Novel Y Chromosome Retrocopies in Canids Revealed through a Genome-Wide Association Study for Sex

 ,
,
Abstract
1. Introduction
2. Materials and Methods
3. Results
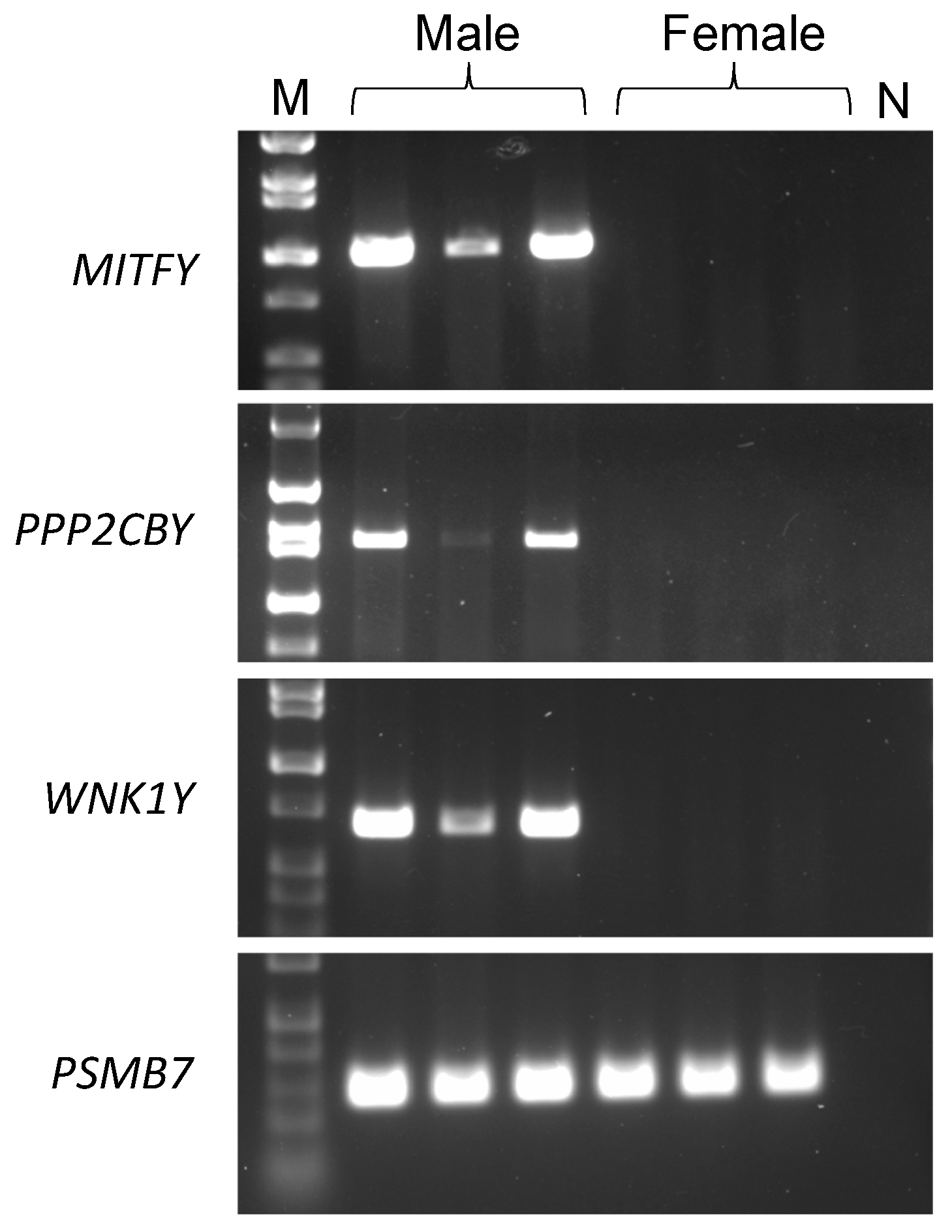
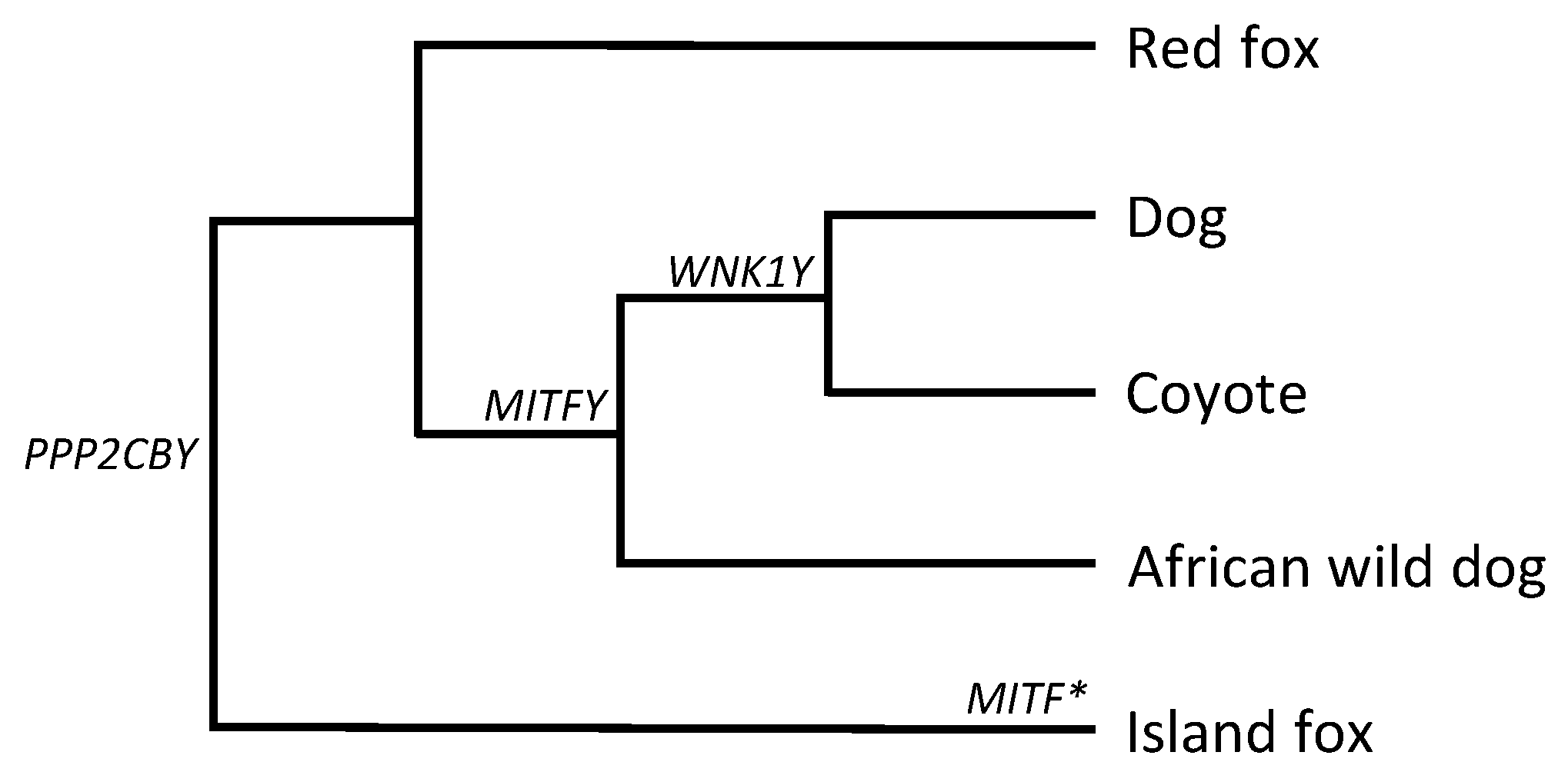
3.1. MITFY
3.2. PPP2CBY
3.3. WNK1Y
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Skaletsky, H.; Kuroda-Kawaguchi, T.; Minx, P.J.; Cordum, H.S.; Hillier, L.; Brown, L.G.; Repping, S.; Pyntikova, T.; Ali, J.; Bieri, T.; et al. The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 2003, 423, 825–837. [Google Scholar] [CrossRef]
- Yang, Y.; Chang, T.C.; Yasue, H.; Bharti, A.K.; Retzel, E.F.; Liu, W.S. ZNF280BY and ZNF280AY: autosome derived Y-chromosome gene families in Bovidae. BMC Genomics 2011, 13. [Google Scholar] [CrossRef]
- Dyer, K.A.; White, B.E.; Bray, M.J.; Piqué, D.G.; Betancourt, A.J. Molecular evolution of a Y chromosome to autosome gene duplication in Drosophila. Mol. Biol. Evol. 2011, 28, 1293–1306. [Google Scholar] [CrossRef][Green Version]
- Hughes, J.F.; Skaletsky, H.; Koutseva, N.; Pyntikova, T.; Page, D.C. Sex chromosome to autosome transposition events counter Y-chromosome gene loss in mammals. Genome Biol. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Page, D.C.; Harper, M.E.; Love, J.; Botstein, D. Occurence of a transposition from the X-chromosome long arm to the Y-chromosome short arm during human evolution. Nature 1984, 311, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Brown, L.G.; Hawkins, T.; Alagappan, R.K.; Skaletsky, H.; Reeve, M.P.; Reijo, R.; Rozen, S.; Dinulos, M.B.; Disteche, C.M.; Page, D.C. The DAZ gene cluster on the human Y chromosome arose from an autosomal gene that was transposed, repeatedly amplified and pruned. Nat. Genet. 1996, 14, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Lahn, B.T.; Page, D.D. Retroposition of autosomal mRNA yielded testis-specific gene family on human Y chromosome. Nat. Genet. 1999, 21, 429–433. [Google Scholar] [CrossRef]
- Cao, P.; Wang, L.; Jiang, Y.; Yi, Y.; Qu, F.; Liu, T.; Lv, L. De Novo origin of VCY2 from autosome to y transposed amplicon. PLoS ONE 2015, 10, e0119651. [Google Scholar] [CrossRef][Green Version]
- Carvalho, A.B.; Vicoso, B.; Russo, C.A.M.; Swenor, B.; Clark, A.G. Birth of a new gene on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2015, 112, 12450–12455. [Google Scholar] [CrossRef]
