Zinc and Iron Agronomic Biofortification of Brassicaceae Microgreens
by
 , , and
, , and
Francesco Di Gioia
1,2,* ,
,
Spyridon A. Petropoulos
3,
Monica Ozores-Hampton
2,
Kelly Morgan
2 and
Erin N. Rosskopf
4 1
Department of Plant Science, The Pennsylvania State University, University Park, PA 16802, USA
2
South West Florida Research and Education Center, Institute of Food and Agricultural Sciences, University of Florida, Immokalee, FL 34142, USA
3
Department of Agriculture, Crop Production and Rural Environment, University of Thessaly, 38446 N. Ionia, Magnissia, Greece
4
U.S. Horticultural Research Laboratory, USDA-ARS, 2001 South Rock Rd., Fort Pierce, FL 34945, USA
*
Author to whom correspondence should be addressed.
Agronomy 2019, 9(11), 677; https://doi.org/10.3390/agronomy9110677
Submission received: 27 September 2019
/
Revised: 22 October 2019
/
Accepted: 23 October 2019
/
Published: 25 October 2019
(This article belongs to the Special Issue Biofortification of Crops)
Abstract
:Insufficient or suboptimal dietary intake of iron (Fe) and zinc (Zn) represent a latent health issue affecting a large proportion of the global population, particularly among young children and women living in poor regions at high risk of malnutrition. Agronomic crop biofortification, which consists of increasing the accumulation of target nutrients in edible plant tissues through fertilization or other eliciting factors, has been proposed as a short-term approach to develop functional staple crops and vegetables to address micronutrient deficiency. The aim of the presented study was to evaluate the potential for biofortification of Brassicaceae microgreens through Zn and Fe enrichment. The effect of nutrient solutions supplemented with zinc sulfate (Exp-1; 0, 5, 10, 20 mg L−1) and iron sulfate (Exp-2; 0, 10, 20, 40 mg L−1) was tested on the growth, yield, and mineral concentration of arugula, red cabbage, and red mustard microgreens. Zn and Fe accumulation in all three species increased according to a quadratic model. However, significant interactions were observed between Zn or Fe level and the species examined, suggesting that the response to Zn and Fe enrichment was genotype specific. The application of Zn at 5 and 10 mg L−1 resulted in an increase in Zn concentration compared to the untreated control ranging from 75% to 281%, while solutions enriched with Fe at 10 and 20 mg L−1 increased Fe shoot concentration from 64% in arugula up to 278% in red cabbage. In conclusion, the tested Brassicaceae species grown in soilless systems are good targets to produce high quality Zn and Fe biofortified microgreens through the simple manipulation of nutrient solution composition.
1. Introduction
Iron (Fe) and zinc (Zn) are key trace-elements essential for human health, and their dietary deficiency affects over a quarter of the world population, causing physiological disorders, diseases, and even death, thus constituting a primary global public health challenge [1,2,3,4]. Iron is a crucial constituent of proteins such as hemoglobin and myoglobin devoted to oxygen transportation within the human body, and it is a constituent and activator of a variety of enzymes that are involved in electron transport, oxidation-reduction reactions, and other biological functions [5]. According to the World Health Organization (WHO), iron deficiency anemia is the most ubiquitous health disorder on the global level, and it is closely associated with poor diet, prevalence of cereal-based food, and low consumption of meat, fish, and fruit and vegetables, diets typical of low-income regions in both developed and developing countries [6,7]. Iron is particularly critical for women of childbearing age and young children, and its deficiency increases the risks of maternal mortality. Deficiency can cause stunted growth and irreversible effects on the psychomotor and cognitive function of young children [6,7]. Following Fe in terms of quantitative body requirements, Zn is a structural constituent of proteins or a catalyzing cofactor involved in the activity of several vital enzymes such as RNA polymerase, superoxide dismutase, cellular signaling proteins, thus playing a key role in multiple biological functions [8,9]. Like Fe, Zn deficiency is associated with poor, non-diversified diets characterized by high prevalence of cereal-based food products which have low Zn content and bioavailability [2,4]. Inadequate intake of Zn during pregnancy and in pre-school children can permanently impair physical growth, immunity to disease, reproductive function, and neuro-behavioral development [2,9,10,11]. Similarly to Fe deficiency, the risk of Zn deficiency is significantly higher and widespread in low-income communities in both industrialized and developing countries and primarily affects young children and women [2,6,7,10,11].
Regions with higher risks of Fe and Zn deficiency are generally characterized by soils containing low levels of these nutrients or with properties that limit phytoavailability of trace elements [12]. Moreover, today, the negative effects of climate change on the availability of nutritious food is exacerbating the risks of malnutrition, especially in developing countries [13], and addressing Fe and Zn deficiency in a growing world population could become an emergency. Increasing awareness about the negative impact, especially on women and children, of such an extended and underlying public health issue raised the attention of the scientific community, and a number of studies have highlighted the importance of supplementing the diet of populations that are at risk of Fe and Zn deficiency with food enriched with these microelements [4,6,7,14]. Besides diversifying the diet and trying to improve food selection habits, biofortification of staple crops has been proposed as a primary solution to address Fe and Zn deficiency, as well as for other essential micronutrients [4,15,16]. A number of research projects have focused on crop breeding and genetic engineering, demonstrating that it is possible to fortify and enhance the bioavailability of Fe and Zn in staple crops mainly by: (i) increasing the expression of metal-binding proteins and enzymes; (ii) enhancing the movement and transportation of Fe and Zn from the root to the edible portions; (iii) reducing the content of antinutrients that inhibit Fe and Zn absorption such as phytates, tannins, and dietary fibers; or (iv) increasing the concentration of compounds and factors that promote the absorption of Fe and Zn such as vitamin C, organic and amino acids, and β-carotene [4,14,17,18,19,20,21,22,23]. Nevertheless, this approach has several constraints, and despite the availability of modern breeding and biotechnology tools, the cost and time required to develop genetically biofortified crops make this solution sustainable only in the long run [12]. Especially in the case of engineered crops, due to the high level of regulation and long process of evaluation required, the development and final introduction of a biofortified crop can take years, if not decades, as it requires the approval of plant scientists, policy makers, nutritionists, economists, and last but not least, the acceptance of the entire community [24,25].
As an alternative, agronomic biofortification, which consists of increasing the accumulation of target nutrients in edible plant tissues through fertilization or other eliciting factors has been increasingly proposed in recent years as a simpler, short-term approach to develop functional staple crops and vegetables and address micronutrient deficiency [26,27,28,29,30,31,32]. The exogenous application of cations in plants is not new and has been long used in crop cultivation to overcome problems such as the use of marginal irrigation water and high salinity through the regulation of nutrient uptake and the alleviation of nutrient imbalances caused in plants grown under stress conditions [33,34]. Moreover, according to Chrysargyris et al. [35], foliar application of minerals such as Si and Zn may have a positive effect on growth, development, and mineral content of Lavandula angustifolia (Mill.) plants, especially when grown under salinity stress conditions. However, simply increasing the concentration of micronutrients in edible plant tissues is not enough. Crop biofortification, either genetic or agronomic, is particularly challenging in the case of Fe and Zn, especially in the case of cereals, legumes, and oilseeds, which are characterized by limited micronutrient uptake and high levels of anti-nutrients such as phytates which significantly limit the absorption and bioavailability of Fe and Zn [36,37].
