4.2. Flour Characterization
The milling protocol was designed to obtain the minimum particle size since it correlates directly with the extraction efficiency. The value P
50 is the highest diameter (in μm) of the 50% of the particles within the flour, and this value is crucial to determine the efficiency of the oil, protein, and alkaloid extraction in terms of time and yield [
12]. The milling process depends on the moisture of the seed, oil concentration and strength of the seed coat. The SEM analysis showed that
L. angustifolius seeds had the thickest pericarp (
Figure 3b), which was related to the larger particle size of flour of this variety. In the case of proximal composition, the only difference worth to be enhanced is the
L. angustifolius fiber content, higher than
mutabilis variety (around 14% vs. 6%, respectively).
According to Osborne solubility, from the five fractions, the non-extractable was in the last group in
Figure 5 (residue). It is evident that
L. mutabilis, despite its high percentage of protein, about 30% stayed at the end of a high and intensive protocol, so it will not be available for extraction and protein isolates production using conventional techniques. Varieties
albus and
angustifolius seem to be good alternatives for industrial extraction process despite seed’s physical differences and different protein conformation.
Regarding the amino acid profile, the concentration of the sulfur-containing amino acids was low. The recommended dietary allowance for both cysteine and methionine are 58, 27, 22, and 13 mg/kg per day depending on the age group (infants < 6 months old, 2-year old children, 10–12 years old, and adults respectively). The results obtained for
L. albus,
L. angustifolius, and
L. mutabilis were 2.50, 2.58 and 2.20 g/100 g protein, respectively. Taking that into consideration, the infant must consume about 50 g of
L. mutabilis seed (protein: 40% dry weight) in a day to achieve the dietary requirements [
36].
The SDS-PAGE analysis allows the study of the globulins, which are the main fraction of lupin’s seed storage proteins since they comprise the most nutritional, technological, nutraceutical and allergenic potential. Globulins are the salt-soluble storage proteins and can be divided into four (4) groups, depending on its electrophoretic mobility, α-, β-, γ- and δ-conglutins.
There are few reports regarding the electrophoretic pattern of lupine, and this information is very useful, more than just for characterization, to create strategies for protein separation and fractioning. The size and weight of each fraction give an idea of the centrifugal force needed to separate the protein curd from the serum. Also, the isoelectric point is a little bit different among fractions and species and could cause the loss of specific portions crucial to nutritional and functional properties.
Regarding QA concentration in
Lupinus flour and considering a maximal QA daily dose for adults of 0.35 mg/kg body weight, a 75 kg grown man can consume up to 26.25 mg of QA per day [
10]. The lupin’s flour needed to reach the daily dose is 16.4, 65.6 and 1.5 g of
L. albus,
L. angustifolius and
L.
mutabilis, respectively. According to these results,
L. albus and
L. angustifolius are considered sweet species due to the low concentration of QA compared to others like
L. mutabilis which is considered a bitter one.
Lupin flours obtained from the three varieties showed no detectable TIA, this partially agrees with a study conducted by Schoeneberger et al. [
32] where five ecotypes of
L. mutabilis were analyzed, and four (4) did not show any TIA. Only one ecotype presented 1.16 TUI/mL; this was 30 times less than soybean. According to Erbas et al. [
37], the overall lupins inhibitor activity is shallow, and the reported concentration is in the range of 0.1 to 0.2 mg/g.
Fourier Transform Infrared (FTIR) Spectroscopy is an established tool for the characterization of protein secondary structure. Czubinski et al. [
38] described the complete structure of a 7S basic globulin; this is a two subunits monomer present in
L. angustifolius called γ-conglutin (45.4 kDa). The secondary structure of this protein had a rare conformation formed by a 3
10 helix/α-helix complex. In the FTIR deconvolution, a non-common percentage of 3
10 helix was found, possibly indicating the presence of high concentrations of γ-conglutin.
4.3. Effects of the US Treatment in the Protein Isolates Yield and Functional Properties
L. mutabilis and L. angustifolius flours only were used as raw materials for the protein concentration (isolation) procedure to achieve more evidence of the effect of the US treatment that was obtained by preliminary assays. The work was then done with the so-called sweet and bitter lupins (low and high alkaloid percentage).
