Retinol Saturase Knock-Out Mice are Characterized by Impaired Clearance of Apoptotic Cells and Develop Mild Autoimmunity

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Experimental Animals
2.3. Flow Cytometry Analysis of Freshly Isolated Thymocytes
2.4. Retinoid Measurement by High-performance Liquid Chromatography Mass Spectrometry (HPLC/MS/MS)
2.5. Thymocyte Apoptosis in Vitro
2.6. Bone-Marrow-Derived (BMDM), Peritoneal, or Thioglycolate-elicited Macrophage Generation, Cell Culture, and Treatment
2.7. In Vitro Apoptotic Cell Phagocytosis
2.8. Confocal Microscopy
2.9. mRNA Sequencing
2.10. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis of mRNA Expression
2.11. Anti-Nuclear Antibody Detection by Indirect Immunofluorescence Assay
2.12. Anti-dsDNA Antibody ELISA
2.13. Caspase-3 Immunohistochemistry
2.14. Detection of IgM-Containing Immune Complexes
2.15. Determination of Serum Urea Concentration
2.16. Statistical Analysis
3. Results
3.1. Loss of Retinol Saturase Does Not Affect the Induction of Retinoid-Regulated Genes or the Thymic Apopto-Phagocytosis Program
3.2. RetSat-Null Macrophages Are Characterized by Impaired Long-Term Phagocytosis
3.3. RetSat-Null Macrophages Express Less MFG-E8
3.4. Lower MFG-E8 Production Is Responsible for the Defect in Long-Term Phagocytosis of RetSat-Null Macrophages
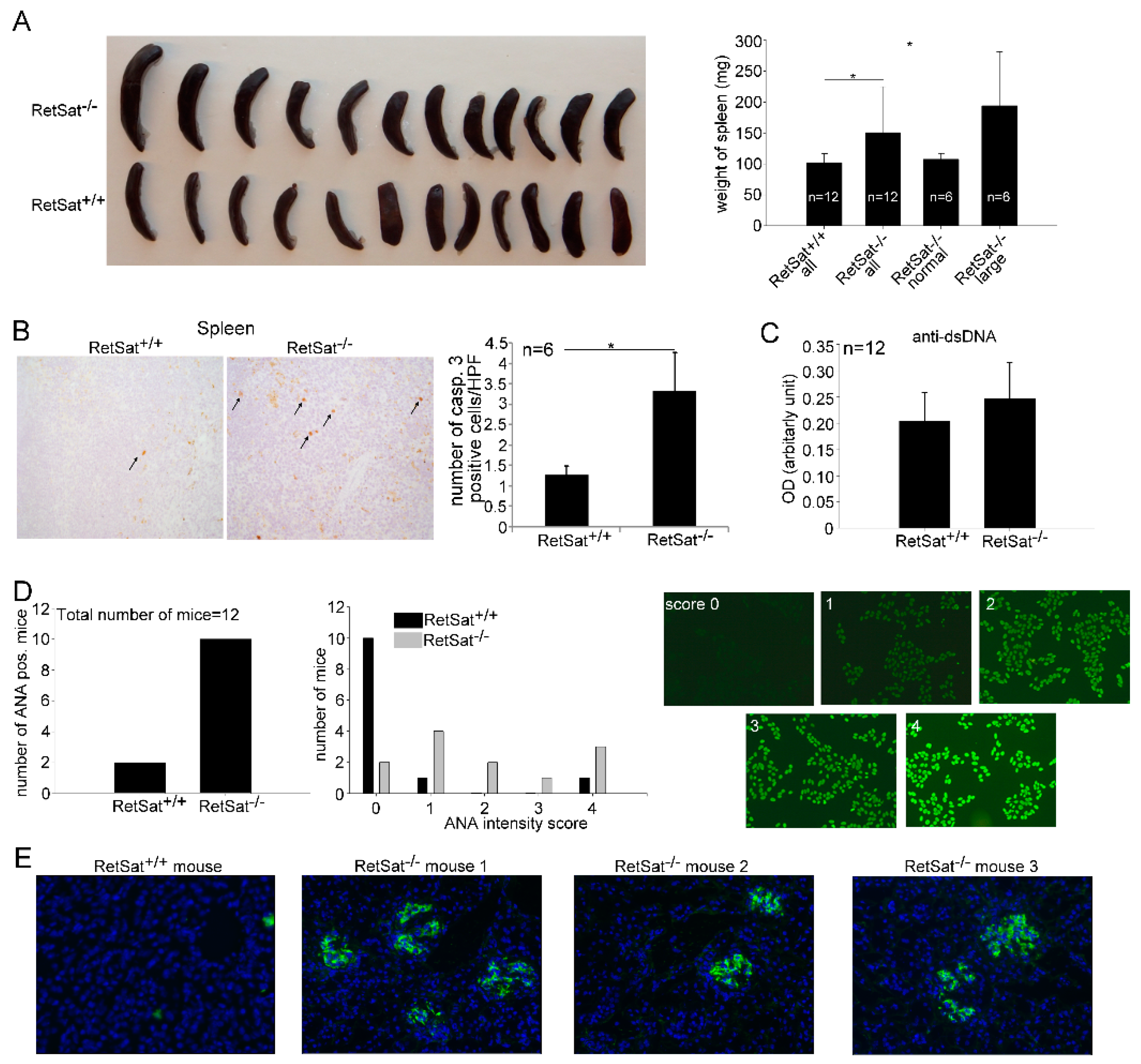
3.5. Female RetSat-Null Mice Are Prone To Develop Mild Systemic Lupus Erythematosus (SLE)-Like Autoimmunity
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arandjelovic, S.; Ravichandran, K.S. Phagocytosis of apoptotic cells in homeostasis. Nat. Immunol. 2015, 16, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Fadok, V.A.; Henson, P.M. Apoptosis: Giving phosphatidylserine recognition an assist--with a twist. Curr. Biol. 2003, 13, R655–R657. [Google Scholar] [CrossRef]
- Park, D.; Tosello-Trampont, A.C.; Elliott, M.R.; Lu, M.; Haney, L.B.; Ma, Z.; Klibanov, A.L.; Mandell, J.W.; Ravichandran, K.S. BAI1 is an engulfment receptor for apoptotic cells upstream of the ELMO/Dock180/Rac module. Nature 2007, 450, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jung, M.Y.; Kim, H.J.; Lee, S.J.; Kim, S.Y.; Lee, B.H.; Kwon, T.H.; Park, R.W.; Kim, I.S. Rapid cell corpse clearance by stabilin-2, a membrane phosphatidylserine receptor. Cell Death Differ. 2008, 15, 192–201. [Google Scholar] [CrossRef]
- Miyanishi, M.; Tada, K.; Koike, M.; Uchiyama, Y.; Kitamura, T.; Nagata, S. Identification of Tim4 as a phosphatidylserine receptor. Nature 2007, 450, 435–439. [Google Scholar] [CrossRef]
- Hanayama, R.; Tanaka, M.; Miwa, K.; Shinohara, A.; Iwamatsu, A.; Nagata, S. Identification of a factor that links apoptotic cells to phagocytes. Nature 2002, 417, 182–187. [Google Scholar] [CrossRef]
