
Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Prospective Consumption, Hunger, Satisfaction and Fullness
- Prospective consumption: “How much do you think you could eat?” anchored with “nothing at all” and “a lot”;
- Hunger: “How hungry do you feel?” anchored with “I am not hungry at all” and “I have never been more hungry”;
- Satisfaction: “How satisfied do you feel?” anchored with “I am completely empty” and “I cannot eat another bite”;
- Fullness: “How full do you feel?” anchored with “not at all full” and “totally full”.
2.3. Core Body Temperature and Physical Activity
2.4. Circadian Phase Estimates
2.5. Laboratory Conditions
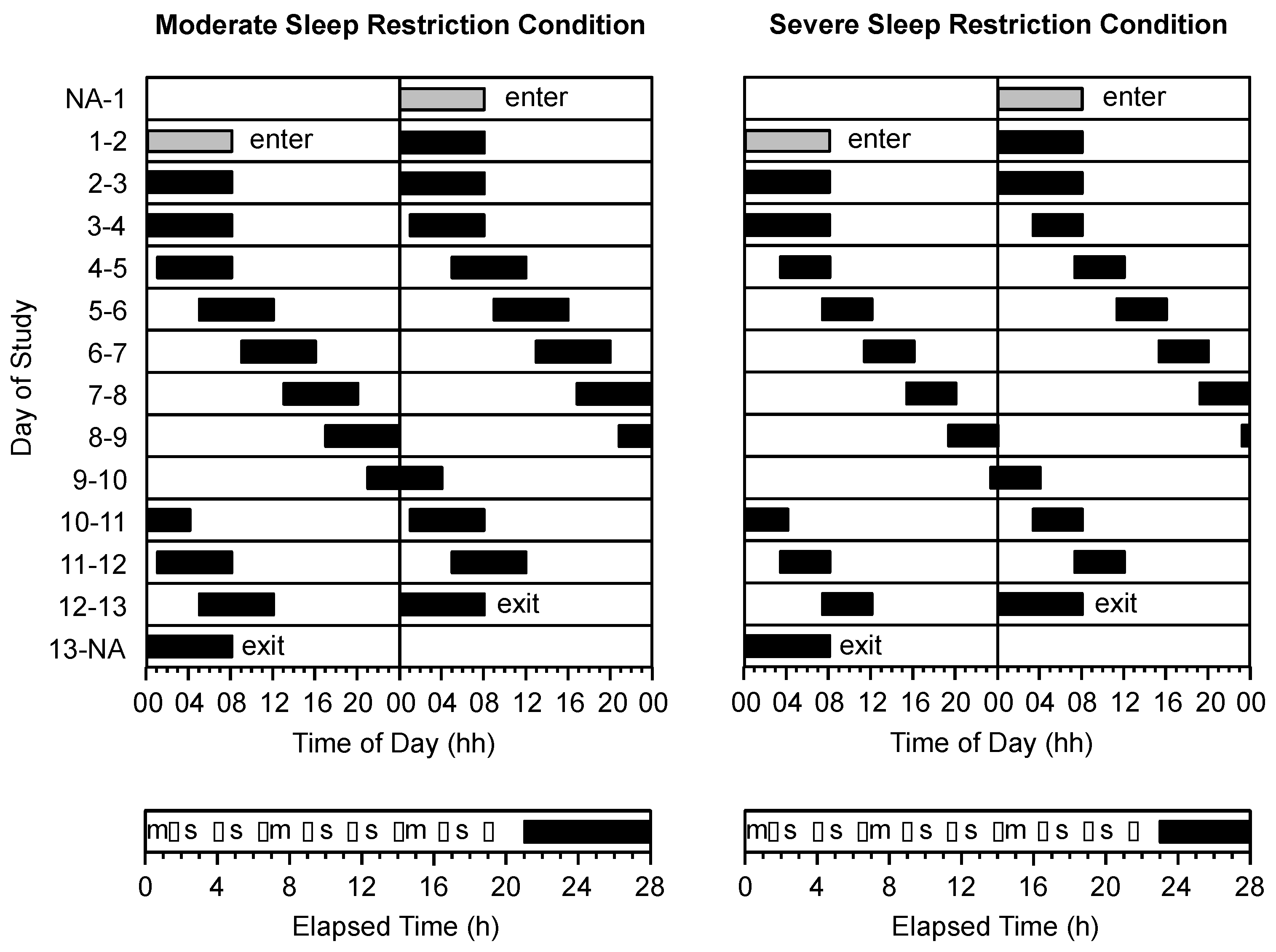
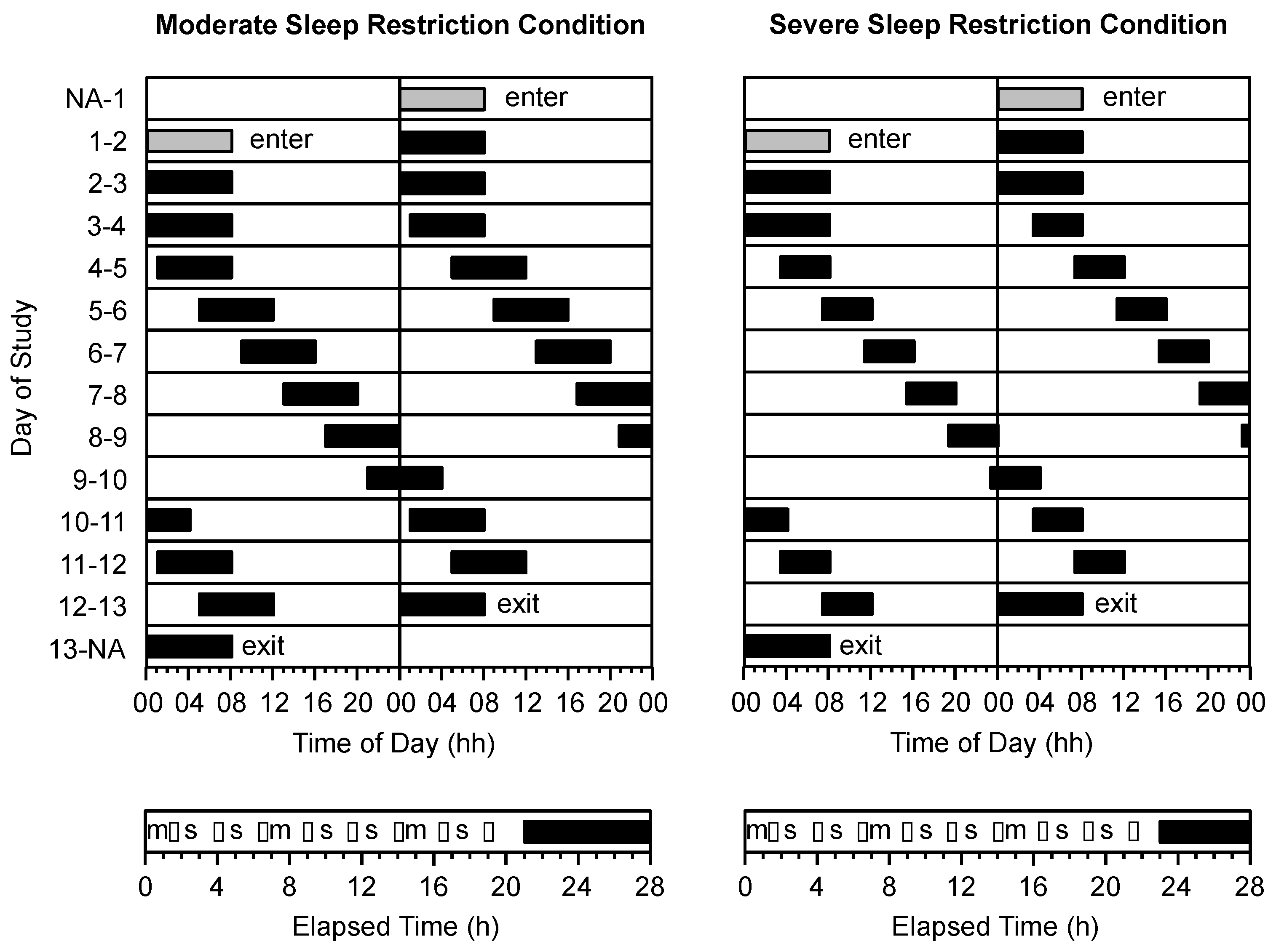
2.6. Protocol
2.7. Procedure
2.8. Meals
2.9. Statistical Analyses
3. Results
3.1. Condition