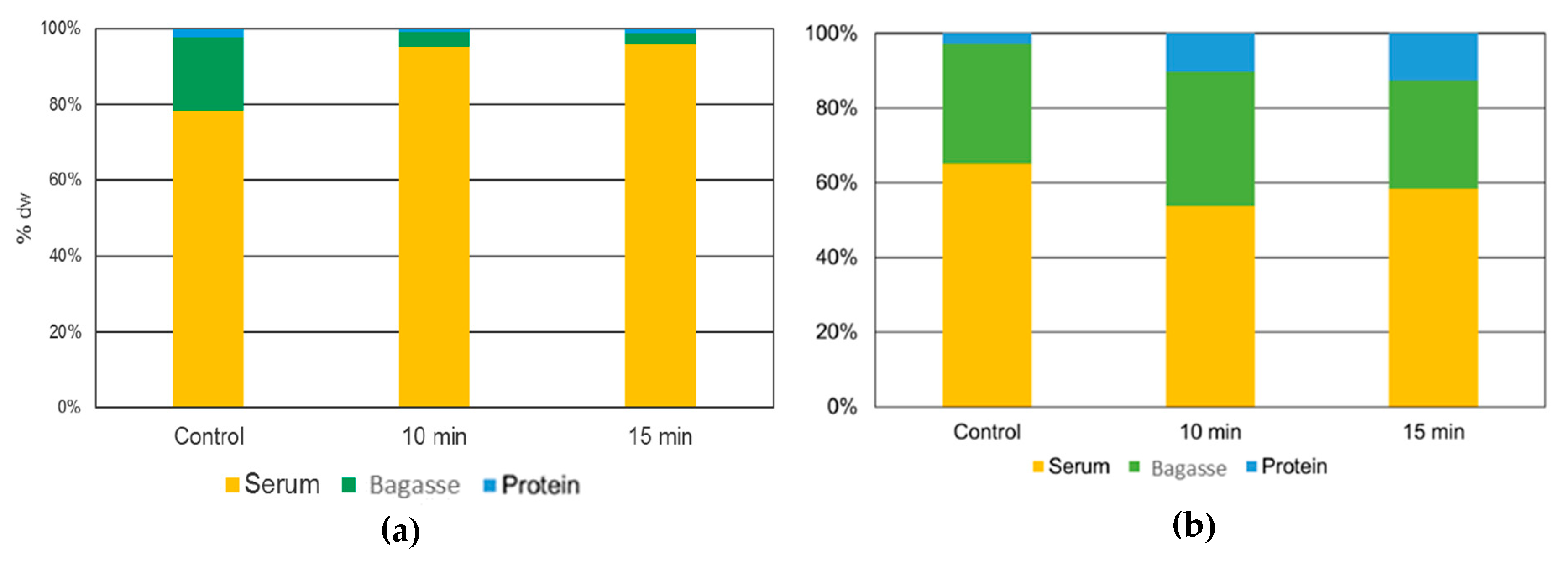
For industrial and commercial proposes, protein yield is significantly essential. The yield of the process is influenced by three principal factors: flour particle size, temperature and protein composition. Protein yield for
L. mutabilis isolates the 10 min US treatment result in the highest protein yield extraction on protein curd. Karki et al. [
39] found a direct correlation between US time and protein concentration on isolates made from soybean flakes. This phenomenon was attributed to the cell rupture caused by the cavitation that releases sugars and protein from the seed tissue. Nevertheless, the longest time proved was 120 s, where a 78 ± 0.40% of protein yield was obtained. For lupin, 10 min was the maximal US time treatment to enhance the protein release. The 15 min treatment was counterproductive in
L. mutabilis isolates (
Figure 7a), dropping the protein yield. This could be due to extreme protein structural damage, which produces aggregates that do not solubilize with rise in pH and possibly end in the bagasse (centrifugation residue), where the 15 min US treatment bagasse fraction was represented.
For
L. angustifolius isolates, the highest protein yield was found in the protein curd treated with 15 min of US. This result showed a different tendency when compared with the
L. mutabilis extraction, where the highest yield was reached with the 10 min US-assisted process. Comparing the protein yield results with Muranyi et al. [
40], who combine several methods like isoelectric point precipitation, salt-induced extraction, alkaline extraction, dilutive precipitation, and their combination over lupin flour the results obtained with US are higher. The most comparable result presented by those authors was a 31.7 ± 0.8% yield using alkaline extraction and a combination of dilutive and isoelectric precipitation. In this research, using US alkaline extraction, more than 50% yield was obtained compared with conventional alkaline extraction in combination with isoelectric precipitation. The lower yield of
L. angustifolius protein fraction (compared to
L. mutabilis) may be explained by the differences in the flour particle size (
Section 3.3.1), the composition and structure of storage proteins.