- Marques, A.C.; Dupanloup, I.; Vinckenbosch, N.; Reymond, A.; Kaessmann, H. Emergence of young human genes after a burst of retroposition in primates. PLoS ONE 2005, 3, e357. [Google Scholar] [CrossRef]
- Bai, Y.; Casola, C.; Betrán, E. Evolutionary origin of regulatory regions of retrogenes in Drosophila. BMC Genomics 2008, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J., III; Zody, M.C.; et al. Genome sequence, comparative analysis and haplotype structure of domestic dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef] [PubMed]
- Hoeppner, M.P.; Lundquist, A.; Pirun, M.; Meadows, J.R.S.; Zamani, N.; Johnson, J.; Sundström, G.; Cook, A.; FitzGerald, M.G.; Swofford, R.; Mauceli, E.; et al. An improved canine genome and a comprehensive catalogue of coding genes and non-coding transcripts. PLoS ONE 2014, 9, e91172. [Google Scholar] [CrossRef] [PubMed]
- Natanaelsson, C.; Oskarsson, M.C.R.; Angleby, H.; Lundeberg, J.; Kirkness, E.; Savolainen, P. Dog Y chromosomal DNA sequence: identification, sequencing and SNP discovery. BMC Genet. 2006, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Davis, B.W.; Raudsepp, T.; Pearks Wilkerson, A.J.; Mason, V.C.; Ferguson-Smith, M.; O’Brien, P.C.; Waters, P.D.; Murphy, W.J. Comparative analysis of mammalian Y chromosomes illuminates ancestral structure and lineage-specific evolution. Genome Res. 2013, 23, 1486–1495. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.D.; Nacu, S. Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics 2010, 7, 873–881. [Google Scholar] [CrossRef]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative genomics viewer (IGV): high-performance genomics data visualization and exploration. Brief bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef]
- Parker, H.G.; VonHoldt, B.M.; Quignon, P.; Margulies, E.H.; Shao, S.; Mosher, D.S.; Spady, T.C.; Elkahloun, A.; Cargill, M.; Jones, P.G.; et al. An expressed fgf4 retrogene is associated with breed-defining chondrodysplasia in domestic dogs. Science 2009, 325, 995–998. [Google Scholar] [CrossRef]
- Brown, E.A.; Dickinson, P.J.; Mansour, T.; Sturges, B.K.; Aguilar, M.; Young, A.E.; Korff, C.; Lind, J.; Ettinger, C.L.; Varon, S.; et al. FGF4 retrogene on CFA12 is responsible for chondrodystrophy and intervertebral disc disease in dogs. Proc. Natl. Acad. Sci. USA 2017, 114, 11476–11481. [Google Scholar] [CrossRef] [PubMed]