A solution to this problem could be the biofortification of sprouted seeds instead of the regular grain crop as has been already proposed for soybean [38], pea [39], brown rice [29], alfalfa, broccoli, radish, and mung bean [40,41]. For several crops, it has been demonstrated in fact that during the germination process, as the content of phytate in the seed decreases, the bioavailability of Fe and Zn increases [29,42,43]. Yet, agronomic biofortification conducted on a large scale also has some risks related to the variability of micronutrient uptake, which cannot be easily standardized due to the variable interactions between genotypes and environments, as well as to the interactions among nutrients during uptake. Repeated supplemental fertilization of the soil with metals like Fe and Zn can pose environmental and health risks due to their potential leaching into the ground water or the accumulation in the soil or in the plant tissues at excessive levels that can be toxic for the plants and other living organisms, as well as for the consumers [11,17,44]. From this perspective, soilless growing systems combined with customized nutrient solutions of known concentrations could allow a standardization of the process of biofortification and fine control of the product quality, while avoiding or minimizing some of the risks associated with agronomic biofortification of soil grown crops [45,46,47,48]. A number of studies have in fact proven the potential benefits of soilless growing systems in assuring efficient production of high quality and nutritious food crops, especially of leafy vegetables, even under conditions that are limiting or prohibitive for typical soil cultivation, such as in the case of contaminated soils, lack of growing space or availability of non-conventional growing space, scarce and/or low quality water resources, or other limiting conditions that are easily encountered in developing countries and marginal rural or urban areas [49,50,51]. In selecting the method of biofortification and the crops that are most suitable for Fe and Zn biofortification, besides considering the potential physiological limits for the enrichment of a crop with specific micronutrients, it is important to consider the presence of factors that can inhibit or promote micronutrient availability, the presence of antinutritional factors or other risks, the potential for consumer acceptability of the biofortified product, and ideally, it would be good to consider the possibility of developing a method of biofortification simple enough to be transferred to the community in need, allowing them to self-produce biofortified food.
In recent years, microgreens have become increasingly popular as a rich source of vitamins, bioactive compounds, and minerals and have rapidly gained the appellative of “super food” or “functional food” [52,53,54,55,56,57,58,59,60]. Like sprouts, microgreens are a promising crop category for Fe and Zn biofortification [55,57,58]. In fact, using a variety of species, including wild edible plants, even with minimum availability of seeds, microgreens can be easily produced domestically in a relatively short time (7–21 days) within the framework of urban horticulture [54,57,58,61]. The multitude of species suitable for the production of microgreens could guarantee a very rich and complete diet, providing a variety of essential nutrients, while the process of germination with the reduction of phytate as observed in sprouts, and the high content of Fe- and Zn-absorption promoters such as ascorbic acid and β-carotene that characterize microgreens, could assure a high bioavailability of both these trace elements. Although very limited research data is available on microgreen concentrations of Fe and Zn, data reported by Di Gioia et al. [54,58] and Xiao et al. [56] suggest that while there is significant variability among different genotypes, Brassicaceae species could be considered as a good source of both Fe and Zn, and their actual concentration in young plant tissues is highly influenced by the availability of nutrients during the growth period [55,58,59]. Moreover, Brassicaceae microgreens are very popular, relatively inexpensive, easy to germinate and grow, and have great potential health-benefits thanks to their high content of glucosinolates, vitamins, and polyphenols [46,52,53,57,58,59,62]. With such considerations, it is worth exploring the efficacy and potential implications of producing Fe and Zn agronomically biofortified Brassicaceae microgreens. To this purpose, a study was conducted to investigate the effects of increasing levels of Zn and Fe supplied through the nutrient solution on yield components and the mineral profiles of three popular species of Brassicaceae microgreens, namely arugula, red cabbage, and red mustard.
2. Materials and Methods
2.1. Experimental Site, Treatments, and Growing System
Two experiments were conducted during the spring of 2016, in a high tunnel covered with polyethylene film and lateral openings for natural air circulation, at the University of Florida (UF) Institute of Food and Agriculture Science (IFAS) Southwest Florida Research and Education Center (SWFREC) located in Immokalee, FL.
Three Brassicaceae species commonly used to produce microgreens were selected for both experiments: arugula (Eruca sativa (Mill.) Thell.) cv. ‘Astro’, red cabbage (Brassica oleracea L. var. capitata) cv. ‘Red Acre’ and red mustard (Brassica juncea (L.) Czern.) cv. ‘Purple Osaka’. Seeds of all three species were provided by Seedway LLC (Lakeland, FL, USA) and tested for germinability prior to seeding. Seeds were of high quality with a germination rate of 98%, 95%, and 97% at constant 20 °C, while the number of seeds per g was 520 ± 6.4, 180 ± 2.1, and 417 ± 10.8 (mean ± SD, n = 5) for arugula, red cabbage, and red mustard, respectively.