- Savill, J.S.; Hogg, N.; Ren, Y.; Haslett, C. Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis. J. Clin. Invest. 1992, 90, 1513–1522. [Google Scholar] [CrossRef]
- Stitt, T.N.; Conn, G.; Gore, M.; Lai, C.; Bruno, J.; Radziejewski, C.; Mattsson, K.; Fisher, J.; Gies, D.R.; Jones, P.F.; et al. The anticoagulation factor protein S and its relative, Gas6, are ligands for the Tyro 3/Axl family of receptor tyrosine kinases. Cell 1995, 80, 661–670. [Google Scholar] [CrossRef]
- Botto, M.; Dell’Agnola, C.; Bygrave, A.E.; Thompson, E.M.; Cook, H.T.; Petry, F.; Loos, M.; Pandolfi, P.P.; Walport, M.J. Homozygous C1q deficiency causes glomerulonephritis associated with multiple apoptotic bodies. Nat. Genet. 1998, 19, 56–59. [Google Scholar] [CrossRef]
- Park, D.; Hochreiter-Hufford, A.; Ravichandran, K.S. The phosphatidylserine receptor TIM-4 does not mediate direct signaling. Curr. Biol. 2009, 19, 346–351. [Google Scholar] [CrossRef]
- Devitt, A.; Parker, K.G.; Ogden, C.A.; Oldreive, C.; Clay, M.F.; Melville, L.A.; Bellamy, C.O.; Lacy-Hulbert, A.; Gangloff, S.C.; Goyert, S.M.; et al. Persistence of apoptotic cells without autoimmune disease or inflammation in CD14−/− mice. J. Cell. Biol. 2004, 167, 1161–1170. [Google Scholar] [CrossRef]
- Greenberg, M.E.; Sun, M.; Zhang, R.; Febbraio, M.; Silverstein, R.; Hazen, S.L. Oxidized phosphatidylserine-CD36 interactions play an essential role in macrophage-dependent phagocytosis of apoptotic cells. J. Exp. Med. 2006, 203, 2613–2625. [Google Scholar] [CrossRef]
- Cohen, P.L.; Caricchio, R.; Abraham, V.; Camenisch, T.D.; Jennette, J.C.; Roubey, R.A.; Earp, H.S.; Matsushima, G.; Reap, E.A. Delayed apoptotic cell clearance and lupus-like autoimmunity in mice lacking the c-Mertk membrane tyrosine kinase. J. Exp. Med. 2002, 196, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Tóth, B.; Garabuczi, E.; Sarang, Z.; Vereb, G.; Vámosi, G.; Aeschlimann, D.; Blaskó, B.; Bécsi, B.; Erdődi, F.; Lacy-Hulbert, A.; et al. Transglutaminase 2 is needed for the formation of an efficient phagocyte portal in macrophages engulfing apoptotic cells. J. Immunol. 2009, 182, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Kinchen, J.M.; Cabello, J.; Klingle, D.; Wong, K.; Freichtinger, R.; Schnabel, H.; Schnabel, R.; Hengartner, M.O. Two pathways converge at CED-10 to mediate actin rearrangement and corpse removal in C. elegans, Nature 2005, 43, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Miksa, M.; Amin, D.; Wu, R.; Jacob, A.; Zhou, M.; Dong, W.; Yang, W.L.; Ravikumar, T.S.; Wang, P. Maturation-induced down-regulation of MFG-E8 impairs apoptotic cell clearance and enhances endotoxin response. Int. J. Mol. Med. 2008, 22, 743–748. [Google Scholar] [PubMed]
