Clove Essential Oil as an Alternative Approach to Control Postharvest Blue Mold Caused by Penicillium italicum in Citrus Fruit



Abstract
:1. Introduction
2. Materials and Methods
2.1. Clove Essential Oil and Pathogens
2.2. In Vivo Antifungal Assay
2.3. Clove Essential Oil Treatment and Sample Collection
2.4. Assay of Hydrogen Peroxide (H2O2) Content
2.5. Determination of Lipid Peroxidation
2.6. Assay of Defense-related Enzymes Activities
2.7. Gene Expression Analysis by Real Time-Quantitative Polymerase Chain Reaction
2.7.1. Total RNA Extraction and cDNA Synthesis
2.7.2. Primer Design
2.7.3. Real Time-Quantitative Polymerase Chain Reaction Analysis
2.8. Statistical Analysis
3. Results and Discussion
3.1. Effect of Clove Essential Oil for Inhibiting Disease Development of Blue Mold Caused by Penicillium italicum
3.2. Effect of Clove Essential Oil Treatment on H2O2 Content in Citrus Fruit
3.3. Effect of Clove Essential Oil Treatment on Malondialdehyde Content in Citrus Fruit
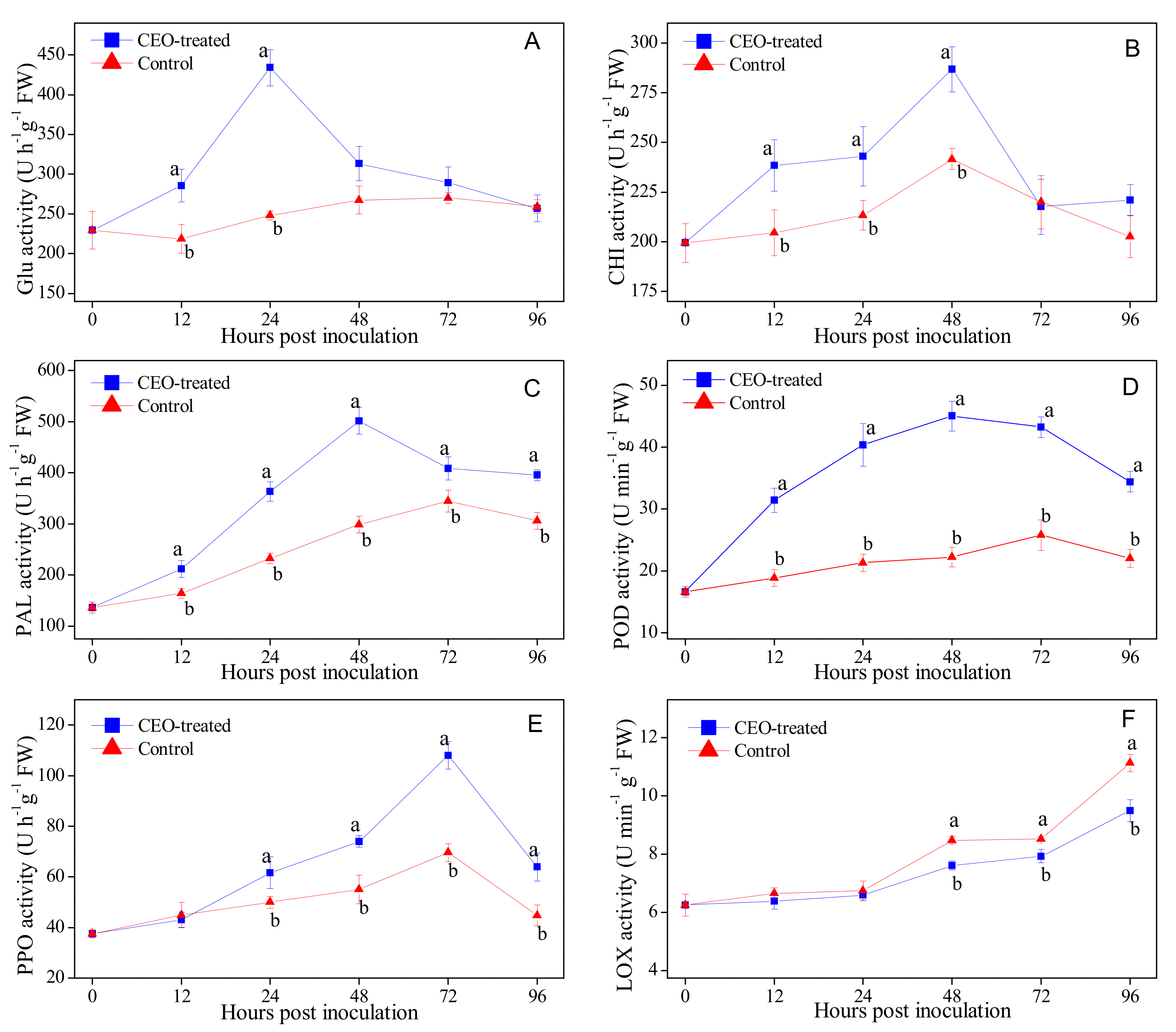
3.4. Effects of Clove Essential Oil Treatment on the Activities of Defense-related Enzymes in Citrus Fruit
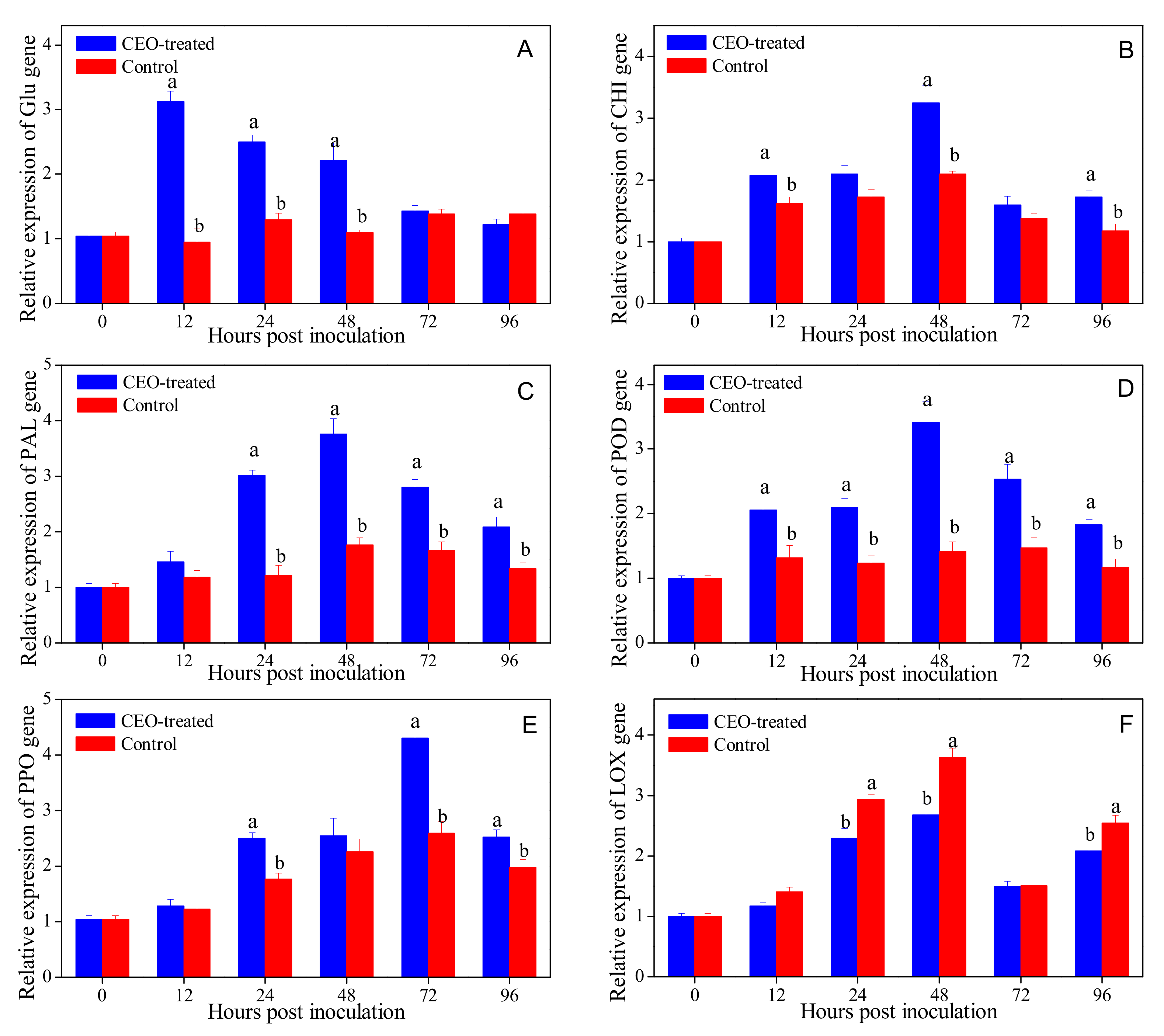
3.5. Effects of Clove Essential Oil Treatment on Defense-related Gene Expression in Citrus Fruit
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El-Otmani, M.; Ait-Oubahou, A.; Zacarías, L. 21—Citrus spp.: Orange, mandarin, tangerine, clementine, grapefruit, pomelo, lemon and lime. In Postharvest Biology and Technology of Tropical and Subtropical Fruits; Yahia, E.M., Ed.; Woodhead Publishing: Cambridge, UK, 2011; pp. 437–516. [Google Scholar] [CrossRef]
- Chen, J.Y.; Shen, Y.T.; Chen, C.Y.; Wan, C.P. Inhibition of key citrus postharvest fungal strains by plant extracts in vitro and in vivo: A Review. Plants 2019, 8, 26. [Google Scholar] [CrossRef]
- Chen, C.Y.; Qi, W.W.; Peng, X.; Chen, J.Y.; Wan, C.P. Inhibitory effect of 7-Demethoxytylophorine on Penicillium italicum and its possible mechanism. Microorganisms 2019, 7, 36. [Google Scholar] [CrossRef] [PubMed]
- Palou, L.; Smilanick, J.L.; Droby, S. Alternatives to conventional fungicides for the control of citrus postharvest green and blue moulds. Stewart Postharvest Rev. 2008, 4, 1–16. [Google Scholar] [CrossRef]