In both experiments, microgreens were grown in a soilless system using grow channels (Cropking Inc., Lodi, OH, USA) 0.25 m wide by 2.8 m long placed on adjustable benches with a slope of approximately 5% to enhance nutrient solution movement. Each channel hosted the three species grown on BioStrate-Felt (Cropking Inc., Lodi, OH, USA) growing mats (23 × 60 cm = 1380 cm2). Plants were fertigated with a half-strength modified Hoagland nutrient solution containing selected levels of zinc (0, 5, 10, and 20 mg L−1) in the first experiment (Exp-1), and selected levels of iron (0, 10, 20, and 40 mg L−1) in the second experiment (Exp-2). The standard nutrient solution was prepared using deionized water containing (mg L−1) 105.1 nitrogen, 15.5 phosphorus, 117.4 potassium, 26.0 magnesium, 92.5 calcium, 34.6 sulfur, 1.20 iron, 0.60 manganese, 0.15 zinc, 0.30 boron, 0.08 copper, and 0.03 molybdenum. Both macro and micronutrients were added to the nutrient solution using simple fertilizer grade salts such as calcium nitrate, potassium nitrate, ammonium nitrate, monopotassium phosphate, potassium sulphate, magnesium sulfate, copper EDTA, manganese sulfate, sodium borate, and sodium molybdate (Helena, Immokalee, FL, USA). In Exp-1, using laboratory grade zinc sulfate heptahydrate (ZnSO4 + 7H2O, Sigma Aldrich, St. Louis, MO, USA), zinc level was adjusted based on the predefined treatment rates (0, 5, 10, and 20 mg L−1), while iron concentration was the same for all Zn enrichment treatments (1.20 mg L−1). In Exp-2, using laboratory grade iron sulfate heptahydrate (FeSO4 + 7H2O, Sigma Aldrich, St. Louis, MO, USA), iron levels were adjusted based on the predefined treatment rates (0, 10, 20, and 40 mg L−1), while zinc concentration was the same for all Fe enrichment treatments (0.15 mg L−1). The final nutrient solution had an average electrical conductivity (EC) of 1.25 dSm−1 and pH 6.2. The nutrient solutions, stored in separate containers for each level of Zn and Fe in Exp-1 and Exp-2, respectively, were pumped to the growing benches and delivered at the upper end of each channel through an irrigation line with five pressure-compensated drippers (each with a delivery rate of 4.0 L h−1) per channel. Particular attention was paid to leveling the growing channels and defining the length of fertigation events to assure adequate and uniform distribution of the nutrient solution along the width and length of each channel at each fertigation event. The nutrient solution delivery was managed with an open cycle system and a 20% minimum drainage fraction in order to ensure that the entire growing mat area was completely wet at every fertigation event, while avoiding nutrient accumulation in the growing mats. The frequency of fertigation events of one or two minutes was adjusted through a timer that activated the individual pumps on each nutrient solution tank. The excess nutrient solution was collected in buckets at the end of each channel to measure the drainage volume and then was discarded. In a real production scenario, a closed cycle system with recirculating nutrient solution would be used for producing Zn and Fe enriched microgreens in order to optimize the use of water and nutrients; however, for the purpose of this research, an open management cycle was used to maintain a constant Zn and Fe concentration during the entire growing cycle in order to record the actual effect of Zn and Fe addition in the nutrient solution on plant nutrient content. Treatments were replicated three times and were arranged according to a split plot design, in which the single channels constituted the main plots and the growing mats of each species constituted the sub-plots.
2.2. Planting Date, Seeding Density, Treatment Differentiation, and Harvest
All three species were sown on 15 February 2016 and on 29 February 2016 for Exp-1 and Exp-2, respectively. Seeding density was defined after preliminary experiments, considering the number of seeds per gram and the germinability of the batch of seeds of each species. The equivalent of 55, 125, and 67 g m−2 of seeds were used in order to obtain a density of 2.9, 2.2, and 2.8 seeds cm−2 for arugula, red cabbage, and red mustard, respectively. After sowing, deionized water was sprayed on top of the seeds using a misting nozzle and growing benches were covered for two days with a white on black polyethylene film allowing seed germination in the dark in both experiments. After complete germination, the third day after sowing (on 18 February 2016 and 3 March 2016, in Exp-1 and Exp-2, respectively), the growth benches were uncovered and fertigation treatments with different levels of Zn or Fe enrichment were applied at each fertigation event until harvest. Microgreens of all three species were harvested 11 days after sowing (DAS) on 26 February 2016 and 11 March 2016 in Exp-1 and Exp-2, respectively. At harvest, all three species had fully expanded and turgid cotyledons with the appearance of the tips of the first true leaves. Harvest was conducted by cutting the shoots a few millimeters above the growing pad surface using clean cutter blades. To avoid border effects, a 2.5 cm strip on each end of the growing mat was excluded from the sampling area. Harvested microgreens were weighed to determine the fresh yield (g m−2) and a pre-marked 10 × 10 cm area in the middle section of each growing area was used for counting the number of germinated shoots to determine the shoot population density (shoots m−2) and the mean shoot fresh weight (mg shoot−1). Dry matter content (g 100 g−1 FW) was determined on fresh samples of approximately 300 g dried until constant weight at 65 °C in a forced-air oven. Dried plant tissue samples were ground using a mill and passed through a 1.0 mm sieve and were used to determine the concentration of P, K, Ca, Mg, Cu, Fe, and Zn.
2.3. Mineral Analysis
Dry and finely ground microgreen tissue samples collected at harvest were analyzed to determine the concentration of P, K, Ca, Mg, Cu, Fe, and Zn using the dry ash combustion digestion method [63,64]. Analyses were performed at the University of Florida in the soil and plant analysis laboratory at the Southwest Research and Education Center, using an inductively coupled plasma atomic emission spectrometry (ICP-AES) system (OES Optima 7000 DV, PerkinElmer, Santa Clara, CA, USA). Dry plant tissue samples of 1.5 g were weighed and dry ashed at 500 °C for 16 h [65]. The ash was equilibrated with 15 mL of 0.5 M HCl at room temperature for 0.5 h. The solution was decanted into 15 mL plastic disposable tubes and placed in a refrigerator at <4 °C [66] until analyses using ICP-AES were performed using calibration and quality assurance procedures as described by Munter et al. [67]. A three-point calibration was performed with standards within the expected range of concentrations. A quality control sample representing a blank, standard, or replicate was included after every 10 unknows in each sample set. Macronutrients and trace-element concentrations determined on dry weight (DW) basis (mg kg−1 of DW) were then transformed and presented on a fresh weight (FW) basis (mg 100 g−1 of FW) using the dry matter percentage previously determined on the same plant tissue samples (see Section 2.2).
2.4. Statistical Analysis
Linear and quadratic regression analyses were used to estimate the yield and nutrient component response of each species to Zn and Fe concentration in the nutrient solution using the regression procedure (PROC REG) of the Statistical Analysis System software (SAS Institute, Cary, NC, USA). Collected data were subject to analysis of variance using the General Linear Models procedure of the Statistical Analysis System software (SAS Institute, Cary, NC, USA). Significant differences between treatments were analyzed using the Student–Newman–Keuls post-hoc multiple comparison procedure at p = 0.05.