The functional properties and technological value of the proteins may be influenced negatively by industrial processing. This issue may lead to non-enzymatic browning and secondary structural changes due to heat [
7]. Even with the same amount of protein, two protein isolates can have different water absorption; this could be because of the different globulin fractions between species and cultivars. The reported lupin flour water absorption (
L. albus and
L. mutabilis) were 2.3 and 1.2 g water/g flour DW, respectively. The reason for low WAI value of
L. mutabilis is because of the fat content and probably absence of δ-conglutin, rich in polar amino acids [
12]. The most important effect of the US during the alkaline extraction for
L. mutabilis process is the 15 min treatment, resulting in more than four times higher WAI than the control, probably due to the denaturalization of the protein during the procedure, which causes the exposition of polar amino acids in the medium. The polar amino acids can form hydrogen bonds that enhance the interaction with polar solvents like water [
9]. The water absorbed by flour reported for
L. angustifolius was 2.4 water/g flour DW [
12], this result did not correspond with that depicted in
Figure 8b and could be linked to the high carbohydrate concentration in flour. The lower WAI on
L. angustifolius protein isolates could be linked to composition differences, glycosylation pattern and protein concentration [
41]. The difference in protein concentration for
mutabilis and
angustifolius isolates can indeed affect the water absorption, mainly because in the case of
L. angustifolius, the protein concentration was lower (
Figure 8), indicating the presence of other compounds that affect directly to the protein swelling process [
1]. In other words, to have a higher water absorption, the protein needs to change its structure and make space for the water molecules, but when other compounds as fiber are in the system, this absorption process can be altered.
WSI is a parameter that can be used as an indicator for the degradation of molecular compounds and it depends on the availability of hydrophilic groups that bind water and enhance gel formation of macromolecules. An increase in protein concentration could lead to the diminution of WSI due to the ligation with other soluble molecules [
42]. The protein concentrates had a low concentration of oligosaccharides, which plays an important role in the water solubility of the product [
43]. Other proteins like albumins can increase the WSI due to their affinity to water. Nevertheless, legumes have low concentration of albumins, and protein isolates obtained by isoelectric precipitation do not contain albumin, and the amount of protein-polysaccharide complex is meager. The WSI results of protein isolates of
L. mutabilis compared with the control treatment were low; both US treatments (10 and 15 min) showed half the values of those found in the control treatment. The US treatment could lead to the protein denaturalization and the possible formation of non-soluble structures capable of interacting with other macromolecules present in the protein isolate. Besides, the US treatment can affect the flour carbohydrates too i.e., gelatinizing the starch leading to gel formations and insolubilization of previously soluble fiber. In general, WSI values in
L. angustifolius isolates were the lowest, and no significant differences were found among US treatments.
The NSI describes the ratio between soluble nitrogen and total nitrogen. This parameter is affected by pH, temperature, and ionic strength of the protein solubilization media. For
L. mutabilis isolated, the US treatment with 10 and 15 min showed a reduction of 50% than the NSI in the control treatment due to the thermal treatment caused by US at a pH higher than the isoelectric point. The temperature in the beaker was 63 °C and 77 °C for the 10 min and 15 min treatment, respectively. The protein isolate was not neutralized after freeze-drying; this also can drop the solubility of the proteins as more time is stored in their isoelectric point [
44]. The explanation of this could be the irreversible formation of protein conglomerates and stable conformation with low solubility where the protein is trapped. Isolates of
L. angustifolius showed lower values of
L. mutabilis; however, the samples obtained with 10 min of US application presented the highest values when compared with the other processes. Only one report was found regarding NSI for
L. angustifolius samples, where NSI was about 13.1% for flour and 19.2 to 33.8% for the isolates obtained with different pH levels through alkaline extraction [
44].
The results from
L. mutabilis isolates
in vitro digestibility was significantly higher than the 71.07% and 75.05% reported by Sathe, Desphanade, & Salunkhe [
45] when
L. mutabilis flour and protein concentrate were assessed, respectively. The same authors described the effect of a 30 min boiling step of flour and protein extraction of this variety, yielding an increase of 4.63 and 3.81% in this parameter, respectively, probably due to the protein denaturation, which facilitated the enzymatic degradation. In vitro protein digestibility for
L. angustifolius was reported as 80% and 86.3% for flour and protein isolate at pH 12 respectively [
46]. These results are similar to the ones depicted in
Figure 8b, where the null effect of US was cleared and no significant difference were found for the treatments. Protein Digestibility Corrected Amino Acid Score (PD-CAAS) is the most useful tool to determine the ability of certain proteins to proportionate essential amino acids to the human diet. It reflects the essential amino acid content, true protein digestibility and the bioavailability of the amino acids on the food [
47]. The values of PD-CAAS obtained in
L. mutabilis isolates (
Figure 8a), are very close to those reported but apparently the higher the US time, the lower the PD-CAAS; not because of a low digestibility but due to reduction in cysteine (0.03 g/100 g of protein). In addition, a slight decrease in cysteine could be seen on 10 min US treatment, but this is compensated with the higher digestibility, and that is why this treatment did not present significant difference in the PD-CAAS when compared with the control. For
L. angustifolius isolates, there were no differences among treatments, but there was a slight tendency to increase as the US time arises. This result was associated with a 0.23 g/g crude protein increase in methionine concentration beside a small increase on
in vitro protein digestibility.

 and
and 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}