- Marchant, T.W.; Johnson, E.J.; McTeir, L.; Johnson, C.I.; Gow, A.; Liuti, T.; Kuehn, D.; Svenson, K.; Bermingham, M.L.; Drögemüller, M.; et al. Canine Brachycephaly is associated with a retrotransposon-mediated missplicing of SMOC2. Curr. Biol. 2017, 27, 1573–1584. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Morais, D.; Ivanga, M.; Harrison, P.M. Analysis of the role of retrotransposition in gene evolution in vertebrates. BMC Bioinformatics 2007, 8, 308. [Google Scholar] [CrossRef] [PubMed]
- Potrzebowski, L.; Vinckenbosch, N.; Marques, A.C.; Chalmel, F.; Jégou, B.; Kaessmann, H. Chromosomal gene movement reflect the recent origin and biology of therian sex chromosomes. PLoS Biol. 2008, 6, e80. [Google Scholar] [CrossRef]
- Pan, D.; Zhang, L. Burst of young retrogenes and independent retrogene formation in mammals. PLoS ONE 2009, 4, e5040. [Google Scholar] [CrossRef] [PubMed]
- Fablet, M.; Bueno, M.; Potrzebowski, L.; Kaessmann, H. Evolutionary origin and function of retrogene introns. Mol. Biol. Evol. 2009, 26, 2147–2156. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lindeboom, R.G.; Supek, F.; Lehner, B. The rules and impact of nonsense mediated RNA decay in human cancers. Nat. Genet. 2016, 48, 1112–1118. [Google Scholar] [CrossRef]
- Hemesath, T.J.; Steingrímsson, E.; McGill, G.; Hansen, M.J.; Vaught, J.; Hodgkinson, C.A.; Arnheiter, H.; Copeland, N.G.; Jenkins, N.A.; Fisher, D.E. Microphthalmia, a critical factor in melanocyte development, defines a discrete transcription factor family. Genes. Dev. 1994, 8, 2770–2780. [Google Scholar] [CrossRef]
- Saito, H.; Takeda, K.; Yasumoto, K.; Ohtani, H.; Watanabe, K.; Takahashi, K.; Fukuzaki, A.; Arai, Y.; Yamamoto, H.; Shibahara, S. Germ cell-specific expression of microphthalmia associated transcription factor mRNA in mouse testis. J. Biochem. 2003, 134, 143–150. [Google Scholar] [CrossRef]
- Hartman, M.L.; Czyz, M. MITF in melanoma: mechanisms behind its expression and activity. Cell. Mol. Life Sci. 2015, 72, 1249–1260. [Google Scholar] [CrossRef]
- Vidal-Petiot, E.; Cheval, L.; Faugeroux, J.; Malard, T.; Doucet, A.; Jeunemaitre, X.; Hadchouel, J. A new methodology for quantification of alternatively spliced exons reveals a highly tissue-specific expression pattern of WNK1 isoforms. PLoS ONE 2012, 7, e37751. [Google Scholar] [CrossRef]