3. Results and Discussion
3.1. Microgreens Biometric Response to Zinc (Zn) Enrichment
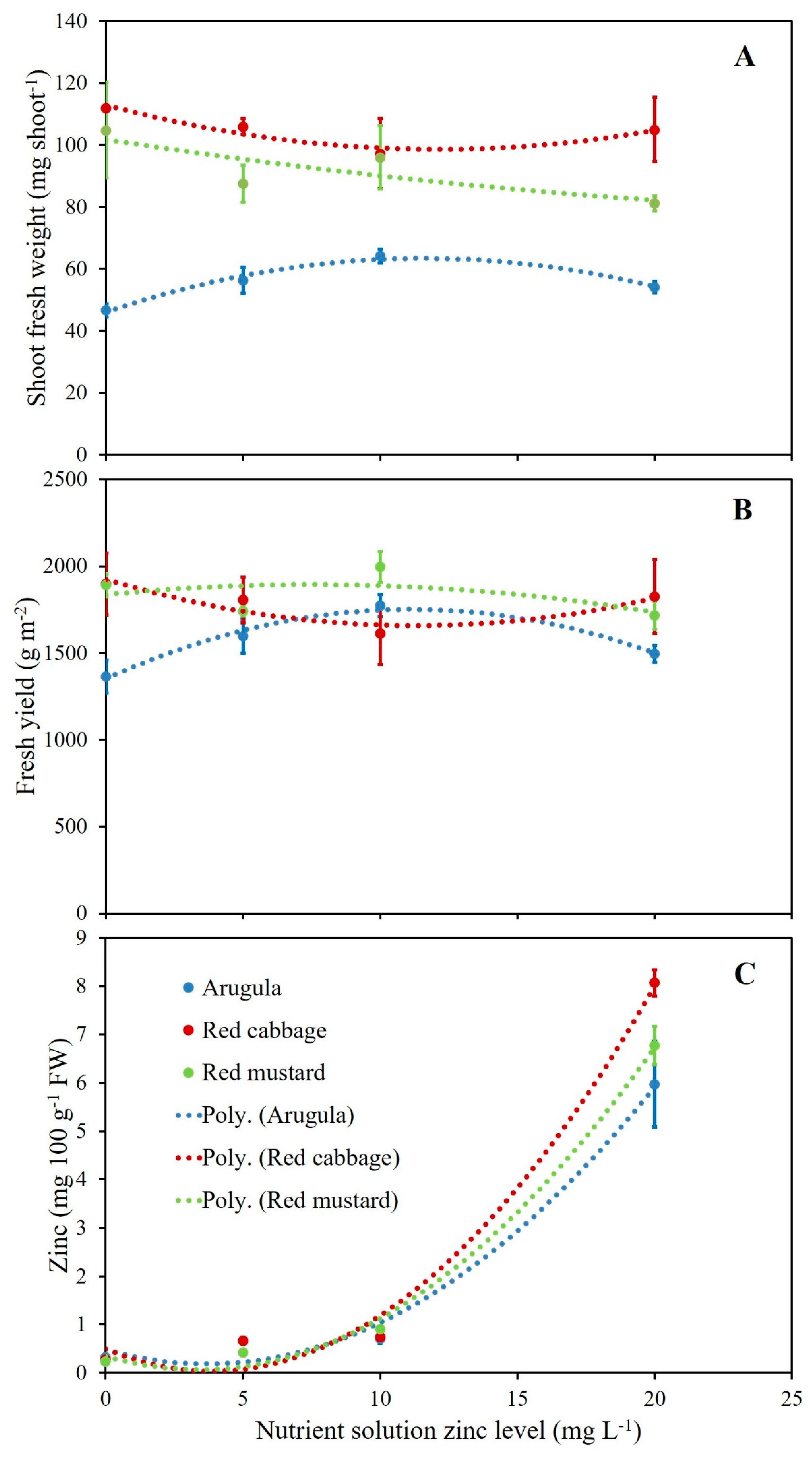
Based on regression analysis shoot density, fresh yield, single-shoot fresh weight, and dry matter content responded to increasing levels of Zn in the nutrient solution in a general quadratic fashion (Figure 1 and Table S1). Nevertheless, while the intercept was always significantly different from zero, slope and quadratic coefficient were significant for fresh yield, shoot fresh weight, and dry matter content only in the case of arugula; for the same parameters R2 values of the quadratic model were 0.63, 0.70, and 0.42, respectively. Apart from arugula and for the three parameters mentioned above, quadratic coefficients were in general very low suggesting a lack of response of red cabbage and red mustard to increasing levels of Zn in the nutrient solution (Figure 1A,B). The low values of R2 observed for both the linear and quadratic model in the case of red cabbage and red mustard for all the biometric parameters examined also suggested a low biometric response to the application of increasing Zn levels (Table S1).
The Zn concentration in the nutrient solution had no effect on the tested parameters (shoot population density, fresh yield, shoot fresh weight, and dry matter content), which were affected only by the species, whereas no significant interaction between the studied factors (Zn levels and species) was observed (Table 1). As expected, the shoot population density was influenced by the seeding density and germination rate of each species and was highest for arugula, which had the highest seeding density and germinability. In contrast, the lowest values for seeding density and germinability were recorded for red cabbage. Fresh yield was lower for arugula compared to the other two species, which may be explained by the smaller seed size and the different plant morphology that characterizes this species compared to the other two brassicas. Consequently, the single shoot mean fresh weight was highest in red cabbage and lowest for arugula, which had the lowest yield and the highest shoot population density, despite the fact that both Brassica species did not differ in terms of fresh yield. Fresh yield and single-shoot fresh weight values in the present study were comparable or higher than those observed in the case of rapini microgreens grown in a similar growing system with the same standard nutrient solution [59]. Dry matter content was higher in red cabbage, followed by arugula, and red mustard, and observed values were in the same range of values observed for rapini microgreens grown in a greenhouse using a similar growing system and the same standard nutrient solution [59].
In a similar study, Wei et al. [29] observed no effect on seed germination in the case of brown rice treated with ZnSO4 solutions up to 150 mg L−1, and Ramezani et al. [68] reported no significant effect of Zn foliar application on the yield of Carum copticum. In the case of soybean sprouts treated with zinc sulfate, testing several application rates from 0 to 100 mg L−1, Zou et al. [38] observed a slight increase in fresh yield with 10 mg L−1 of Zn and a slight yield decrease with 100 mg L−1, while seed germination remained the same or was improved for most of the treated seeds. In contrast, an increase in fresh yield was observed in pea sprouts soaked or sprayed with solutions containing from 10 to 60 mg L−1 of Zn compared to the untreated control [39]. Similarly, Moghimipour et al. [69] and Hassanpouraghdam et al. [70] suggested that Zn application may increase essential oil yield in holy basil (Ocimum sanctum) and affect essential oil composition of basil (Ocimum basilicum). In contrast to the present study, Ghorbanpour et al. [71] reported a positive effect of foliar application of Zn up to 10 mg kg−1 on root and dry weight of sweet basil (Ocimum basilicum L.), a difference that could be due to the application method (foliar vs nutrient solution) as well as to the broader range of the applied Zn levels (0–50 mg kg−1). Moreover, Ahl and Omer [72] suggested that the combined application of Zn and Fe (400 ppm and 200 ppm, respectively) increased plant height and biomass production of coriander (Coriandrum sativum L.). In the study of Hanif et al. [73], who used a similar experimental layout to the present study, the addition of very low Zn concentrations (up to 0.09 mg L−1) in the nutrient solution resulted in the best growth in terms of plant height and fresh yield in sweet basil, although for all the applied Zn concentrations (0.09–0.2 mg L−1), these parameters were higher when compared to the control treatment (0 mg L−1 of Zn). These differences between studies may indicate that the positive effects of Zn application are limited to very low concentrations of Zn and further increases of Zn content in nutrient solution may have limited impact or could have an adverse effect due to toxicity symptoms. However, considering that the aim of the present study was the biofortification of microgreen shoots with adequate amounts of Zn for nutritional purposes, high Zn concentrations were tested in order to define the optimal ratio between high yield and high Zn content in the young shoots.