- Noelia, A.; Bensinger, S.J.; Hong, C.; Beceiro, S.; Bradley, M.N.; Zelcer, N.; Deniz, J.; Ramirez, C.; Díaz, M.; Gallardo, G.; et al. Apoptotic cells promote their own clearance and immune tolerance through activation of the nuclear receptor LXR. Immunity 2009, 31, 245–258. [Google Scholar] [CrossRef]
- Roszer, T.; Menéndez-Gutiérrez, M.P.; Lefterova, M.I.; Alameda, D.; Núñez, V.; Lazar, M.A.; Fischer, T.; Ricote, M. Autoimmune kidney disease and impaired engulfment of apoptotic cells in mice with macrophage peroxisome proliferator-activated receptor gamma or retinoid X receptor alpha deficiency. J. Immunol. 2011, 186, 621–631. [Google Scholar] [CrossRef]
- Mukundan, L.; Odegaard, J.I.; Morel, C.R.; Heredia, J.E.; Mwangi, J.W.; Ricardo-Gonzalez, R.R.; Goh, Y.P.; Eagle, A.R.; Dunn, S.E.; Awakuni, J.U.; et al. PPAR-delta senses and orchestrates clearance of apoptotic cells to promote tolerance. Nat. Med. 2009, 15, 1266–1272. [Google Scholar] [CrossRef]
- Garabuczi, É.; Kiss, B.; Felszeghy, S.; Tsay, G.J.; Fésüs, L.; Szondy, Z. Retinoids produced by macrophages engulfing apoptotic cells contribute to the appearance of transglutaminase 2 in apoptotic thymocytes. Amino Acids 2013, 44, 235–244. [Google Scholar] [CrossRef]
- Sarang, Z.; Joós, G.; Garabuczi, É.; Rühl, R.; Gregory, C.D.; Szondy, Z. Macrophages engulfing apoptotic cells produce nonclassical retinoids to enhance their phagocytic capacity. J. Immunol. 2014, 192, 5730–5738. [Google Scholar] [CrossRef] [PubMed]
- Moise, A.R.; Domínguez, M.; Alvarez, S.; Alvarez, R.; Schupp, M.; Cristancho, A.G.; Kiser, P.D.; de Lera, A.R.; Lazar, M.A.; Palczewski, K. Stereospecificity of retinol saturase: Absolute configuration, synthesis, and biological evaluation of dihydroretinoids. J. Am. Chem. Soc. 2008, 130, 1154–1155. [Google Scholar] [CrossRef] [PubMed]
- Moise, A.R.; Alvarez, S.; Domínguez, M.; Alvarez, R.; Golczak, M.; Lobo, G.P.; von Lintig, J.; de Lera, A.R.; Palczewski, K. Activation of retinoic acid receptors by dihydroretinoids. Mol. Pharmacol. 2009, 76, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Krężel, W.; Rühl, R.; de Lera, A.R. Alternative retinoid X receptor (RXR) ligands. Mol. Cell. Endocrinol. 2019, 491, 110436. [Google Scholar] [CrossRef] [PubMed]
- Schupp, M.; Lefterova, M.; Janke, J.; Leitner, K.; Cristancho, A.G.; Mullican, S.E.; Qatanani, M.; Szwergold, N.; Steger, D.J.; Curtin, J.C.; et al. Retinol saturase promotes adipogenesis and is downregulated in obesity. Proc. Natl Acad. Sci. USA. 2009, 106, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.Y.; Wang, S.; Jurczak, M.J.; Shulman, G.I.; Moise, A.R. Retinolsaturase modulates lipid metabolism and the production of reactive oxygen species. Arch. Biochem. Biophys. 2017, 633, 93–102. [Google Scholar] [CrossRef]