- Sánchez-González, L.; Cháfer, M.; Chiralt, A.; González-Martínez, C. Physical properties of edible chitosan films containing bergamot essential oil and their inhibitory action on Penicillium italicum. Carbohyd. Polym. 2010, 82, 277–283. [Google Scholar] [CrossRef]
- Chafer, M.; Sanchez-Gonzalez, L.; Gonzalez-Martinez, C.; Chiralt, A. Fungal decay and shelf life of oranges coated with chitosan and bergamot, thyme, and tea tree essential oils. J. Food Sci. 2012, 77, 182–187. [Google Scholar] [CrossRef]
- Xing, Y.G.; Xu, Q.L.; Yang, S.M.; Chen, C.K.; Tang, Y.; Sun, S.M.; Zhang, L.; Che, Z.M.; Li, X.H. Preservation mechanism of chitosan-based coating with cinnamon oil for fruits storage based on sensor data. Sensors 2016, 16, 1111–1133. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, T.; Kim, C.; Rahemi, A. Role of Natural volatiles and essential oils in extending shelf life and controlling postharvest microorganisms of small fruits. Microorganisms 2018, 6, 104. [Google Scholar] [CrossRef]
- Chen, C.Y.; Zheng, J.P.; Wan, C.P.; Chen, M.; Chen, J.Y. Effect of carboxymethyl cellulose coating enriched with clove oil on postharvest quality of ‘Xinyu’ mandarin oranges. Fruits 2016, 71, 319–327. [Google Scholar] [CrossRef]
- Omidbeygi, M.; Barzegar, M.; Hamidi, Z.; Naghdibadi, H. Antifungal activity of thyme, summer savory and clove essential oils against Aspergillus flavus in liquid medium and tomato paste. Food Control 2007, 18, 1518–1523. [Google Scholar] [CrossRef]
- He, J.L.; Wu, D.T.; Zhang, Q.; Chen, H.; Li, H.Y.; Han, Q.H.; Lai, X.Y.; Wang, H.; Wu, Y.X.; Yuan, J.G.; et al. Efficacy and mechanism of cinnamon essential oil on inhibition of Colletotrichum acutatum isolated from ‘Hongyang’ kiwifruit. Front. Microbiol. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Goñi, P.; López, P.; Sánchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Mari, M.; Bautista-Baños, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Technol. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential Oils and Antifungal Activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities—Potentials and challenges. Food Control 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Yahyazadeh, M.; Omidbaigi, R.; Zare, R.; Taheri, H. Effect of some essential oils on mycelial growth of Penicillium digitatum Sacc. World J. Microbiol. Biotechnol. 2008, 24, 1445–1450. [Google Scholar] [CrossRef]