3.2. Microgreens Nutrient Accumulation Response to Zinc (Zn) Enrichment
Based on regression analysis, it was determined that nutrient accumulation response to increasing levels of Zn in the nutrient solution was, in general, quadratic except for the case of K in arugula and K and Fe in red mustard, which showed a linear response (Table S1). In the case of the Zn accumulation response to Zn level, the quadratic coefficient was significant for all three species and the intercept was not significantly different from zero, and the good fit of the quadratic model was confirmed by high R2 values observed: 0.92, 0.98, 0.98 for arugula, red cabbage, and red mustard, respectively (Figure 1C). A significant quadratic coefficient was observed also in the case of Ca and Mg in arugula and in the case of P and Cu in red mustard. Excluding these cases, the nutrient accumulation response to increasing Zn level in the nutrient solution had low quadratic coefficients and relatively low R2, suggesting that the effect of increasing Zn levels on the accumulation of other nutrients in the shoots of the microgreens tested was weak.
The effect of Zn enrichment and species on mineral content based on the ANOVA is presented in Table 2, where a varied response was observed. For most of the detected minerals, no significant interactions between the tested factors (Zn level and species) were recorded (only in the case of P and Zn content), while the main effects were significant in most cases (except for the effect of Zn enrichment on K and P content). Interestingly, Ca content increased when high Zn levels were applied in the nutrient solution (>5 mg L−1), while in terms of the species effect, red cabbage had the highest Ca content regardless of the Zn level. Similar trends were observed for Mg content although its content did not differ significantly between Zn levels lower than 20 mg L−1. K content was not affected by Zn enrichment since only the species effect was significant, with arugula and red mustard having the highest values. P and Fe content was the highest for red cabbage, whereas the effect of Zn enrichment was significant only for Fe content, where the highest values were recorded for the control treatment (0 mg L−1) without being significantly different from the levels of 5 and 10 mg L−1. In the case of Cu, both factors affected its content in the shoots, with the highest accumulation being detected when 20 mg L−1 of Zn were added in the nutrient solution, as well as in the shoots of arugula and red mustard. Finally, as expected, the highest Zn concentration was recorded at the highest level of Zn application (20 mg L−1) without significant differences between the studied species, although a significant interaction was observed suggesting that the response to Zn was influenced by the species. According to the literature, antagonistic effects between Zn and K, Ca, and Na or Zn and Cu and Fe are commonly detected, since they share the same plasma membrane transporters (P3A-type H-ATPases and P1B-Zn-ATPases, respectively) [74]. However, most of the reports refer to soil and/or foliar applications, which could explain the differences with the results of the current study where Zn was applied directly through the nutrient solution making it more easily available through pH adjustment and lack of Zn adsorption to soil particles [75]. On the other hand, root external cell layers and cell vacuoles may regulate nutrient uptake through influx and efflux transport systems and compartmentalization, which is not possible when nutrients are applied through the foliage [76]. Another reason for the contradictory results would be the short growth cycle of microgreens compared to conventionally cultivated crops which does not allow nutrient imbalances and deficiencies to be expressed within the short time between sowing and harvest, while the absorbed Zn is transported from roots to shoots at these early growth stages [75].
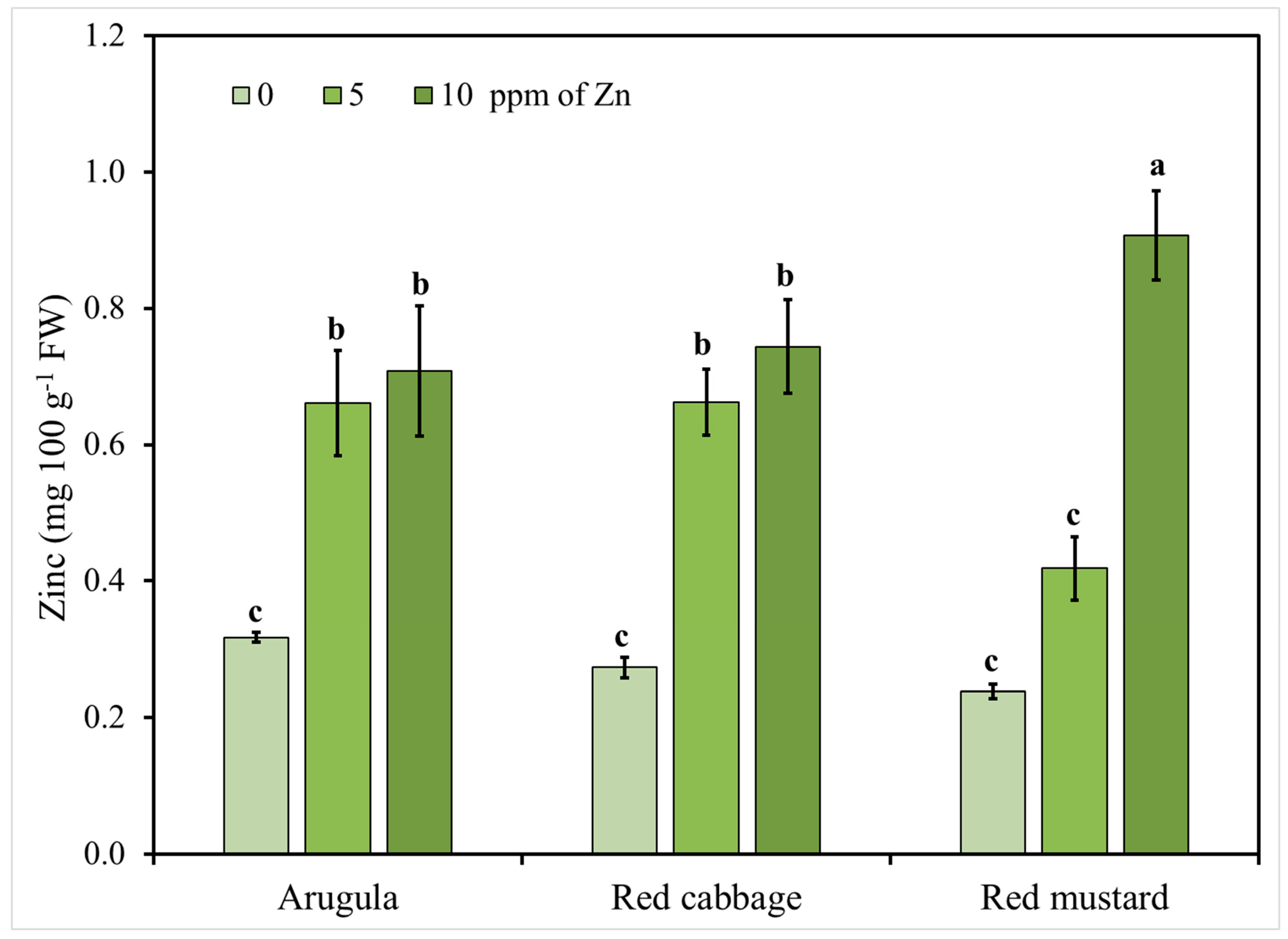
Considering the significant interactions between the tested factors (Zn level and species), Zn accumulation in the shoots of the tested microgreens in relation to Zn application is presented in Figure 2. The results for the highest level of 20 mg L−1 of Zn presented in Table 2 are not presented in this figure since Zn content in shoots reached levels that could be potentially toxic for consumers. It is clearly shown in Figure 2 that increasing the levels of the applied Zn resulted in an increase of Zn content in the shoots of all the tested species, except for the case of red mustard, where Zn content of shoots increased significantly only when more than 5 mg L−1 of Zn were added in the nutrient solution. Moreover, when excluding the highest level of the applied Zn, significant differences were observed between the studied species indicating a genotype dependent response to Zn addition. This finding is of major importance in order to define the safe limits where Zn enrichment is beneficial through the biofortification of the produced microgreens without exceeding toxicity levels for both plants and the consumers.