- Heidenreich, S.; Witte, N.; Weber, P.; Goehring, I.; Tolkachov, A.; von Loeffelholz, C.; Döcke, S.; Bauer, M.; Stockmann, M.; Pfeiffer, A.F.H.; et al. Retinolsaturase coordinates liver metabolism by regulating ChREBP activity. Nat. Commun. 2017, 8, 384. [Google Scholar] [CrossRef]
- Moise, A.R.; Lobo, G.P.; Erokwu, B.; Wilson, D.L.; Peck, D.; Alvarez, S.; Domínguez, M.; Alvarez, R.; Flask, C.A.; de Lera, A.R.; et al. Increased adiposity in the retinol saturase-knockout mouse. FASEB J. 2010, 24, 1261–1270. [Google Scholar] [CrossRef]
- Lauber, K.; Keppeler, H.; Munoz, L.E.; Koppe, U.; Schröder, K.; Yamaguchi, H.; Krönke, G.; Uderhardt, S.; Wesselborg, S.; Belka, C.; et al. Milk fat globule-EGF factor 8 mediates the enhancement of apoptotic cell clearance by glucocorticoids. Cell Death Differ. 2013, 20, 1230–1240. [Google Scholar] [CrossRef]
- Rühl, R. Method to determine 4-oxo-retinoic acids, retinoic acids and retinol in serum and cell extracts by liquid chromatography/diode-array detection atmospheric pressure chemical ionisation tandem mass spectrometry. Rapid. Commun. Mass Spectrum 2006, 20, 2497–2504. [Google Scholar] [CrossRef]
- Rahmatullah, M.; Boyde, T.R. Improvements in the determination of urea using diacetyl monoxime; methods with and without deproteinisation. Clin. Chim. Acta. 1980, 107, 3–9. [Google Scholar] [CrossRef]
- Starr, T.K.; Jameson, S.C.; Hogquist, K.A. Positive and negative selection of T Cells. Ann. Rev. Immunol. 2003, 21, 139–176. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Konkel, J.E. Development of thymic Foxp3(+) regulatory T cells: TGF-β matters. Eur. J. Immunol. 2015, 45, 958–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarang, Z.; Garabuczi, É.; Joós, G.; Kiss, B.; Tóth, K.; Rühl, R.; Szondy, Z. Macrophages engulfing apoptotic thymocytes produce retinoids to promote selection, differentiation, removal and replacement of double positive thymocytes. Immunobiology 2013, 218, 1354–1360. [Google Scholar] [CrossRef]
- Buttari, B.; Profumo, E.; Darcangelo, D.; Di Raimo, T.; Businaro, R.; Capoano, R.; Salvati, B.; Saso, L.; Elenkov, I. Neuropeptide Y as regulator of macrophage phenotype and functions: A neuroimmune CUE in atherosclerosis regression? Atherosclerosis 2017, 263, e2. [Google Scholar] [CrossRef]
- Zizzo, G.; Hilliard, B.A.; Monestier, M.; Cohen, P.L. Efficient clearance of early apoptotic cells by human macrophages requires M2c polarization and MerTK induction. J. Immunol. 2012, 189, 3508–3520. [Google Scholar] [CrossRef] [Green Version]