- Pérez-Alfonso, C.O.; Martínez-Romero, D.; Zapata, P.J.; Serrano, M.; Valero, D.; Castillo, S. The effects of essential oils carvacrol and thymol on growth of Penicillium digitatum and P. italicum involved in lemon decay. Int. J. Food Microb. 2012, 158, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Ramezanian, A.; Azadi, M.; Mostowfizadeh-Ghalamfarsa, R.; Saharkhiz, M.J. Effect of Zataria multiflora Boiss and Thymus vulgaris L. essential oils on black rot of ‘Washington Navel’ orange fruit. Postharvest Biol. Technol. 2016, 112, 152–158. [Google Scholar] [CrossRef]
- Bill, M.; Sivakumar, D.; Beukes, M.; Korsten, L. Expression of pathogenesis-related (PR) genes in avocados fumigated with thyme oil vapours and control of anthracnose. Food Chem. 2016, 194, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Pheng, T.W.; Mustafa, M.A. Application of lemongrass oil in vapour phase for the effective control of anthracnose of ‘Sekaki’ papaya. J. Appl. Microb. 2015, 118, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Shao, Y.L.; Tang, Y.J.; Zhou, W.W. Antifungal activity of essential oil compounds (Geraniol and Citral) and inhibitory mechanisms on grain pathogens (Aspergillus flavus and Aspergillus ochraceus). Molecules 2018, 23, 2108. [Google Scholar] [CrossRef]
- Banani, H.; Olivieri, L.; Santoro, K.; Garibaldi, A.; Gullino, M.L.; Spadaro, D. Thyme and savory essential oil efficacy and induction of resistance against Botrytis cinerea through priming of defense responses in apple. Foods 2018, 7, 11. [Google Scholar] [CrossRef]
- Aminifard, M.H.; Mohammadi, S. Essential oils to control Botrytis cinerea in vitro and in vivo on plum fruits. J. Sci. Food Agric. 2013, 93, 348–353. [Google Scholar] [CrossRef]
- Chaieb, K.; Hajlaoui, H.; Zmantar, T.; Kahla-Nakbi, A.B.; Rouabhia, M.; Mahdouani, K.; Bakhrouf, A. The chemical composition and biological activity of clove essential oil, Eugenia caryophyllata (Syzigium aromaticum L. Myrtaceae): A short review. Phytother. Res. 2007, 21, 501–506. [Google Scholar] [CrossRef]