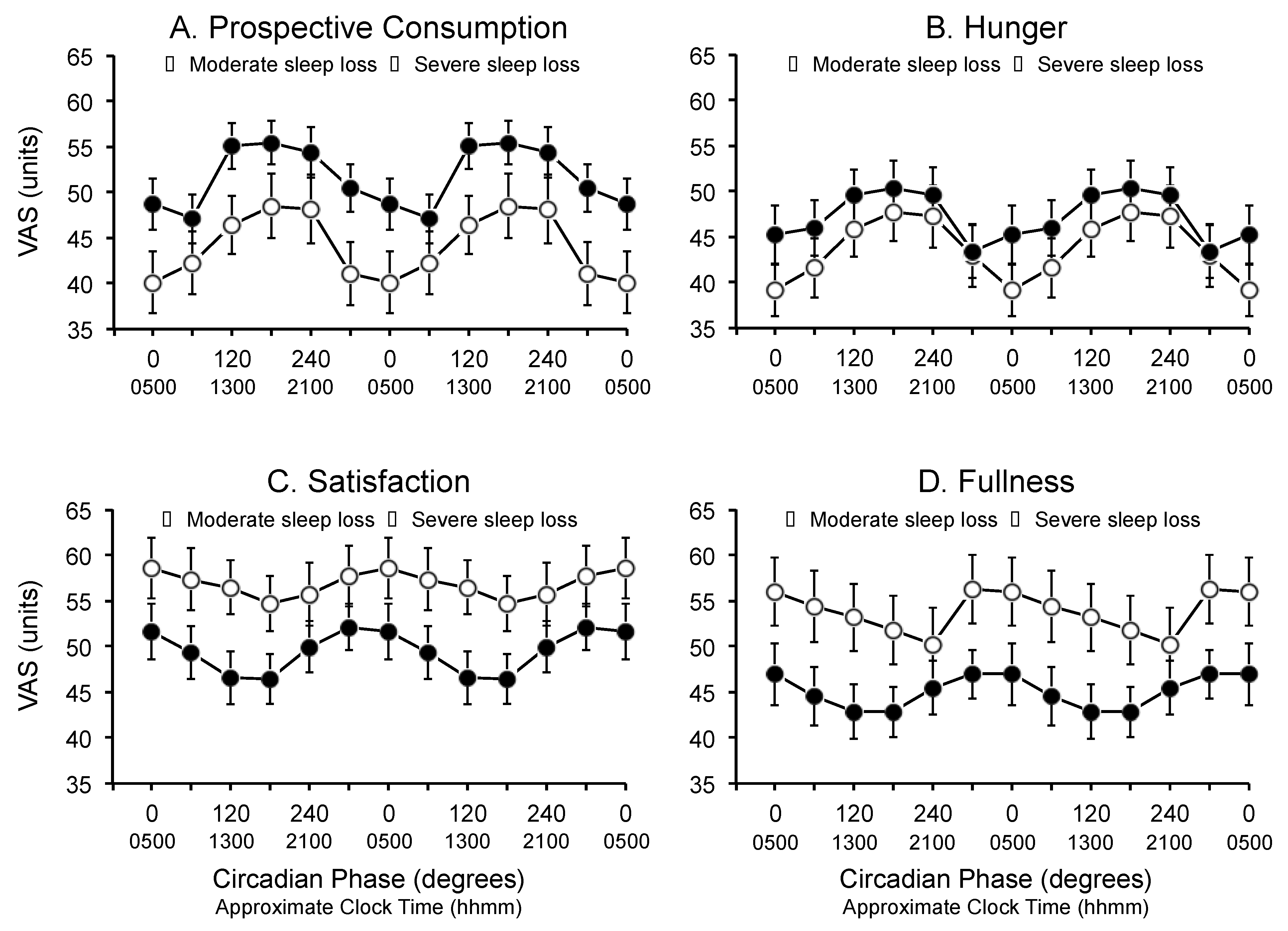
3.2. Circadian Phase
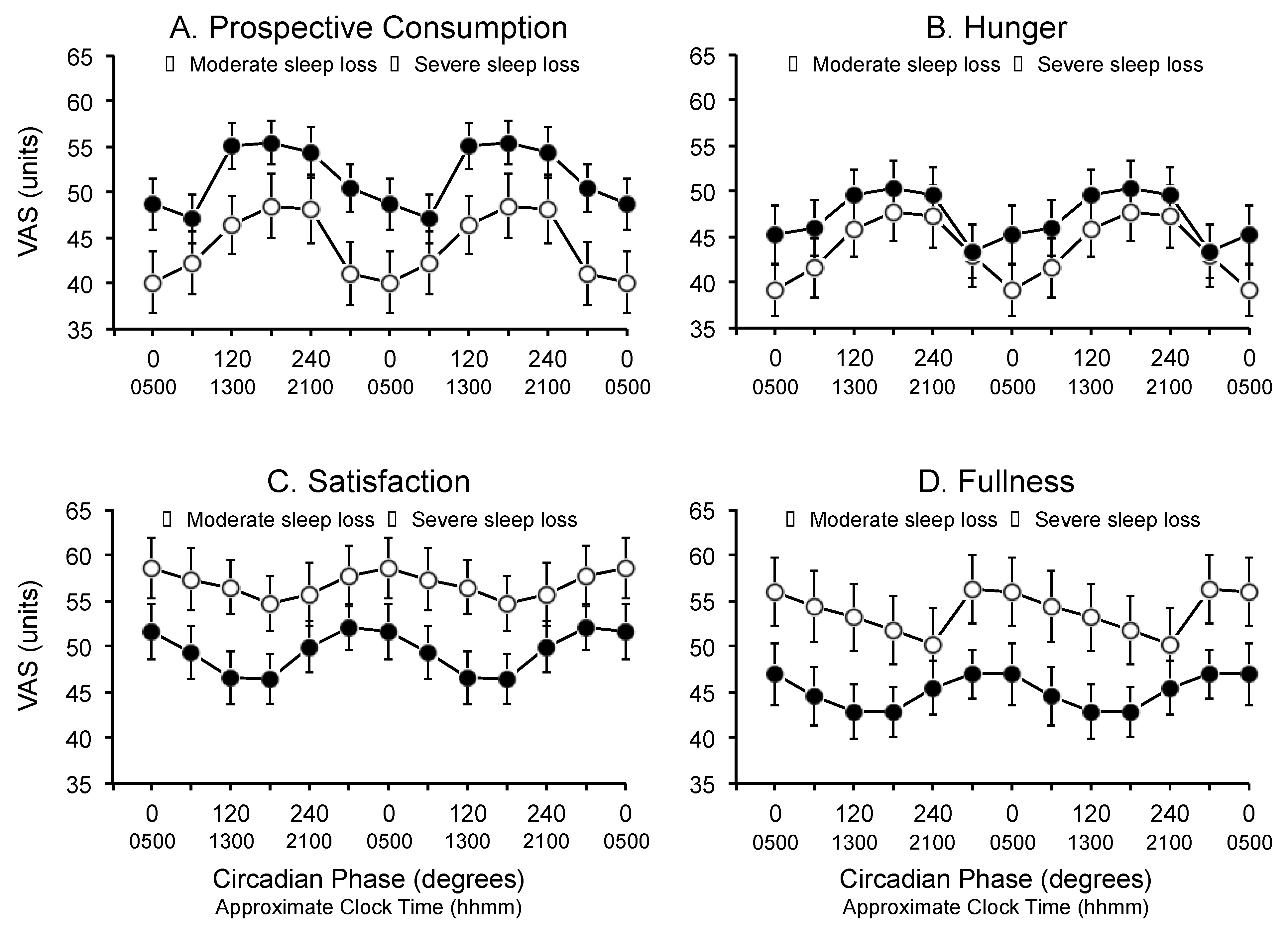
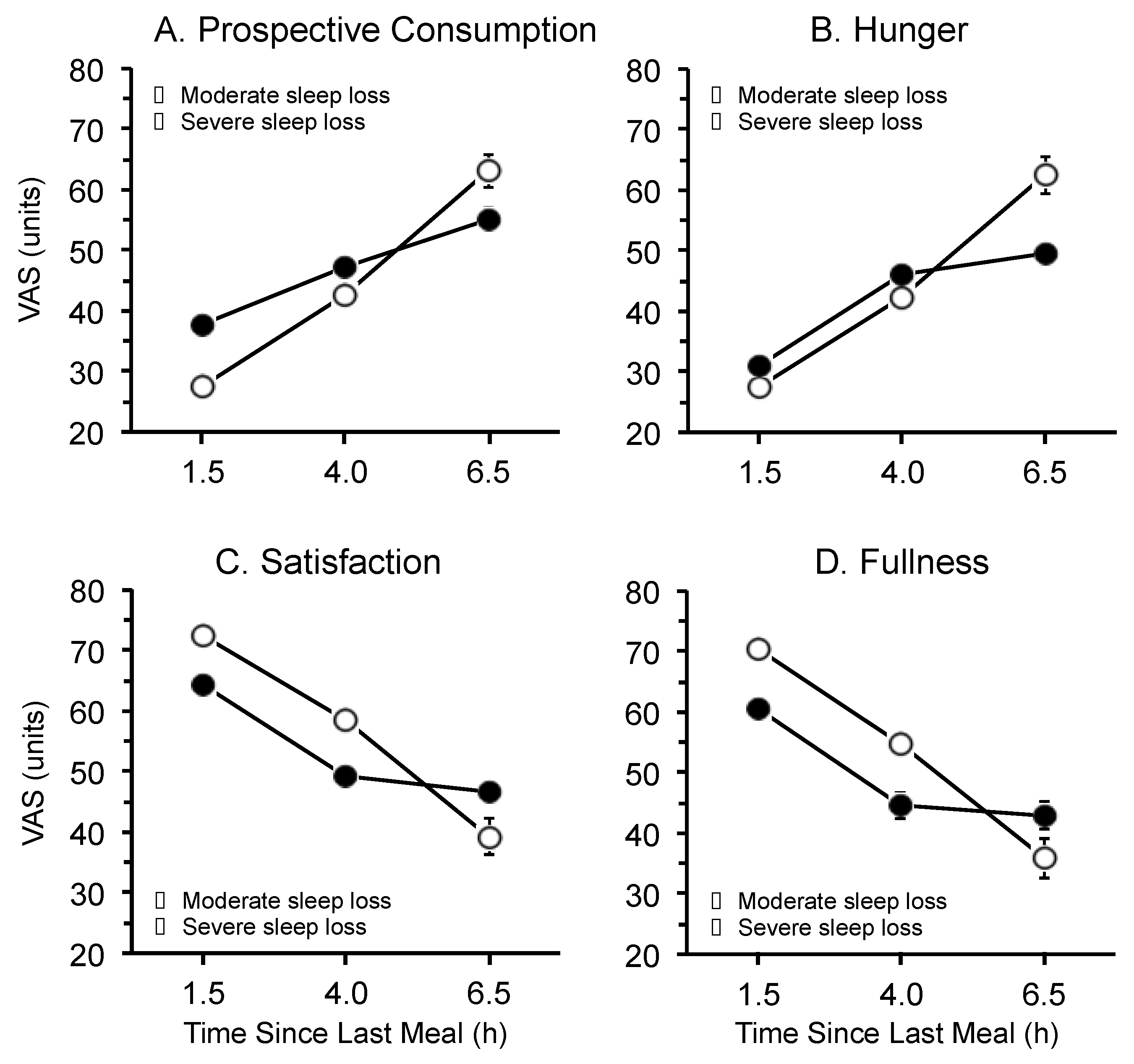
3.3. Time Since Last Meal
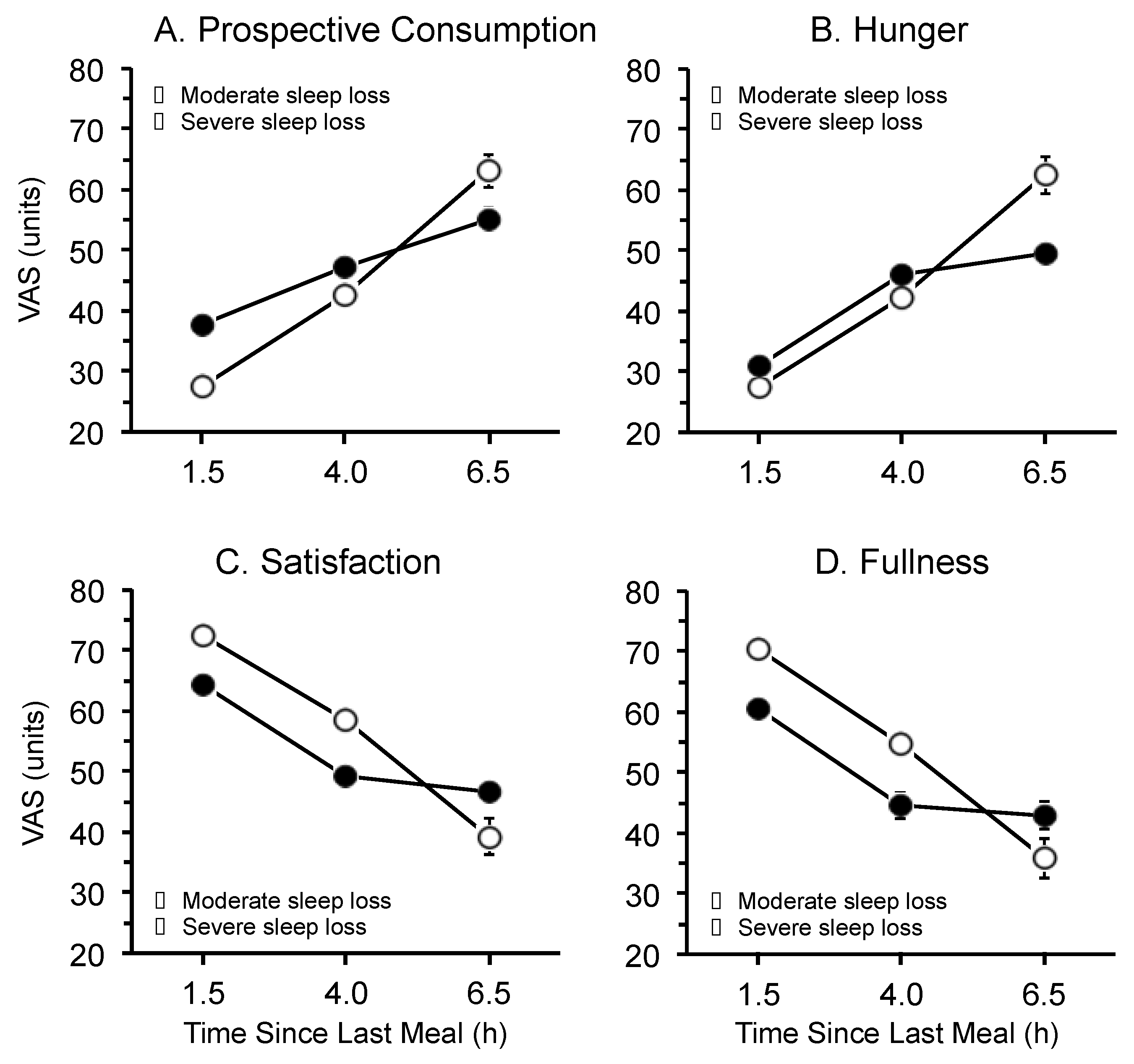
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Australian Bureau of Statistics. Working Time Arrangements, Australia, 6342.0, November 2009; Australian Bureau of Statistics: Canberra, ACT, Australia, 2009; p. 2.