- Pedragosa-Badia, X.; Stichel, J.; Beck-Sickinger, A.G. Neuropeptide Y receptors: How to get subtype selectivity. Front. Endocrinol. 2013, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Hanayama, R.; Tanaka, M.; Miyasak, K.; Aozasa, K.; Koike, M.; Uchiyama, Y.; Nagata, S. Autoimmune disease and impaired uptake of apoptotic cells in MFG-E8-deficient mice. Science 2004, 304, 1147–1150. [Google Scholar] [CrossRef] [Green Version]
- Abdolmaleki, F.; Farahani, N.; Gheibi Hayat, S.M.; Pirro, M.; Bianconi, V.; Barreto, G.E.; Sahebkar, A. The Role of Efferocytosis in Autoimmune Diseases. Front. Immunol. 2018, 9, 1645. [Google Scholar] [CrossRef] [Green Version]
- Mevorach, D.; Zhou, J.L.; Song, X.; Elkon, K.B. Systemic exposure to irradiated apoptotic cells induces autoantibody production. J. Exp. Med. 1998, 188, 387–392. [Google Scholar] [CrossRef] [Green Version]
- Weckerle, C.E.; Niewold, T.B. The Unexplained Female Predominance of Systemic Lupus Erythematosus: Clues from Genetic and Cytokine Studies. Clin. Rev. Allergy Immunol. 2011, 40, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szondy, Z.; Garabuczi, E.; Joós, G.; Tsay, G.J.; Sarang, Z. Impaired clearance of apoptotic cells in chronic inflammatory diseases: Therapeutic implications. Front. Immunol. 2014, 5, 354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GeneAtlas MOE430, gcrma. Available online: http://ds.biogps.org/?dataset=GSE10246&gene=67442 (accessed on 14 October 2019).
- Matsuda, A.; Jacob, A.; Wu, R.; Zhou, M.; Nicastro, J.M.; Coppa, G.F.; Wang, P. Milk fat globule-EGF factor VIII in sepsis and ischemia-reperfusion injury. Mol. Med. 2011, 17, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Michalski, M.N.; Seydel, A.L.; Siismets, E.M.; Zweifler, L.E.; Koh, A.J.; Sinder, B.P.; Aguirre, J.I.; Atabai, K.; Roca, H.; McCauley, L.K. Inflammatory bone loss associated with MFG-E8 deficiency is rescued by teriparatide. FASEB J. 2018, 32, 3730–3741. [Google Scholar] [CrossRef] [Green Version]
- Szondy, Z.; Sarang, Z.; Molnar, P.; Nemeth, T.; Piacentini, M.; Mastroberardino, P.G.; Falasca, L.; Aeschlimann, D.; Kovacs, J.; Kiss, I.; et al. Transglutaminase 2−/− mice reveal a phagocytosis-associated crosstalk between macrophages and apoptotic cells. Proc. Natl. Acad. Sci. USA 2003, 100, 7812–7817. [Google Scholar] [CrossRef] [Green Version]
- Dimitrijević, M.; Stanojević, S.; Mitić, K.; Kustrimović, N.; Vujić, V.; Miletić, T.; Kovacević-Jovanović, V. The anti-inflammatory effect of neuropeptide Y (NPY) in rats is dependent on dipeptidyl peptidase 4 (DP4) activity and age. Peptides 2008, 29, 2179–2187. [Google Scholar]