- Wang, Y.; Bian, W.; Ren, X.; Song, X.; He, S. Microencapsulatioof clove essential oil improves its antifungal activity against Penicillium digitatum in vitro and green mould on Navel oranges. J. Hort. Sci. Biotechnol. 2018, 93, 159–166. [Google Scholar] [CrossRef]
- Chen, C.Y.; Nie, Z.P.; Wan, C.P.; Chen, J.Y. Preservation of Xinyu tangerines with an edible coating using Ficus hirta Vahl. fruits extract-incorporated chitosan. Biomolecules 2019, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Abeles, F.B.; Bosshart, R.P.; Forrence, L.E.; Habig, W.H. Preparation and purification of glucanase and chitinase from bean leaves. Plant Physiol. 1971, 47, 129–134. [Google Scholar] [CrossRef]
- Surrey, K. Spectrophotometric method for determination of lipoxidase activity. Plant Physiol. 1964, 39, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Ballester, A.R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit–Penicillium digitatum interaction. Postharvest Biol. Technol. 2006, 39, 115–124. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.Y.; Peng, X.; Zeng, R.; Wan, C.P.; Chen, M.; Chen, J.Y. Physiological and biochemical responses in cold-stored citrus fruits to carboxymethyl cellulose coating containing ethanol extract of Impatiens balsamina L. stems. J. Food Process. Pres. 2017, 41. [Google Scholar] [CrossRef]
- Regnier, T.; Combrinck, S.; Veldman, W.; Du Plooy, W. Application of essential oils as multi-target fungicides for the control of Geotrichum citri-aurantii and other postharvest pathogens of citrus. Ind. Crop. Prod. 2014, 61, 151–159. [Google Scholar] [CrossRef]
- Jhalegar, M.J.; Sharma, R.R.; Singh, D. In vitro and in vivo activity of essential oils against major postharvest pathogens of Kinnow (Citrus nobilis × C. deliciosa) mandarin. J. Food Sci. Technol. 2015, 52, 2229–2237. [Google Scholar] [CrossRef]
- Kouassi, K.H.S.; Bajji, M.; Jijakli, H. The control of postharvest blue and green molds of citrus in relation with essential oil–wax formulations, adherence and viscosity. Postharvest Biol. Technol. 2012, 73, 122–128. [Google Scholar] [CrossRef]
