A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment

 , ,
, ,  ,
,  and
and 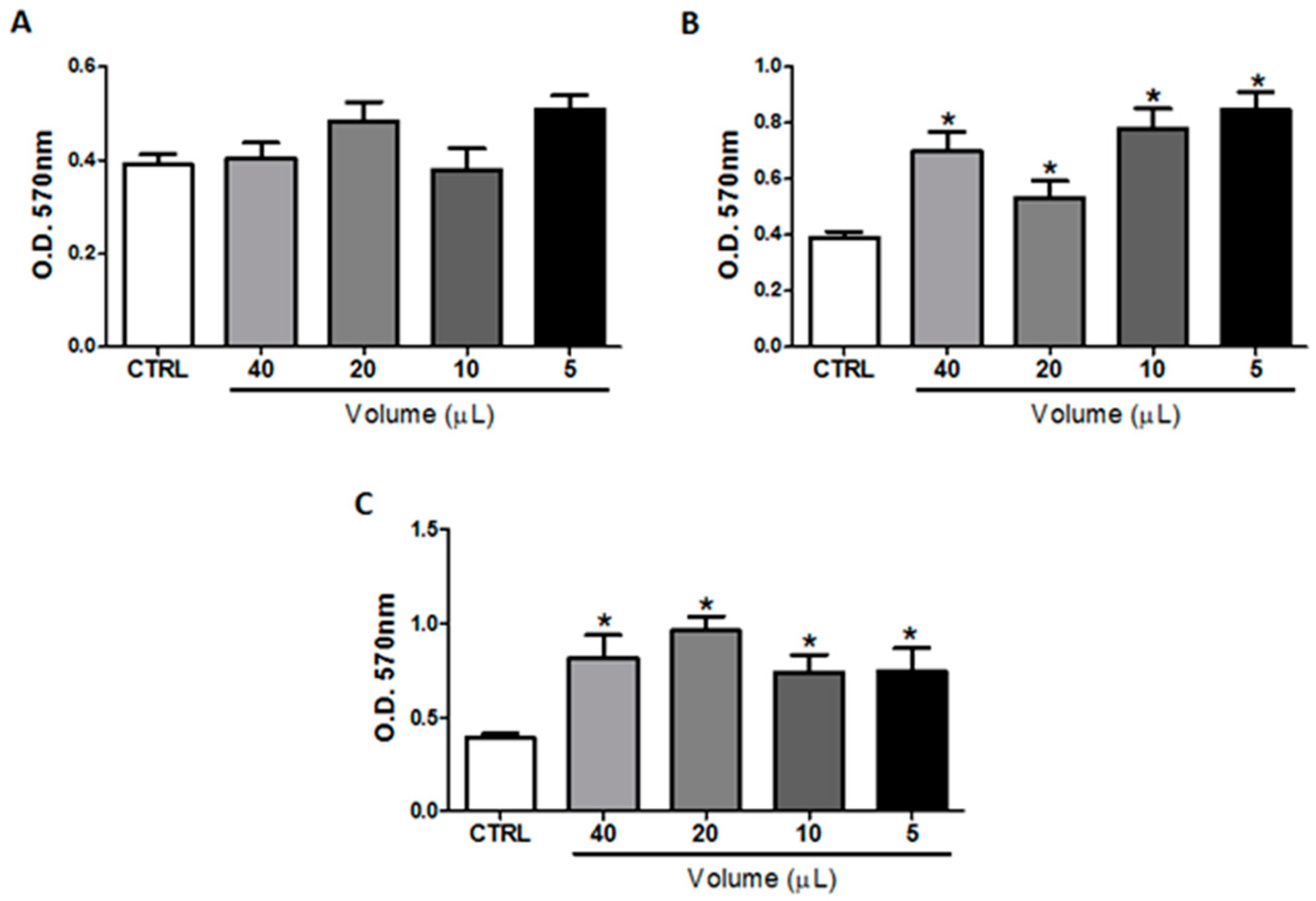{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. In Vitro Treatment with Omega-3 or Omega-6 Did Not Impair B16F10 Cells Viability
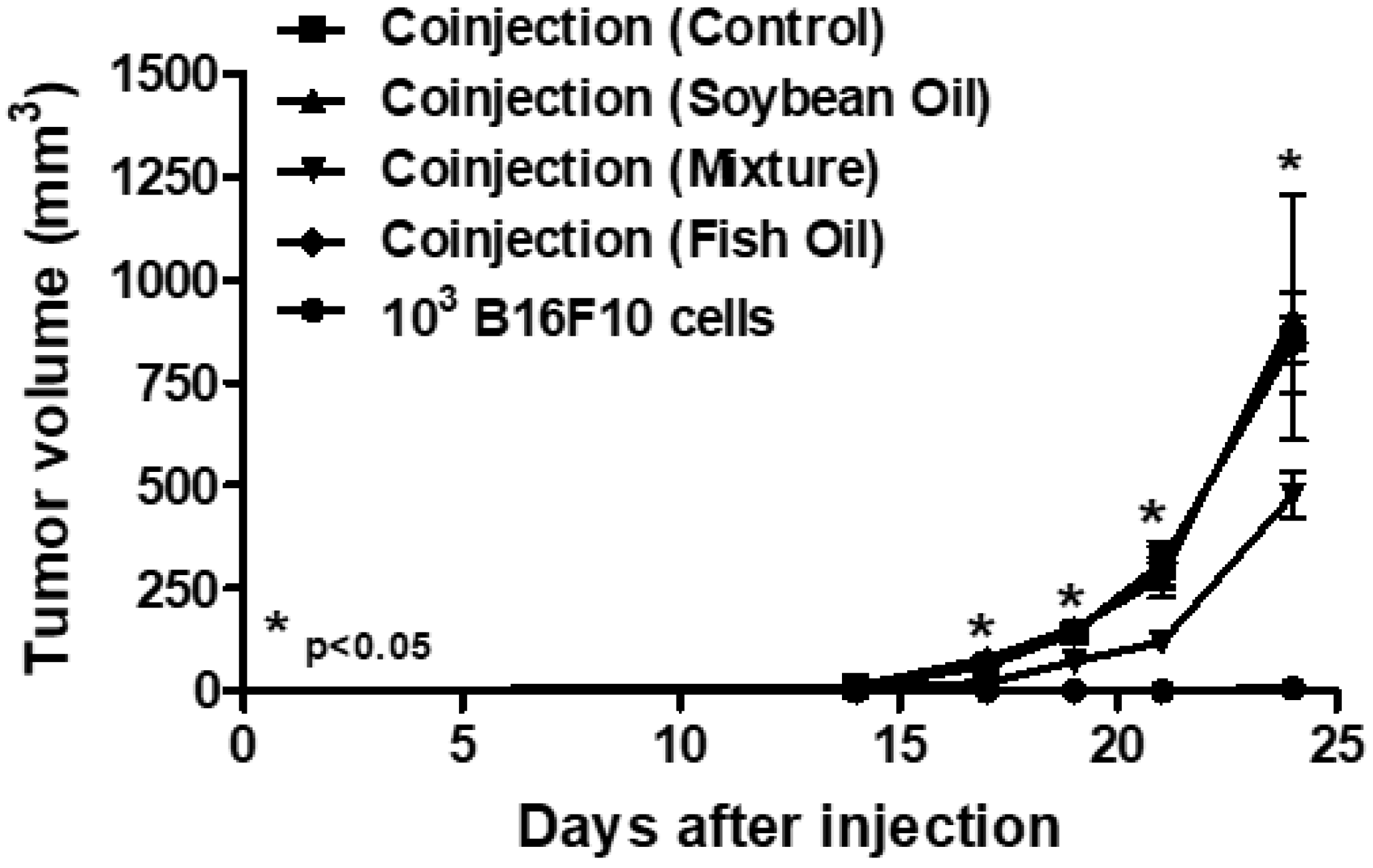
2.2. In Vivo Treatment with a Mixture of Omega-3 Andomega-6 Reduced Tumor Growth
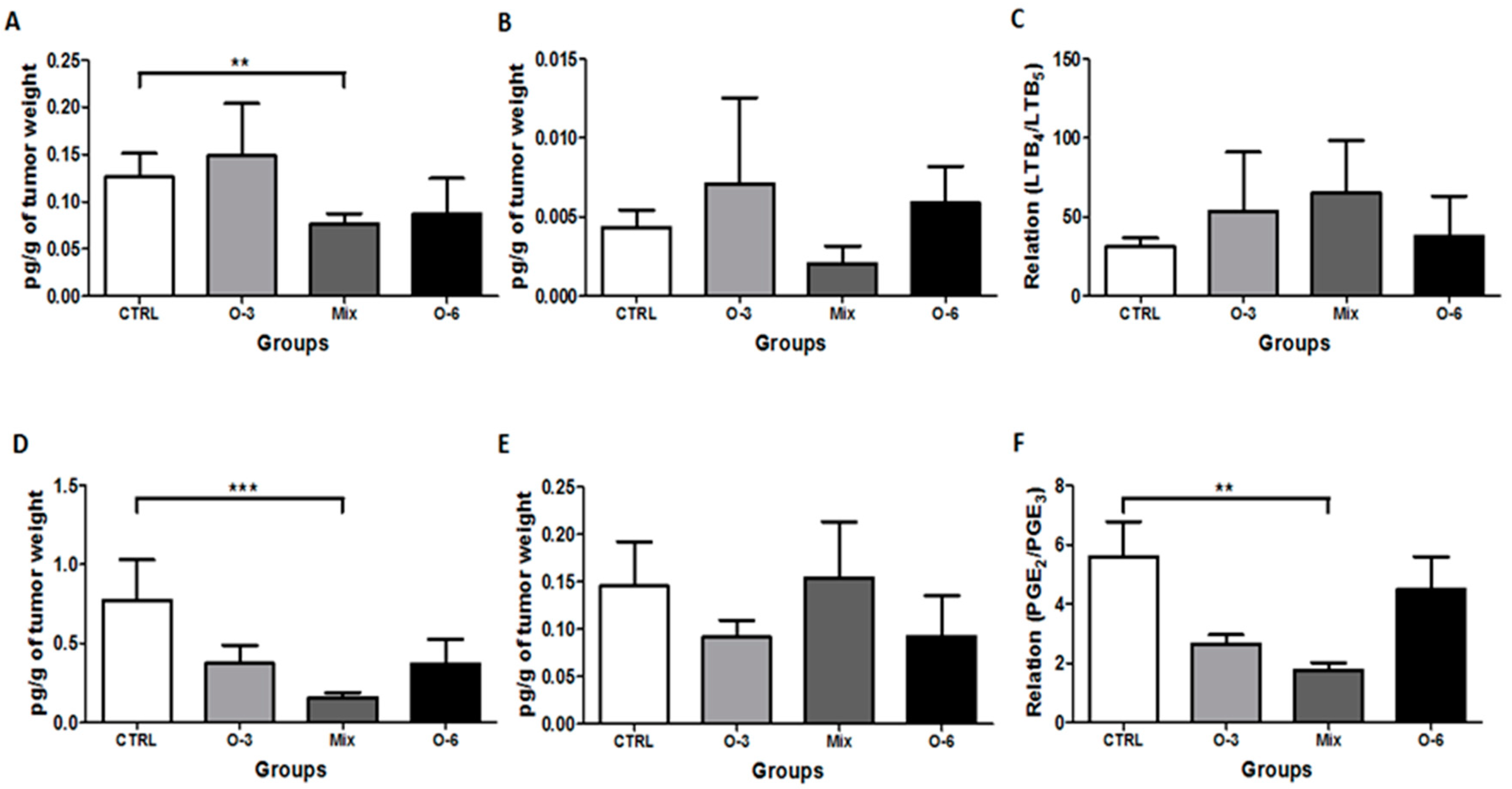
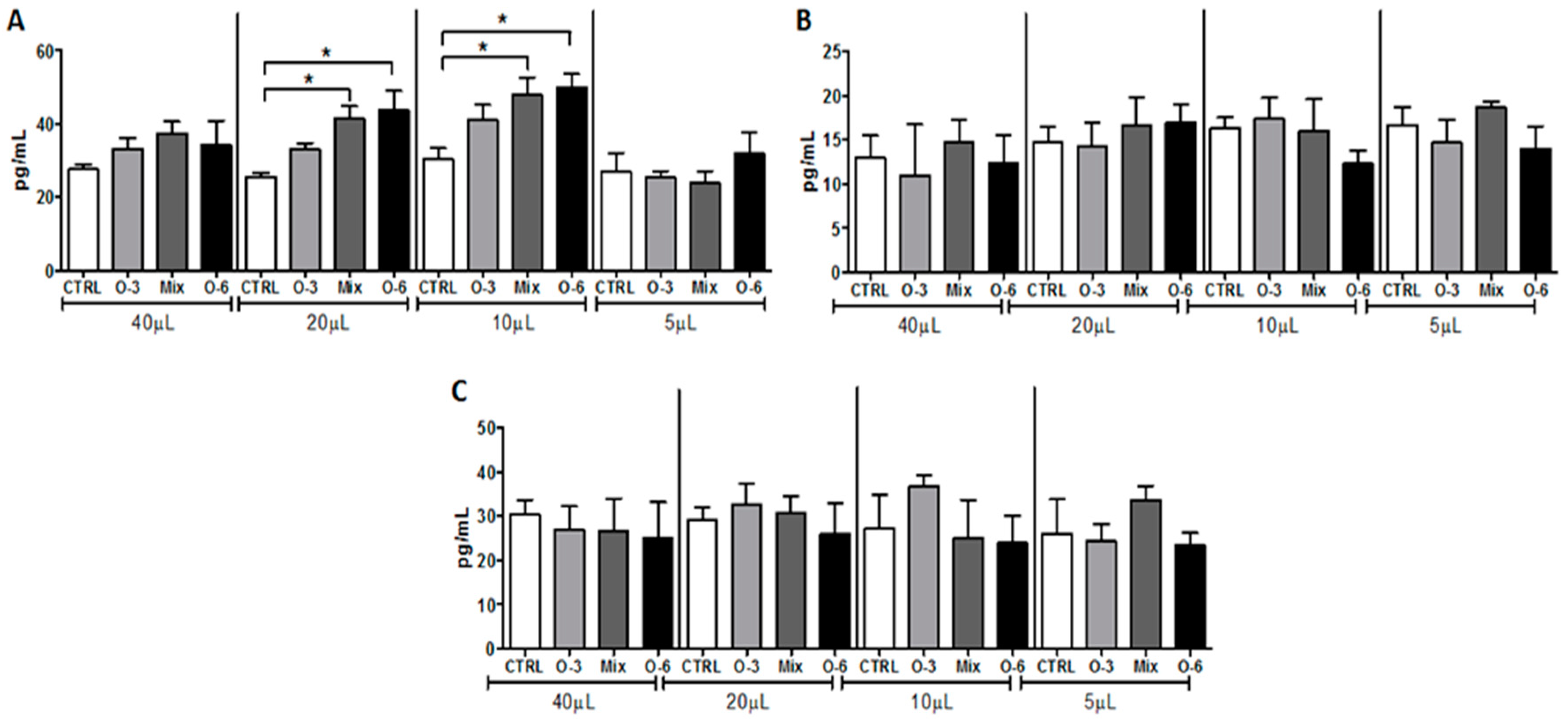
2.3. Treatment with a Mixture of Fish and Soybean Oils Reduced the Pro-Inflammatory Mediators in the Tumor Microenvironment
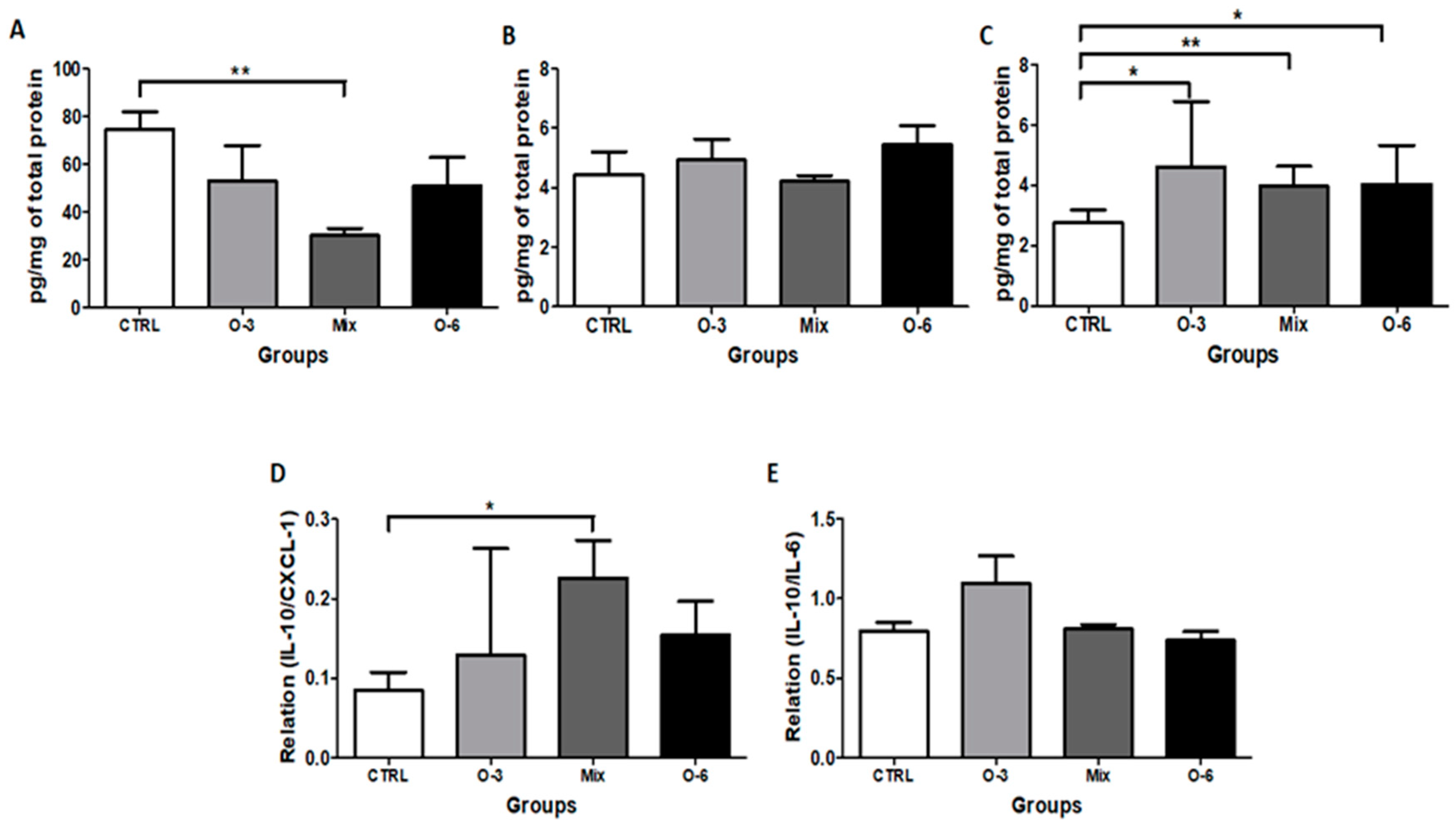