- Soki, F.N.; Koh, A.J.; Jones, J.D.; Kim, Y.W.; Dai, J.; Keller, E.T.; Pienta, K.J.; Atabai, K.; Roca, H.; McCauley, L.K. Polarization of prostate cancer-associated macrophages is induced by milk fat globule-EGF factor 8 (MFG-E8)-mediated efferocytosis. J. Biol. Chem. 2014, 289, 24560–24572. [Google Scholar] [CrossRef] [Green Version]
- Atri, C.; Guerfali, F.Z.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Jinushi, M.; Nakazaki, Y.; Dougan, M.; Carrasco, D.R.; Mihm, M.; Dranoff, G. MFG-E8-mediated uptake of apoptotic cells by APCs links the pro- and antiinflammatory activities of GM-CSF. J. Clin. Invest. 2007, 117, 1902–1913. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S.; Locati, M.; Mantovani, A. Transcriptional profiling of the human monocyte-to-macrophage differentiation and polarization: New molecules and patterns of gene expression. J. Immunol. 2006, 177, 7303–7311. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Singh, L.S.; Zhang, L.; Xu, Y. Role of OGR1 in myeloid-derived cells in prostate cancer. Oncogene 2014, 33, 157–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifeh-Soltani, A.; McKleroy, W.; Sakuma, S.; Cheung, Y.Y.; Tharp, K.; Qiu, Y.; Turner, S.M.; Chawla, A.; Stahl, A.; Atabai, K. Mfge8 promotes obesity by mediating the uptake of dietary fats and serum fatty acids. Nat. Med. 2014, 20, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifeh-Soltani, A.; Gupta, D.; Ha, A.; Iqbal, J.; Hussain, M.; Podolsky, M.J.; Atabai, K. Mfge8 regulates enterocyte lipid storage by promoting enterocyte triglyceride hydrolase activity. JCI Insight. 2016, 1, e87418. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, S.E.; Grijalva, A.; Xu, X.; Ables, E.; Nomani, A.; Ferrante, A.W. A lipase-independent pathway of lipid release and immune modulation by adipocytes. Science 2019, 363, 989–993. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell. Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, N.P.; Boyd, A.S.; Fugazza, C.; May, G.E.; Guo, Y.; Tipping, A.J.; Scadden, D.T.; Vyas, P.; Enver, T. GATA-2 regulates granulocyte-macrophage progenitor cell function. Blood 2008, 112, 4862–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, W.; Jin, Z.; Zhe, G.; Song, H. The methylation of C/EBP β gene promoter and regulated by GATA-2 protein. Mol. Biol. Rep. 2013, 40, 797–801. [Google Scholar] [CrossRef]
- Aziz, M.M.; Ishihara, S.; Rumi, M.A.; Mishima, Y.; Oshima, N.; Kadota, C.; Moriyama, I.; Li, Y.Y.; Rahman, F.B.; Otani, A.; et al. Prolactin induces MFG-E8 production in macrophages via transcription factor C/EBPbeta-dependent pathway. Apoptosis. 2008, 13, 609–620. [Google Scholar] [CrossRef]
