus, I.C. Induction specificity and catabolite repression of the early enzymes in camphor degradation by Pseudomonas putida. J. Bacteriol. 1971, 106, 468–478. [Google Scholar] [PubMed]
- Devescovi, G.; Aguilar, C.; Majolini, M.B.; Marugg, J.; Weisbeek, P.; Venturi, V. A siderophore peptide synthetase gene from plant growth-promoting Pseudomonas putida WCSSystem. Appl. Microbiol. 2001, 24, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Lau, C.K.Y.; Krewulak, K.D.; Vogel, H.J. Bacterial ferrous iron transport: The Feo system. FEMS Microbiol. Rev. 2015, 40, 273–298. [Google Scholar] [CrossRef] [PubMed]
- Callanan, M.; Sexton, R.; Dowling, D.N.; O’Gara, F. Regulation of the iron uptake genes in Pseudomonas fluorescens M114 by pseudobactin M1214: The phr A sigma factor gene does not mediate the sidrophore regulatory response. FEMS Microbiol. Lett. 1996, 144, 61–66. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Babich, H.; Stotzky, G. Toxicity of zinc to fungi, bacteria, and coliphages: Influence of chloride ions. Appl. Enriron. Microbiol. 1978, 36, 906–914. [Google Scholar]
- Ma, L.; Terwilliger, A.; Maresso, A.W. Iron and zinc exploitation during bacterial pathogenesis. Metallomics 2015, 7, 1541–1554. [Google Scholar] [CrossRef]
- Loeschcke, A.; Thies, S. Pseudomonas putida—A versatile host for production of natural products. Appl. Microbiol. Biotechnol. 2015, 99, 6197–6214. [Google Scholar] [CrossRef]
- Aiman, S.; Shehroz, M.; Munir, M.; Gul, S.; Shah, M.; Khan, A. Species-wide genome mining of Pseudomonas putida for potential secondary metabolites and drug-like natural products characterization. J. Proteomics Biochem. 2018, 11, 001–007. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmid | Degradative Pathway | Conjugative or Non-Conjugative | Approx. Size (Daltons × 106) | Key Reference |
|---|---|---|---|---|
| CAM | Camphor | Conjugative | 300 | Rheinwald et al. [1] |
| OCT | n-Octane | Non-conjugative | 250 | Shapiro et al. [2] |
| SAL | Salicylate | Conjugative | 42–55 (various) | Chakrabarty [3] |
| NAH | Naphthalene | Conjugative | 46 | Yen and Gunsalus [4] |
| TOL | Toluene/xylene | Conjugative | 76 | Duggleby et al. [5] |
| XYL-K | Xylene/toluene | Conjugative | 90 | Friello et al. [6] |
| 2-HP | 2-Hydroxypyridine | N.D. | 63 | Weinberger and Kolenbrander [7] |
| NIC | Nicotine/nicotinate | Conjugative | N.D. | Thacker and Gunsalus [8] |
| pOAD2 | 6-Aminohexanoic acid cyclic dimer | Non-conjugative | 29 | Fisher et al. [9] |
| pJP1 | 2,4-dichlorophenoxy acetic acid | Conjugative | 58 | Don and Pemberton [10] |
| pAC8 | Xylene/toluene | Conjugative | 76 | Chakrabarty et al. [11] |
| pAC21 | p-Chlorobiphenyl | Conjugative | 65 | Chatterjee and Chakrabarty [12] |
| pAC25 | 3-Chlorobenzoate | Conjugative | 68 | Chatterjee and Chakrabarty [12] |
| pAC27 | 3- and 4-Chloro-benzoate | Conjugative | 59 | Chatterjee and Chakrabarty [12] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Willetts, A. Conferring the Metabolic Self-Sufficiency of the CAM Plasmid of Pseudomonas putida ATCC 17453: The Key Role of Putidaredoxin Reductase. Microorganisms 2019, 7, 395. https://doi.org/10.3390/microorganisms7100395
Willetts A. Conferring the Metabolic Self-Sufficiency of the CAM Plasmid of Pseudomonas putida ATCC 17453: The Key Role of Putidaredoxin Reductase. Microorganisms. 2019; 7(10):395. https://doi.org/10.3390/microorganisms7100395
Chicago/Turabian StyleWilletts, Andrew. 2019. "Conferring the Metabolic Self-Sufficiency of the CAM Plasmid of Pseudomonas putida ATCC 17453: The Key Role of Putidaredoxin Reductase" Microorganisms 7, no. 10: 395. https://doi.org/10.3390/microorganisms7100395
APA StyleWilletts, A. (2019). Conferring the Metabolic Self-Sufficiency of the CAM Plasmid of Pseudomonas putida ATCC 17453: The Key Role of Putidaredoxin Reductase. Microorganisms, 7(10), 395. https://doi.org/10.3390/microorganisms7100395