2.4. The Treatment with a Mixture of Fish and Soybean Oils Reduced the CXCL1 Levels in the Tumor Microenvironment
2.5. In Vitro Treatment with Soybean Oil Associated or not with Fish Oil, but not with Fish Oil Alone, Induced IL-10 Release from B16F10 Cells
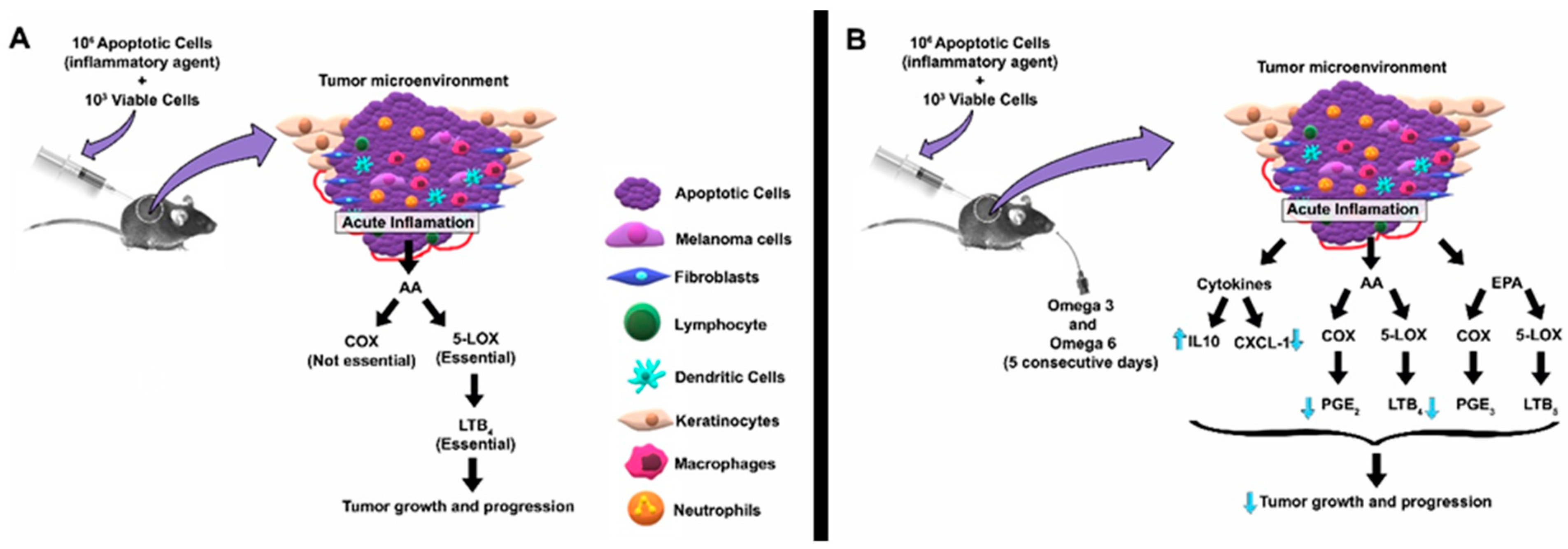
3. Discussion
4. Materials and Methods
4.1. Mice, Reagents, Cell Culture, and Cell Viability Assay
4.2. Animals
4.3. Evaluation of the Effect of Omega-3 and Omega-6 on Tumor Progression
4.4. Tumor Homogenate
4.5. Determination of Cytokines and Inflammatory Mediators in the Tumor Homogenate
4.6. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef]
- Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Bachi, A.L.L.; Kim, F.J.K.; Nonogaki, S.; Carneiro, C.R.W.; Lopes, J.D.; Jasiulionis, M.G.; Correa, M. Leukotriene B4 Creates a Favorable Microenvironment for Murine Melanoma Growth. Mol. Cancer Res. 2009, 7, 1417. [Google Scholar] [CrossRef] [PubMed]
- Nie, D.; Honn, K.V. Cyclooxygenase, lipoxygenase and tumor angiogenesis. Cell Mol. Life Sci. 2002, 59, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N. Endogenous pro-resolving and anti-inflammatory lipid mediators: A new pharmacologic genus. Br. J. Pharmacol. 2008, 153, S200–S215. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Pierotti, M.A. Cancer and inflammation: A complex relationship. Cancer Lett. 2008, 267, 180–181. [Google Scholar] [CrossRef]
- Culp, S.J.; Mellick, P.W.; Trotter, R.W.; Greenlees, K.J.; Kodell, R.L.; Beland, F.A. Carcinogenicity of malachite green chloride and leucomalachite green in B6C3F1 mice and F344 rats. Food Chem. Toxicol. 2006, 44, 1204–1212. [Google Scholar] [CrossRef]
- Torisu-Itakura, H.; Lee, J.H.; Scheri, R.P.; Huynh, Y.; Ye, X.; Essner, R.; Morton, D.L. Molecular characterization of inflammatory genes in sentinel and nonsentinel nodes in melanoma. Clin. Cancer Res. 2007, 13, 3125–3132. [Google Scholar] [CrossRef]