- Parent-Thirion, A.; Fernández Macías, E.; Hurley, J.; Vermeylen, G. Fourth European Working Conditions Survey; European Foundation for the Improvement of Living and Working Conditions: Luxembourg, Luxembourg, 2007; pp. 22–23. [Google Scholar]
- McMenamin, T.M. A time to shift work: Recent trends in shift work and flexible schedules. Mon. Labor Rev. 2007, 130, 3–15. [Google Scholar]
- Karlsson, B.; Knutsson, A.; Lindahl, B. Is there an association between shift work and having a metabolic syndrome? Results from a population based study of 27,485 people. Occup. Environ. Med. 2001, 58, 747–752. [Google Scholar] [CrossRef]
- Pan, A.; Schernhammer, E.S.; Sun, Q.; Hu, F.B. Rotating night shift work and risk of type 2 diabetes: Two prospective cohort studies in women. PLoS Med. 2011, 8. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Power, C. Shift work and risk factors for cardiovascular disease: A study at age 45 years in the 1958 British birth cohort. Eur. J. Epidemiol. 2010, 25, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Lowden, A.; Moreno, C.; Lennernäs, M.; Tucker, P. Eating and shift work—Effects on habits, metabolism and performance. Scand. J. Work Environ. Health 2010, 36, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.J.L.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [PubMed]
- Dibner, C.; Schibler, U.; Albrecht, U. The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 2001, 72, 517–549. [Google Scholar] [CrossRef] [PubMed]
- Harrington, J.M. Health effects of shift work and extended hours of work. Occup. Environ. Med. 2001, 58, 68–72. [Google Scholar] [CrossRef]
- Rajaratnam, S.M.W.; Howard, M.E.; Grunstein, R.R. Sleep loss and circadian disruption in shift work: Health burden and management. Med. J. Aust. 2013, 199, s11–s15. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.J.L.; Morris, C.J.; Shea, S.A. The internal circadian clock increases hunger and appetite in the evening independent of food intake and other behaviours. Obesity 2013, 21, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Åkerstedt, T. Is there an optimal sleep-wake pattern in shift work? Scand. J. Work Environ. Health 1998, 24, 18–27. [Google Scholar]
- Zhou, X.; Ferguson, S.A.; Matthews, R.W.; Sargent, C.; Darwent, D.; Kennaway, D.J.; Roach, G.D. Sleep, wake and phase dependent changes in neurobehavioural function under forced desynchrony. Sleep 2011, 34, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Ferguson, S.A.; Matthews, R.W.; Sargent, C.; Darwent, D.; Kennaway, D.J.; Roach, G.D. Mismatch between subjective alertness and objective performance under sleep restriction is greatest during the biological night. J. Sleep Res. 2012, 21, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.-J.; von Schantz, M. Timing and consolidation of human sleep, wakefulness, and performance by a symphony of oscillators. J. Biol. Rhythm. 2005, 20, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Flint, A.; Raben, A.; Blundell, J.E.; Astrup, A. Reproducibility, power and validity of visual analogue scales in assessment of appetite sensations in single test meal studies. Int. J. Obes. 2000, 24, 38–48. [Google Scholar] [CrossRef]
- Stubbs, R.J.; Hughes, D.A.; Johnstone, A.M.; Rowley, E.; Reid, C.; Elia, M.; Stratton, R.; Delargy, H.; King, N.; Blundell, J.E. The use of visual analogue scales to assess motivation to eat in human subjects: A review of their reliability and validity with an evaluation of new hand-held computerized systems for temporal tracking of appetite ratings. Br. J. Nutr. 2000, 84, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, J.; Weinert, D.; Minors, D.; Folkard, S.; Owens, D.; Atkinson, G.; Macdonald, I.; Sytnik, N.; Tucker, P.; Reilly, T. A comparison of some different methods for purifying core temperature data from humans. Chronobiol. Int. 2000, 17, 539–566. [Google Scholar] [CrossRef] [PubMed]