The recommended daily allowance (RDA) for zinc is estimated to range between 2 and 5 mg for infants and pre-school children and up to 8–11 mg per day for adult females and males, respectively [14,77]. Therefore, it becomes obvious from our results that the consumption of the tested microgreens can cover only a small percentage of daily requirements, especially for adults, and other complementary zinc sources have to be included in the human diet to achieve 100% of the RDA. However, Zn uptake by roots may vary depending on the genotype and the growing conditions [75]; therefore, further studies are required with more species and/or cultivars of the same species under variable growing conditions to define the best practice guidelines for Zn biofortification. Moreover, further research is required to estimate Zn bioavailability in biofortified microgreens.
3.3. Microgreens Biometric Response to Iron (Fe) Enrichment
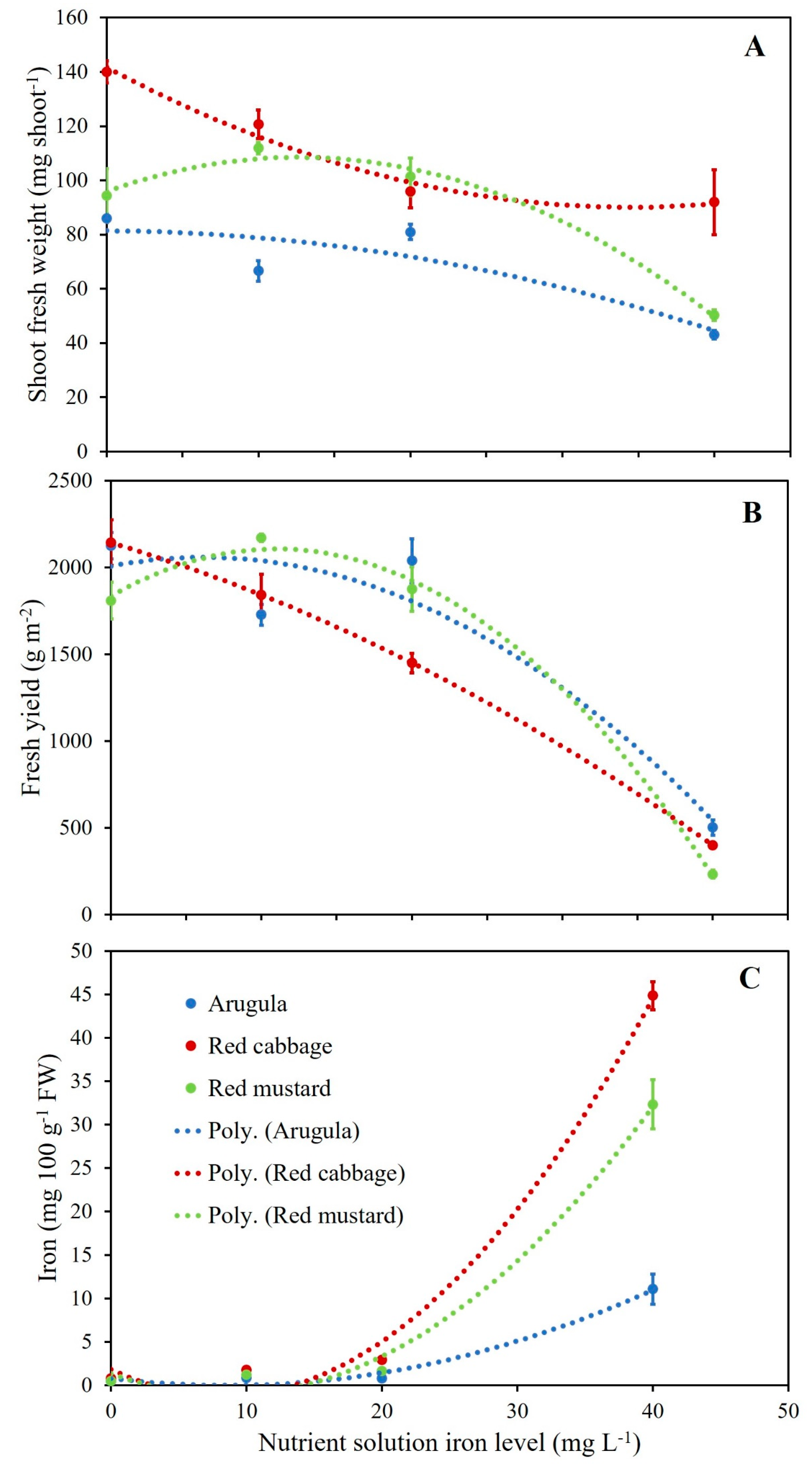
Examining the shoot density, fresh yield, single-shoot fresh weight, and dry matter content response to increasing levels of Fe in the nutrient solution through regression analysis, it was observed that a quadratic model had the best fit for all of the parameters examined in all three species as demonstrated by the relatively high R2 values that ranged between 0.72 and 0.98 (Figure 3 and Table S2). Intercepts were always significantly different from zero and the quadratic coefficient was always significant except for fresh yield and shoot fresh weight in red cabbage and shoot fresh weight and dry matter content in arugula. Quadratic coefficients were all negative except for the shoot fresh weight of red cabbage. Such results suggest significant variations of the biometric parameters in response to the level of Fe in the nutrient solution. Variations in shoot fresh weight and fresh yield in response to Fe level are presented in Figure 3A,B, respectively, and clearly show the phytotoxic effect of Fe at the highest level.