- Eo, J.; Song, H.; Lim, H.J. Etv5, a transcription factor with versatile functions in male reproduction. Clin. Exp. Reprod. Med. 2012, 39, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.; Shin, S.; Janknecht, R. ETV1, 4 and 5: an oncogenic subfamily of ETS transcription factors. Biochim. Biophys. Acta 2012, 1826, 1–12. [Google Scholar] [CrossRef]
- Bai, Y.; Casola, C.; Feschotte, C.; Betrán, E. Comparative genomics reveals a constant rate of origination and convergent acquisition of functional retrogenes in Drosophila. Genome Biol. 2007, 8, R11. [Google Scholar] [CrossRef]
- Mouse Genome Sequencing Consortium. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef]
- Rosikiewicz, W.; Kabza, M.; Kosiński, J.G.; Ciomborowska-Basheer, J.; Kubiak, M.R.; Makałowska, I. Retrogene DB — a database of plant and animal retrocopies. Database 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- O’Bryan, M.K.; Grealy, A.; Stahl, P.J.; Schlegel, P.N.; McLachlan, R.I.; Jamsai, D. Genetic variants in the ETV5 gene in fertile and infertile men with nonobstructive azoospermia associated with Sertoli cell-only syndrome. Fertil. Steril. 2012, 98, 827–835. [Google Scholar] [CrossRef]
- Su, Y.; Sugiura, K.; Sun, F.; Pendola, J.K.; Cox, G.A.; Handel, M.A.; Schimenti, J.C.; Eppig, J.J. MARF1 regulates essential oogenic processes in mice. Science 2012, 335, 1496–1499. [Google Scholar] [CrossRef]
- Shekarabi, M.; Lafrenière, R.G.; Gaudet, R.; Laganière, J.; Marcinkiewicz, M.M.; Dion, P.A.; Rouleau, G.A. Comparative analysis of the expression profile of Wnk1 and Wnk1/Hsn2 splice variants in developing and adult mouse tissues. PLoS ONE 2013, 8, e57807. [Google Scholar] [CrossRef] [PubMed]
- Rohozinski, J. Lineage-independent retrotransposition of UTP14 associated with male fertility has occurred multiple times throughout mammalian evolution. R. Soc. Open Sci. 2017, 4, 171049. [Google Scholar] [CrossRef] [PubMed]
- Ciomborowska, J.; Rosikiewicz, W.; Szklarczyk, D.; Makałowski, W.; Makałowska, I. “Orphan” retrogenes in the human genome. Mol. Biol. Evol. 2012, 30, 384–396. [Google Scholar] [CrossRef] [PubMed]
- Skinner, B.M.; Sargent, C.A.; Churcher, C.; Hunt, T.; Herrero, J.; Loveland, J.E.; Dunn, M.; Louzada, S.; Fu, B.; Chow, W. The pig X and Y chromosomes: structure, sequence, and evolution. Genome Res. 2016, 26, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Chang, T.C.; Klabnik, J.L.; Liu, W.S. Regional selection acting on the OFD1 gene family. PLoS ONE 2011, 6, e26195. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Position | Gene | Region | Chr | Position | Gene | Region |