- Sawatani, Y.; Miyamoto, T.; Nagai, S.; Maruya, M.; Imai, J.; Miyamoto, K.; Fujita, N.; Ninomiya, K.; Suzuki, T.; Iwasaki, R.; et al. The role of DC-STAMP in maintenance of immune tolerance through regulation of dendritic cell function. Int. Immunol. 2008, 20, 1259–1268. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | FC | p (Corr) | Gene Symbol | FC | p (Corr) |
|---|---|---|---|---|---|
| Npy | −742.1 | 1.6 × 10−05 | Dgat2 | 19.0 | 1.3 × 10−04 |
| 1700112E06Rik | −9.2 | 1.4 × 10−04 | Nbea | 8.2 | 6.9 × 10−04 |
| Rtn4rl2 | −5.4 | 2.6 × 10−02 | Morn4 | 7.8 | 3.8 × 10−06 |
| Dcstamp | −4.7 | 1.4 × 10−02 | 2410066E13Rik | 7.4 | 1.3 × 10−06 |
| Itgb7 | −4.7 | 2.0 × 10−02 | Fah | 6.7 | 7.2 × 10−08 |
| Gata2 | −4.5 | 1.5 × 10−02 | Hddc3 | 5.9 | 7.1 × 10−07 |
| Wdr54 | −3.8 | 1.4× 10−02 | Wdfy1 | 5.8 | 2.2 × 10−07 |
| Elmod3 | −3.2 | 3.2 × 10−04 | Dkk2 | 4.9 | 2.7 × 10−05 |
| Gm15446 | −2.9 | 5.3 × 10−03 | Cdo1 | 4.4 | 1.9 × 10−04 |
| Gm7799 | −2.9 | 5.0 × 10−02 | Sh3d21 | 4.4 | 6.8 × 10−06 |
| Gpr68 | −2.9 | 4.8 × 10−02 | Slco3a1 | 4.3 | 8.4 × 10−09 |
| Krt80 | −2.9 | 2.5 × 10−02 | AC166773.1 | 3.9 | 4.1 × 10−06 |
| Mfge8 | −2.7 | 2.4 × 10−02 | Gbp10 | 3.6 | 1.7 × 10−03 |
| Pde1c | −2.6 | 7.1 × 10−03 | Gpx3 | 3.5 | 2.6 × 10−04 |
| Galnt9 | −2.5 | 3.5 × 10−02 | Gm13014 | 3.5 | 3.0 × 10−04 |
| Gm12895 | −2.5 | 2.9 × 10−02 | Camk2b | 3.5 | 1.6 × 10−03 |
| Gm14109 | −2.5 | 2.6 × 10−03 | Pde2a | 3.2 | 4.4 × 10−07 |
| Gm9746 | −2.4 | 2.5 × 10−04 | Lrrc9 | 3.2 | 7.3 × 10−04 |
| Kalrn | −2.4 | 4.0 × 10−02 | Itm2a | 3.0 | 2.9 × 10−04 |
| Il1rn | −2.4 | 1.8 × 10−02 | Xkr6 | 2.9 | 7.7 × 10−04 |
| Itpka | −2.3 | 5.4 × 10−05 | Rab4a | 2.7 | 5.3 × 10−07 |
| Dnajb13 | −2.3 | 3.5 × 10−02 | Pyroxd2 | 2.7 | 8.0 × 10−04 |
| Tnip3 | −2.2 | 1.0 × 10−02 | AC123679.1 | 2.6 | 8.2 × 10−06 |
| Gpc1 | −2.2 | 2.0 × 10−02 | Efcab7 | 2.4 | 3.6 × 10−04 |
| RP24-281K23.1 | −2.1 | 7.1 × 10−03 | Prdm5 | 2.3 | 1.7 × 10−04 |
| Gm3788 | −2.1 | 1.0 × 10−02 | Mageh1 | 2.2 | 9.8 × 10−04 |
| Gm9844 | −2.1 | 1.0 × 10−02 | Mlph | 2.2 | 1.5 × 10−03 |
| Tmsb10 | −2.1 | 1.9 × 10−02 | Gm4772 | 2.2 | 3.8 × 10−04 |
| 4931413K12Rik | −2.0 | 1.4 × 10−02 | Nnt | 2.1 | 7.3 × 10−07 |
| D14Ertd449e | −2.0 | 1.8 × 10−03 | Dhcr24 | 2.1 | 5.0 × 10−05 |
| RetSat | −2.0 | 2.9 × 10−04 | Syp | 2.1 | 1.6 × 10−03 |
| RP23-291E6.5 | −2.0 | 2.9 × 10−02 | Klra3 | 2.1 | 1.4 × 10−05 |
| Itgax | −1.9 | 4.0 × 10−02 | B130055M24Rik | 2.0 | 6.3 × 10−04 |
| Ak4 | −1.9 | 3.0 × 10−03 | C1qtnf6 | 2.0 | 6.1 × 10−05 |
| Csf1 | −1.9 | 1.4 × 10−02 | Pdgfc | 2.0 | 7.6 × 10−05 |
| Tiam1 | −1.9 | 3.3 × 10−02 | AL627077.2 | 2.0 | 1.0 × 10−03 |
| Maff | −1.9 | 1.0 × 10−02 | Gm13228 | 2.0 | 9.8 × 10−09 |
| Cd52 | −1.9 | 2.9 × 10−03 | Spr-ps1 | 2.0 | 5.8 × 10−04 |