- Correa, M.; Machado, J.; Carneiro, C.R.; Pesquero, J.B.; Bader, M.; Travassos, L.R.; Chammas, R.; Jasiulionis, M.G. Transient inflammatory response induced by apoptotic cells is an important mediator of melanoma cell engraftment and growth. Int. J. Cancer 2005, 114, 356–363. [Google Scholar] [CrossRef]
- Garófolo, A.; Petrilli, A.S. Balanço entre ácidos graxos ômega-3 e 6 na resposta inflamatória em pacientes com câncer e caquexia. Rev. De Nutr. 2006, 19, 611–621. [Google Scholar] [CrossRef]
- Kang, J.X. The Omega-6/Omega-3 Fatty Acid Ratio in Chronic Diseases: Animal Models and Molecular Aspects. World Rev. Nutr. Diet. 2011, 102, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Roynette, C.E.; Calder, P.C.; Dupertuis, Y.M.; Pichard, C. n-3 polyunsaturated fatty acids and colon cancer prevention. Clin. Nutr. 2004, 23, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ma, D.W. The role of n-3 polyunsaturated fatty acids in the prevention and treatment of breast cancer. Nutrients 2014, 6, 5184–5223. [Google Scholar] [CrossRef] [PubMed]
- Curi, R.; Pompéia, C.; Miyasaka, C.; Procòpio, J. Entendendo a Gordura: Os ácidos Graxos; Manole, E., Ed.; São Paulo, Brazil, 2002. Available online: https://www.amazon.com.br/Entendendo-gordura-Os-%C3%81cidos-Graxos/dp/8520413552 (accessed on 31 July 2019).