- Jongsri, P.; Rojsitthisak, P.; Wangsomboondee, T.; Seraypheap, K. Influence of chitosan coating combined with spermidine on anthracnose disease and qualities of ‘Nam Dok Mai’ mango after harvest. Sci. Hort. 2017, 224, 180–187. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against Penicillium expansum and Botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hort. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Rahmanzadeh Ishkeh, S.; Asghari, M.; Shirzad, H.; Alirezalu, A.; Ghasemi, G. Lemon verbena (Lippia citrodora) essential oil effects on antioxidant capacity and phytochemical content of raspberry (Rubus ulmifolius subsp. sanctus). Sci. Hort. 2019, 248, 297–304. [Google Scholar] [CrossRef]
- Xing, Y.G.; Li, X.H.; Xu, Q.L.; Yun, J.; Lu, Y.Q.; Tang, Y. Effects of chitosan coating enriched with cinnamon oil on qualitative properties of sweet pepper (Capsicum annuum L.). Food Chem. 2011, 124, 1443–1450. [Google Scholar] [CrossRef]
- Stintzi, A.; Heitz, T.; Prasad, V.; Wiedemann-Merdinoglu, S.; Kauffmann, S.; Geoffroy, P.; Legrand, M.; Fritig, B. Plant ‘pathogenesis-related’ proteins and their role in defense against pathogens. Biochimie 1993, 75, 687–706. [Google Scholar] [CrossRef]
- Ballester, A.-R.; Teresa Lafuente, M.; González-Candelas, L. Citrus phenylpropanoids and defense against pathogens. Part II: Gene expression and metabolite accumulation in the response of fruits to Penicillium digitatum infection. Food Chem. 2013, 136, 285–291. [Google Scholar] [CrossRef]
- Bill, M.; Korsten, L.; Remize, F.; Glowacz, M.; Sivakumar, D. Effect of thyme oil vapours exposure on phenylalanine ammonia-lyase (PAL) and lipoxygenase (LOX) genes expression, and control of anthracnose in ‘Hass’ and ‘Ryan’ avocado fruit. Sci. Hort. 2017, 224, 232–237. [Google Scholar] [CrossRef]
- Shao, X.F.; Wang, H.F.; Xu, F.; Cheng, S. Effects and possible mechanisms of tea tree oil vapor treatment on the main disease in postharvest strawberry fruit. Postharvest Biol. Technol. 2013, 77, 94–101. [Google Scholar] [CrossRef]
- Jiwanit, P.; Pitakpornpreecha, T.; Pisuchpen, S.; Leelasuphakul, W. The use of Aloe vera gel coating supplemented with Pichia guilliermondii BCC5389 for enhancement of defense-related gene expression and secondary metabolism in mandarins to prevent postharvest losses from green mold rot. Biol. Control 2018, 117, 43–51. [Google Scholar] [CrossRef]