Consistent with the regression analysis, the two-way ANOVA revealed that the Fe addition in the nutrient solution and the species had a significant effect on plant growth parameters (shoot population density, fresh yield, shoot fresh weight, and dry matter content), while a significant interaction between the studied factors (Fe level and species) was also observed (Table 3). In particular, the highest shoot population density was observed in the case of arugula when Fe addition was between 0 and 10 mg L−1. Fresh yield (expressed in g of harvested tissue per m2) was higher when no Fe (in the case of arugula and red cabbage) or 10 mg L−1 (in the case of red mustard) were added. It is important to point out that the addition of high levels of Fe seemed to be toxic for the tested species, since both shoot population density and fresh yield were the lowest for this level for all the tested species. Similarly, in terms of shoot fresh weight (expressed in mg of fresh weight per shoot), Fe enrichment was not beneficial when applied at high concentrations (40 mg L−1), while the highest values were recorded in red cabbage when no Fe was added in the nutrient solution (control treatment). The phytotoxic effect of the highest Fe level (40 mg L−1) was observed visually on all three species immediately after germination due to a lower seed germination and stunting of plants, which would be considered unmarketable. Finally, dry matter (DM) content values confirm the negative effects of the addition of high Fe levels in the nutrient solution, since although Fe is a key element for plant growth and physiological processes, it is also involved in the production of reactive oxygen species and consequently to oxidative stress through the Fenton reaction [78]. Similarly, Przybysz et al. [41] reported a negative impact on fresh yield of broccoli and radish sprouts enriched with Fe at concentrations of 24 and 36 mg L−1, while no negative effects were observed on alfalfa and mung bean sprouts, suggesting that the response to increasing levels of Fe may vary significantly between species. Giordano et al. have also reported negative effects of high levels of Fe (2 mM Fe) in nutrient solution on plant growth and yield parameters of hydroponically grown lettuce [79]. According to Fihlo et al., Fe levels higher than 25 mg L−1 caused toxicity symptoms in hydroponically grown common chicory (Cichorium intybus), although the toxicity threshold may vary among species [80,81].
3.4. Microgreens Nutrient Accumulation Response to Iron (Fe) Enrichment
The nutrient accumulation response to increasing levels of Fe in the nutrient solution was quadratic in most cases except for K in red mustard and Zn for all three species, which showed a linear increase with increasing Fe level as suggested by the positive slope values (Table S2). Excluding the relationship that responded well to a linear model, the quadratic model was a good fit as demonstrated by the significant quadratic coefficient and the relatively high R2 values, with the exception of Ca, K, Mg, and Cu in arugula for which the quadratic coefficient was not significant and R2 values were relatively low. The level of Fe in the nutrient solution significantly impacted the accumulation of other nutrients, with different interactions depending on the species examined. In the case of the Fe accumulation response to Fe level in the nutrient solution, the quadratic coefficient and the slope were significant for all three species except for the slope of arugula, and the intercept was not significantly different from zero for all three species. R2 values ranging from 0.91 to 0.99 confirmed the good fit of the quadratic model for all three species (Figure 3C).
On the basis of the two-way ANOVA, mineral content in the shoots of the studied species was significantly affected by both tested factors (Fe level and species), while significant interactions between these factors were also observed for all the assayed minerals (Table 4). The enrichment of the nutrient solution with the highest concentration of Fe (40 mg L−1) had a positive effect for all the assayed minerals in the case of red cabbage, except for the case of K content where the highest values were recorded in red mustard for the same Fe enrichment level. However, considering that DM content had the highest values at the same Fe enrichment level and the same species (see Table 3), it could be assumed that the observed mineral content could be the result of a concentration effect. As expected, Fe content in the plant tissues increased with increasing application rates and up to 11.07 mg L−1, 44.85 mg L−1, and 32.33 mg L−1 for arugula, red cabbbage, and red mustard, respectively. In the study of Giordano et al., the addition of 2 mM of Fe in the nutrient solution resulted in increased Fe content in the leaves of lettuce, regardless of the genotype [79]. However, in the same study, a negative effect of high Fe levels was observed on Ca, K, and Mg content in lettuce leaves, which was attributed to competition effects among the cations as well as to root injury and oxidative stress [79]. In contrast to the current study, Przybysz et al. did not observe a significant effect of Fe enrichment (up to 36 mg L−1) on Mg, Ca, K, Zn, and Mn content in broccoli sprouts, while a varied effect was recorded on Zn content in radish sprouts [41]. Another implication from the application of high levels of Fe is that it may also increase the accumulation of antinutritional nitrates, as pointed out in the literature for Swiss chard shoots [82]. This could be critical to assess, especially if biofortified microgreens are designed to supplement the diet of infants and pre-school children, because high levels of nitrates could be detrimental for their health causing methemoglobinemia [83,84] and considering that, like other leafy vegetables, microgreens can accumulate relatively high levels of nitrates [58,85,86].
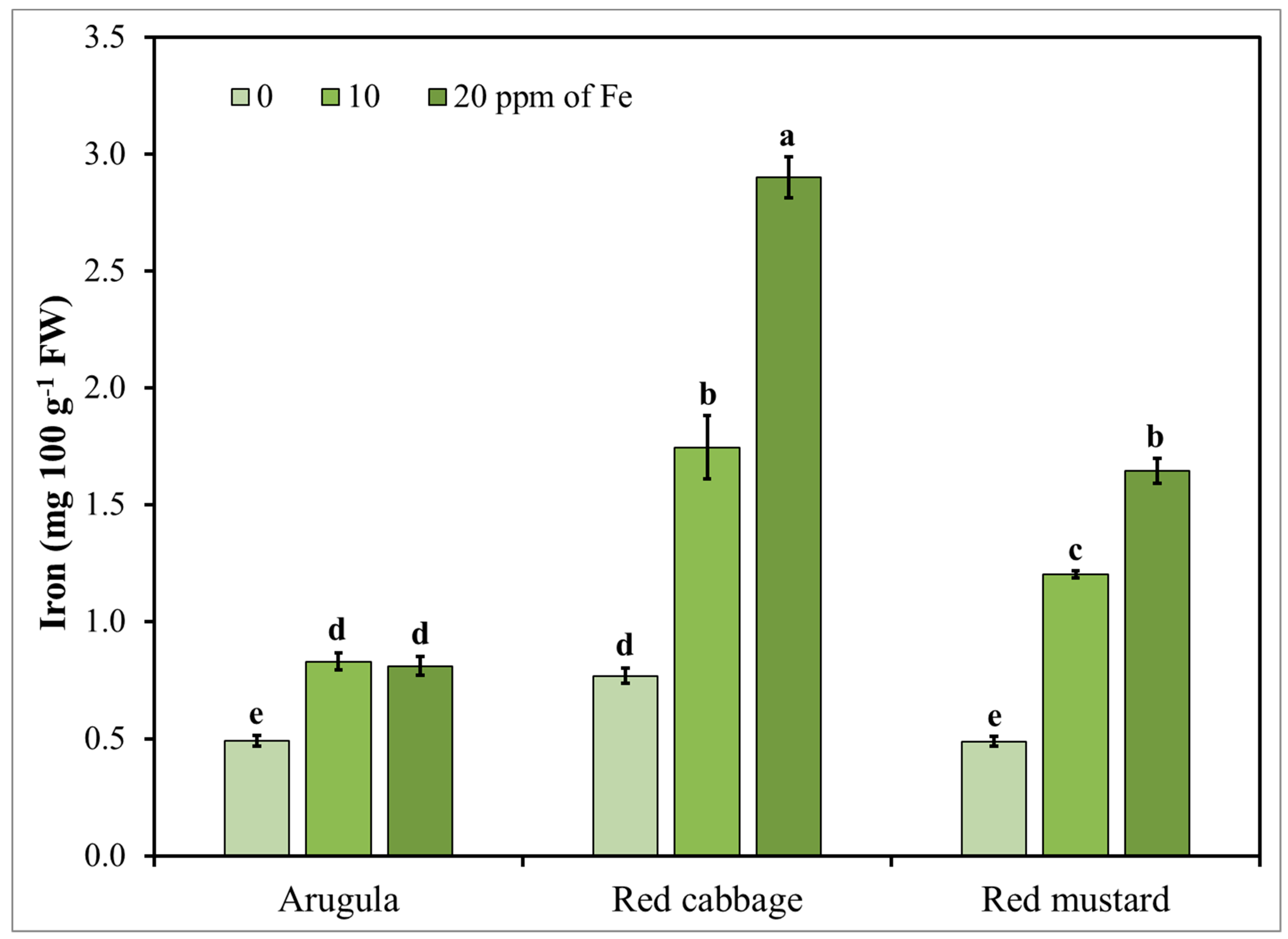