- Darwent, D.; Ferguson, S.A.; Sargent, C.; Paech, G.M.; Williams, L.; Zhou, X.; Matthews, R.W.; Dawson, D.; Kennaway, D.J.; Roach, G.D. Contribution of core body temperature, prior wake time, and sleep stages to cognitive throughput performance during forced desynchrony. Chronobiol. Int. 2010, 27, 898–910. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.W.; Ferguson, S.A.; Zhou, X.; Sargent, C.; Darwent, D.; Kennaway, D.J.; Roach, G.D. Time-of-day mediates the influences of extended wake and sleep restriction on simulated driving. Chronobiol. Int. 2012, 29, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.W.; Ferguson, S.A.; Zhou, X.; Kosmadopoulos, A.; Kennaway, D.J.; Roach, G.D. Simulated driving under the influence of extended wake, time of day and sleep restriction. Accid. Anal. Prev. 2012, 45, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Sargent, C.; Darwent, D.; Ferguson, S.A.; Roach, G.D. Can a simple balance test be used to assess fitness for duty? Accid. Anal. Prev. 2012, 45, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Heath, G.; Roach, G.D.; Dorrian, J.; Ferguson, S.A.; Darwent, D.; Sargent, C. The effect of sleep restriction on snacking behaviour during a week of simulated shiftwork. Accid. Anal. Prev. 2012, 45, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Whitney, E.; Rolfes, S.R. Planning a healthy diet. In Understanding Nutrition, 11th ed.; Thomson Wadsworth: Belmont, CA, USA, 2008; pp. 37–63. [Google Scholar]
- Waterhouse, J.; Buckley, P.; Edwards, B.; Reilly, T. Measurement of, and some reasons for, differences in eating habits between night and day workers. Chronobiol. Int. 2003, 26, 1075–1092. [Google Scholar] [CrossRef]
- Cummings, D.E.; Purnell, J.Q.; Frayo, R.S.; Schmidova, K.; Wisse, B.E.; Weigle, D.S. A preprandial rise in plasma ghrelin levels suggests a role in meal initiation in humans. Diabetes 2001, 50, 1714–1719. [Google Scholar] [CrossRef]
- Simon, C.; Gronfier, C.; Schlienger, J.L.; Brandenberger, G. Circadian and ultradian variations of leptin in normal man under continuous enteral nutrition: Relationship to sleep and body temperature. J. Clin. Endocrinol. Metab. 1998, 83, 1893–1899. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.G.; Englert, E., Jr. Circadian rhythm of gastric acid secretion in man. Nature 1970, 226, 1261–1262. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, G.; Fullick, S.; Grindey, C.; Maclaren, D. Exercise, energy balance and the shift worker. Sports Med. 2008, 38, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, J.; Minors, D.; Atkinson, G.; Benton, D. Chronobiology and meal times: Internal and external factors. Br. J. Nutr. 1997, 77, s29–s38. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sargent, C.; Zhou, X.; Matthews, R.W.; Darwent, D.; Roach, G.D. Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment. Int. J. Environ. Res. Public Health 2016, 13, 170. https://doi.org/10.3390/ijerph13020170
Sargent C, Zhou X, Matthews RW, Darwent D, Roach GD. Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment. International Journal of Environmental Research and Public Health. 2016; 13(2):170. https://doi.org/10.3390/ijerph13020170
Chicago/Turabian StyleSargent, Charli, Xuan Zhou, Raymond W. Matthews, David Darwent, and Gregory D. Roach. 2016. "Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment" International Journal of Environmental Research and Public Health 13, no. 2: 170. https://doi.org/10.3390/ijerph13020170
APA StyleSargent, C., Zhou, X., Matthews, R. W., Darwent, D., & Roach, G. D. (2016). Daily Rhythms of Hunger and Satiety in Healthy Men during One Week of Sleep Restriction and Circadian Misalignment. International Journal of Environmental Research and Public Health, 13(2), 170. https://doi.org/10.3390/ijerph13020170