- Soufli, I.; Toumi, R.; Rafa, H.; Touil-Boukoffa, C. Overview of cytokines and nitric oxide involvement in immuno-pathogenesis of inflammatory bowel diseases. World J. Gastrointest. Pharm. 2016, 7, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Baggiolini, M.; Dewald, B.; Moser, B. Human Chemokines: An Update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef] [PubMed]
- Asensio, V.C.; Campbell, I.L. Chemokines in the CNS: Plurifunctional mediators in diverse states. Trends Neurosci. 1999, 22, 504–512. [Google Scholar] [CrossRef]
- Kim, C. Chemokine-chemokine receptor network in immune cell trafficking. Curr. Drug Targets Immune Endocr. Metab. Disord. 2004, 4, 343–361. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Zlotnik, A.; Yoshie, O. Chemokines: A New Classification System and Their Role in Immunity. Immunity 2000, 12, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Palomino, D.C.T.; Marti, L.C. Chemokines and immunity. Einstein (São Paulo) 2015, 13, 469–473. [Google Scholar] [CrossRef]
- Guerreiro, R.; Santos-Costa, Q.; Azevedo-Pereira, J.M. The chemokines and their receptors: Characteristics and physiological functions. Acta Med. Port. 2011, 24 (Suppl. 4), 967–976. [Google Scholar] [PubMed]
- Varney, M.L.; Johansson, S.L.; Singh, R.K. Distinct expression of CXCL8 and its receptors CXCR1 and CXCR2 and their association with vessel density and aggressiveness in malignant melanoma. Am. J. Clin. Pathol. 2006, 125, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Varney, M.L.; Li, A.; Dave, B.J.; Bucana, C.D.; Johansson, S.L.; Singh, R.K. Expression of CXCR1 and CXCR2 receptors in malignant melanoma with different metastatic potential and their role in interleukin-8 (CXCL-8)-mediated modulation of metastatic phenotype. Clin. Exp. Metastasis 2003, 20, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Kodet, O.; Lacina, L.; Krejčí, E.; Dvořánková, B.; Grim, M.; Štork, J.; Kodetová, D.; Vlček, Č.; Šáchová, J.; Kolář, M.; et al. Melanoma cells influence the differentiation pattern of human epidermal keratinocytes. Mol. Cancer 2015, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Castell, J.V.; Andus, T. Interleukin-6 and the acute phase response. Biochem. J. 1990, 265, 621–636. [Google Scholar] [CrossRef] [PubMed]
- Stumpf, C.; Lehner, C.; Yilmaz, A.; Daniel, W.G.; Garlichs, C.D. Decrease of serum levels of the anti-inflammatory cytokine interleukin-10 in patients with advanced chronic heart failure. Clin. Sci. (Lond.) 2003, 105, 45–50. [Google Scholar] [CrossRef]
- Voltarelli, F.A.; Frajacomo, F.T.; Padilha, C.S.; Testa, M.T.J.; Cella, P.S.; Ribeiro, D.F.; de Oliveira, D.X.; Veronez, L.C.; Bisson, G.S.; Moura, F.A.; et al. Syngeneic B16F10 Melanoma Causes Cachexia and Impaired Skeletal Muscle Strength and Locomotor Activity in Mice. Front. Physiol. 2017, 8, 715. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.-C.; Puaux, A.-L.; Chittezhath, M.; Shalova, I.; Kajiji, T.S.; Wang, X.; Abastado, J.-P.; Lam, K.-P.; Biswas, S.K. Macrophage polarization to a unique phenotype driven by B cells. Eur. J. Immunol. 2010, 40, 2296–2307. [Google Scholar] [CrossRef] [PubMed]
- Lopes, J.D.; Mariano, M. B-1 cell: The precursor of a novel mononuclear phagocyte with immuno-regulatory properties. An. Da. Acad. Bras. De Ciências 2009, 81, 489–496. [Google Scholar] [CrossRef]