| Zfp932 | −1.8 | 1.6 × 10−02 | Neo1 | 1.9 | 9.7 × 10−04 |
| Gm11625 | −1.8 | 3.4 × 10−02 | Tmem231 | 1.9 | 4.7 × 10−04 |
| Ide | −1.7 | 7.2 × 10−04 | Fam115a | 1.9 | 6.3 × 10−04 |
| Dfna5 | −1.7 | 1.4 × 10−02 | Usp11 | 1.8 | 9.5 × 10−04 |
| Sema4a | −1.7 | 1.2 × 10−02 | Aldh1l1 | 1.8 | 4.6 × 10−04 |
| Cldn26 | −1.7 | 1.7 × 10−02 | Zcchc14 | 1.8 | 1.2 × 10−03 |
| Lancl2 | −1.7 | 7.1 × 10−04 | Dynlt1b | 1.8 | 6.8 × 10−04 |
| Blm | −1.7 | 4.3 × 10−02 | Dynlt1a | 1.8 | 8.3 × 10−04 |
| Oit3 | −1.7 | 4.0 × 10−02 | Ric3 | 1.7 | 6.4 × 10−04 |
| Cebpb | −1.7 | 3.1 × 10−02 | Slc40a1 | 1.7 | 7.9 × 10−05 |
| Hilpda | −1.6 | 1.3 × 10−02 | Ppp1r9a | 1.7 | 2.6 × 10−04 |
| S100a8 | −1.6 | 2.5 × 10−03 | Pbxip1 | 1.7 | 8.5 × 10−04 |
| Padi4 | −1.6 | 1.9 × 10−02 | Dynlt1-ps1 | 1.6 | 1.5 × 10−04 |
| Ephx1 | −1.6 | 1.3 × 10−02 | Cd59a | 1.6 | 1.2 × 10−05 |
| Gm7665 | −1.5 | 4.5 × 10−02 | Ms4a6b | 1.6 | 4.9 × 10−05 |
| Entpd4 | −1.5 | 2.9 × 10−04 | Sass6 | 1.6 | 1.3 × 10−04 |
| Gm11810 | −1.5 | 4.2 × 10−02 | Plekha8 | 1.6 | 3.9 × 10−04 |
| Napsa | −1.5 | 3.1 × 10−02 | Cadm1 | 1.6 | 1.4 × 10−03 |
| Ifi30 | −1.5 | 1.1 × 10−02 | Nradd | 1.6 | 4.4 × 10−04 |
| Gm12854 | −1.5 | 1.5 × 10−02 | AC113059.1 | 1.5 | 1.1 × 10−03 |
| Gm10108 | −1.5 | 1.9 × 10−02 |
| GO-Term | Description | Count in Gene Set | FDR |
|---|---|---|---|
| GO:0061003 | positive regulation of dendritic spine morphogenesis | 4 of 23 | 0.0107 |
| GO:0036006 | cellular response to macrophage colony-stimulating factor stimulus | 3 of 8 | 0.0126 |
| GO:0045655 | regulation of monocyte differentiation | 3 of 17 | 0.0294 |
| GO:0032502 | developmental process | 40 of 5213 | 0.0294 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarang, Z.; Sághy, T.; Budai, Z.; Ujlaky-Nagy, L.; Bedekovics, J.; Beke, L.; Méhes, G.; Nagy, G.; Rühl, R.; Moise, A.R.; et al. Retinol Saturase Knock-Out Mice are Characterized by Impaired Clearance of Apoptotic Cells and Develop Mild Autoimmunity. Biomolecules 2019, 9, 737. https://doi.org/10.3390/biom9110737
Sarang Z, Sághy T, Budai Z, Ujlaky-Nagy L, Bedekovics J, Beke L, Méhes G, Nagy G, Rühl R, Moise AR, et al. Retinol Saturase Knock-Out Mice are Characterized by Impaired Clearance of Apoptotic Cells and Develop Mild Autoimmunity. Biomolecules. 2019; 9(11):737. https://doi.org/10.3390/biom9110737
Chicago/Turabian StyleSarang, Zsolt, Tibor Sághy, Zsófia Budai, László Ujlaky-Nagy, Judit Bedekovics, Lívia Beke, Gábor Méhes, Gábor Nagy, Ralph Rühl, Alexander R. Moise, and et al. 2019. "Retinol Saturase Knock-Out Mice are Characterized by Impaired Clearance of Apoptotic Cells and Develop Mild Autoimmunity" Biomolecules 9, no. 11: 737. https://doi.org/10.3390/biom9110737