- Waewthongrak, W.; Leelasuphakul, W.; McCollum, G. Cyclic Lipopeptides from Bacillus subtilis ABS-S14 elicit defense-related gene expression in citrus fruit. PLoS ONE 2014, 9, 11. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | GenBank Accession | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|---|
| β-Glu | AY971953 | ACCTCCGAAGAATCGCTTCCAA | TGTTTCTCATGGCGGGAACA |
| CHI | AF090336 | AATGATGAACGATGCCCTGCCA | CCACTTGATGCTGTCTCCAA |
| PAL | DQ088064.1 | GATTACGGATTCAAGGGTGC | TTGGTGACAGGATTGGCGAG |
| POD | AJ582678.1 | AGCCAGGAGACAATGAACAG | TAGTTTCATGGCCAGTTTGGGC |
| PPO | XM_006468155 | AACTGCTGCCCACCAAAATC | GGTCATAAGCCCCATCACAATA |
| LOX | XM_006483993 | TGACCAAGTCCAAGTTCTGCC | AACGTCTTTTGCGTCCACC |
| β-tubulin | AF052608 | GGTGCAAATCCCACCATGAA | TGGTGTCACTTGCTGCTGCCTGA |
| Index | DPI | Concentrations of CEO (%, v/v) | |||||
|---|---|---|---|---|---|---|---|
| 0.0 | 0.05 | 0.1 | 0.2 | 0.4 | 0.8 | ||
| Lesion diameter | 3 | 23.3 ± 1.33 a | 19.9 ± 0.99 b | 16.4 ± 1.07 c | 13.8 ± 0.92 d | 11.7 ± 1.57 e | 9.8 ± 2.10 f |
| 5 | 35.6 ± 1.71 a | 32.2 ± 1.03 b | 27.6 ± 1.26 d | 22.7 ± 1.16 d | 17.6 ± 1.07 e | 13.9 ± 1.66 f | |
| 7 | 58.9 ± 1.66 a | 56.1 ± 1.20 b | 47.8 ± 1.48 c | 38.4 ± 1.71d | 29.2 ± 1.40 e | 26.8 ± 1.69 e | |
| Disease severity | 3 | 46.9 ± 4.27 a | 42.5 ± 5.40 ab | 33.8 ± 3.23 b | 21.3 ± 2.88 c | 8.8 ± 3.23 d | 5.0 ± 3.53 d |
| 5 | 69.4 ± 5.15 a | 62.5 ± 5.40 a | 58.8 ± 6.61 ab | 48.1 ± 4.27 b | 23.8 ± 4.27 c | 22.5 ± 2.88 c | |
| 7 | 100 ± 0.00 a | 98.8 ± 2.50 a | 95.0 ± 4.56 a | 80.0 ± 4.08 b | 66.3 ± 3.23 c | 63.1 ± 6.25 c | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Cai, N.; Chen, J.; Wan, C. Clove Essential Oil as an Alternative Approach to Control Postharvest Blue Mold Caused by Penicillium italicum in Citrus Fruit. Biomolecules 2019, 9, 197. https://doi.org/10.3390/biom9050197
Chen C, Cai N, Chen J, Wan C. Clove Essential Oil as an Alternative Approach to Control Postharvest Blue Mold Caused by Penicillium italicum in Citrus Fruit. Biomolecules. 2019; 9(5):197. https://doi.org/10.3390/biom9050197
Chicago/Turabian StyleChen, Chuying, Nan Cai, Jinyin Chen, and Chunpeng Wan. 2019. "Clove Essential Oil as an Alternative Approach to Control Postharvest Blue Mold Caused by Penicillium italicum in Citrus Fruit" Biomolecules 9, no. 5: 197. https://doi.org/10.3390/biom9050197