|---|---|---|---|---|---|---|---|
| 6 | 72952463 | 20 | 21870155 | MITF | exonic | ||
| 72964379 | 21870230 | MITF | exonic | ||||
| 15 | 111194 | 21870310 | MITF | exonic | |||
| 29973349 | 21870623 | MITF | 3′ UTR | ||||
| 16 | 33566554 | PPP2CB | 3′ UTR | 21871904 | MITF | 3′ UTR | |
| 33566935 | PPP2CB | 3′ UTR | 21872335 | MITF | 3′ UTR | ||
| 17 | 64209287 | 21872815 | MITF | 3′ UTR | |||
| 19 | 20034966 | 21873532 | MITF | 3′ UTR | |||
| 20040153 | 27 | 42911747 | WNK1 | exonic | |||
| 20052361 | 42911917 | WNK1 | exonic | ||||
| 20062503 | 32 | 38734101 | |||||
| 20072295 | 38767343 | ||||||
| 20100030 | 38788489 | ||||||
| 20114792 | 38789367 | ||||||
| 20130159 | X | 6604781 | SHROOM2 | exonic | |||
| 20152245 | 6621021 | SHROOM2 | intronic | ||||
| 20172164 | 6628533 | SHROOM2 | intronic | ||||
| 20245553 | 6634742 | SHROOM2 | intronic | ||||
| 20256174 | 10131021 | TRAPPC2 | exonic | ||||
| 20286580 | 10175834 | OFD1 | exonic | ||||
| 20292945 | 35604689 | USP9X/Y | exonic | ||||
| 20303661 | 57139861 | ||||||
| 20309777 | 60395963 | PGK1 | |||||
| 20310190 | Y | 26641 | |||||
| 20314276 | 316950 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, K.L.; Evans, J.M.; Noorai, R.E.; Starr-Moss, A.N.; Clark, L.A. Novel Y Chromosome Retrocopies in Canids Revealed through a Genome-Wide Association Study for Sex. Genes 2019, 10, 320. https://doi.org/10.3390/genes10040320
Tsai KL, Evans JM, Noorai RE, Starr-Moss AN, Clark LA. Novel Y Chromosome Retrocopies in Canids Revealed through a Genome-Wide Association Study for Sex. Genes. 2019; 10(4):320. https://doi.org/10.3390/genes10040320
Chicago/Turabian StyleTsai, Kate L., Jacquelyn M. Evans, Rooksana E. Noorai, Alison N. Starr-Moss, and Leigh Anne Clark. 2019. "Novel Y Chromosome Retrocopies in Canids Revealed through a Genome-Wide Association Study for Sex" Genes 10, no. 4: 320. https://doi.org/10.3390/genes10040320
APA StyleTsai, K. L., Evans, J. M., Noorai, R. E., Starr-Moss, A. N., & Clark, L. A. (2019). Novel Y Chromosome Retrocopies in Canids Revealed through a Genome-Wide Association Study for Sex. Genes, 10(4), 320. https://doi.org/10.3390/genes10040320