- Rehman, K.; Mohd Amin, M.C.; Yuen, N.P.; Zulfakar, M.H. Immunomodulatory Effectiveness of Fish Oil and omega-3 Fatty Acids in Human Non-melanoma Skin Carcinoma Cells. J. Oleo Sci. 2016, 65, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Sulciner, M.L.; Serhan, C.N.; Gilligan, M.M.; Mudge, D.K.; Chang, J.; Gartung, A.; Lehner, K.A.; Bielenberg, D.R.; Schmidt, B.; Dalli, J.; et al. Resolvins suppress tumor growth and enhance cancer therapy. J. Exp. Med. 2018, 215, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Elias, E.G.; Hasskamp, J.H.; Sharma, B.K. Cytokines and growth factors expressed by human cutaneous melanoma. Cancers (Basel) 2010, 2, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Tuccitto, A.; Tazzari, M.; Beretta, V.; Rini, F.; Miranda, C.; Greco, A.; Santinami, M.; Patuzzo, R.; Vergani, B.; Villa, A.; et al. Immunomodulatory Factors Control the Fate of Melanoma Tumor Initiating Cells. Stem Cells 2016, 34, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Rehman, K.; Zulfakar, M.H. Novel Fish Oil-based Bigel System for Controlled Drug Delivery and its Influence on Immunomodulatory Activity of Imiquimod Against Skin Cancer. Pharm. Res. 2017, 34, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.G.; Morales, M.M.; Petroianu, A. Brazilian law for scientific use of animals. Acta Cir. Bras. 2009, 24, 69–74. [Google Scholar] [CrossRef] [PubMed]
- de Sá, R.D.C.D.C.; Crisma, A.R.; Cruz, M.M.; Martins, A.R.; Masi, L.N.; do Amaral, C.L.; Curi, R.; Alonso-Vale, M.I.C. Fish oil prevents changes induced by a high-fat diet on metabolism and adipokine secretion in mice subcutaneous and visceral adipocytes. J. Physiol. 2016, 594, 6301–6317. [Google Scholar] [CrossRef] [PubMed]
- Uludag, H.; Sefton, M.V. Colorimetric assay fop cellular activity in microcapsules. Biomaterials 1990, 11, 708–712. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, E.B.; Silva, K.P.H.; Paixão, V.; Amaral, J.B.d.; Rossi, M.; Xavier-Navarro, R.A.; Barros, K.V.; Silveira, V.L.F.; Vieira, R.P.; Oliveira, L.V.F.; et al. A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment. Int. J. Mol. Sci. 2019, 20, 3765. https://doi.org/10.3390/ijms20153765
Almeida EB, Silva KPH, Paixão V, Amaral JBd, Rossi M, Xavier-Navarro RA, Barros KV, Silveira VLF, Vieira RP, Oliveira LVF, et al. A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment. International Journal of Molecular Sciences. 2019; 20(15):3765. https://doi.org/10.3390/ijms20153765
Chicago/Turabian StyleAlmeida, Ewin B., Karina P.H. Silva, Vitoria Paixão, Jônatas B. do Amaral, Marcelo Rossi, Roberta A. Xavier-Navarro, Karina V. Barros, Vera L.F. Silveira, Rodolfo P. Vieira, Luis V.F. Oliveira, and et al. 2019. "A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment" International Journal of Molecular Sciences 20, no. 15: 3765. https://doi.org/10.3390/ijms20153765
APA StyleAlmeida, E. B., Silva, K. P. H., Paixão, V., Amaral, J. B. d., Rossi, M., Xavier-Navarro, R. A., Barros, K. V., Silveira, V. L. F., Vieira, R. P., Oliveira, L. V. F., Perez, E. C., Jasiulionis, M. G., & Bachi, A. L. L. (2019). A Mixture of Polyunsaturated Fatty Acids ω-3 and ω-6 Reduces Melanoma Growth by Inhibiting Inflammatory Mediators in the Murine Tumor Microenvironment. International Journal of Molecular Sciences, 20(15), 3765. https://doi.org/10.3390/ijms20153765