Considering the significant interactions between the tested factors (Fe level and species), Fe accumulation in the shoots of the tested microgreens in relation to Fe application is presented in Figure 4. Similarly to Zn enrichment, the results for the highest level of 40 mg L−1 of Fe are not presented in this figure since Fe content in shoots reached toxicity levels that could be potentially toxic for consumers and were certainly toxic for the plants. As can be seen in Figure 4, increasing the concentration of Fe in the nutrient solution resulted in an increased Fe content in the shoots of all the tested species. Moreover, among the three species, red cabbage was the most sensitive to Fe enrichment where almost a 278% increase in Fe content in shoots was recorded at the level of 20 mg L−1 of Fe compared to the control treatment, while Fe content was about three and two times higher than that of arugula and red mustard, respectively, for the same enrichment level. The levels of Fe increase observed in this study for red mustard and especially red cabbage were considerably higher compared to the increments observed by Przybysz et al., ranging from 50% to 130% at the highest level of Fe (36 mg L−1) in sprouts of broccoli, radish, alfalfa, and mung bean [41]. Such difference, although it could be partially due to a difference in terms of growth cycle length between sprouts and microgreens, further highlights the importance of examining a larger variety of species to identify the ones that are more suitable to Fe biofortification. Differences in Fe content have been also reported between biofortified green and red pigmented lettuce leaves by Giordano et al. and cowpea by Márquez-Quiroz et al., while other factors may also affect the biofortification efficiency including growing conditions and farming practices [28,41,79,80,87]. Considering that the RDA for Fe is estimated at 8–18 mg per day for adults older than 18 years old [14,77], the consumption of even small amounts of these three microgreens could help to cover the total daily requirements and they could be considered as useful dietary supplements, especially in the case of pregnant women where RDA of Fe increases to 27 mg per day. However, considering that Fe uptake mechanisms from plants are closely associated with the uptake processes of other metals potentially toxic to human health, further consideration is needed for Fe biofortification [74]. Yet, as for Zn, further research is required to assess the level of Fe bioavailability in biofortified microgreens.
4. Conclusions
Addressing malnutrition issues such as Fe and Zn deficiency afflicting a large portion of the world population is a big challenge that deserves great attention and dedication in order to enhance the health and life quality of billions of people around the world. While there are no simple solutions to such a complex public health issue, the agronomic biofortification of staple crops and vegetables aimed at artificially enriching their content of essential nutrients has the potential to complement other strategies and contribute to address dietary Fe and Zn deficiency providing a relatively quick short-term solution that may be applied to face nutrition security emergencies. The potential success of this strategy relies on the adequate selection of target crops and the optimization and potential standardization of the biofortification process in order to assure the sustainability of the production process and the obtainment of a biofortified product that is of high quality, safe to eat, effective in delivering adequate levels of micronutrients in order to meet the RDA of the specific element, and easy to introduce as a crop or as a food source to the diet of communities affected by malnutrition and often resident in poor regions of the world. In this study, we propose microgreens as a target crop category for Fe and Zn biofortification. The possibility to self-produce nutrient-dense biofortified microgreens domestically, in a very short time (7–21 days), using only very simple soilless systems with the potential of exploiting a wide variety of species, could contribute to enrich and diversify the diet of malnourished communities in marginal areas of both industrialized and developing countries. By using three Brassicaceae species as test crops, it is demonstrated here that microgreens are a good candidate crop category for Fe and Zn agronomic biofortification. While genotype specific responses were observed for both Fe and Zn enrichment, this study allowed for: (i) identification of levels of Fe and Zn supply through the nutrient solution above which it is possible to observe phytotoxicity and accumulation of Fe and Zn at toxic levels not safe for human consumption; (ii) establishing levels of Fe and Zn supply that may allow for adequate accumulation levels for both micronutrients without decreasing yield and quality of the microgreens; (iii) identification of species that respond better to both Fe and Zn biofortification. Using a simple NFT (nutrient film technique) soilless growing system, supplementing a standard half-strength Hoagland nutrient solution with 5–10 mg L−1 of Zn (ZnSO4) allowed an increase in the concentration in microgreen tissue from a minimum of 75% to a maximum of 281%, the level observed in microgreens grown with a standard nutrient solution without any yield or quality reduction. Similarly, supplementing the standard nutrient solution with 10–20 mg L−1 of Fe allowed an increase in Fe concentration in the microgreens shoots ranging from 64% to 278%, the level observed in the untreated control, with no or only small negative impacts on fresh yield. It is concluded that Brassicaceae microgreens grown in a NFT soilless system are a good candidate crop for Zn and Fe agronomic biofortification through the simple regulation of Fe and Zn concentration in the nutrient solution. Further investigations should be conducted to optimize the biofortification method and (i) explore the suitability of Fe and Zn biofortification of other microgreens species and cultivars; (ii) assess the potential presence of antinutritional (phytates, nitrates, accumulation of toxic metals) or pro-absorption factors (ascorbic acid, β-carotene) in Zn and Fe biofortified microgreens; and (iii) assess the bioavailability and bioaccessibility of both Fe and Zn biofortified microgreens.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4395/9/11/677/s1, Table S1: Parameter estimates, significance, R2, and equation of linear and quadratic response of examined biometric parameters and mineral concentration to nutrient solution zinc concentration in arugula, red cabbage, and red mustard, Table S2: Parameter estimates, significance, R2 and equation of linear and quadratic response of examined biometric parameters and mineral concentration to nutrient solution iron (Fe) concentration in arugula, red cabbage, and red mustard.
Author Contributions
F.D.G. developed the idea, defined the experimental protocol, coordinated the research and carried out the experiment, performed statistical analyses, and prepared the first draft of the manuscript. S.A.P. was significantly involved in data interpretation and manuscript writing and contributed in improving the final version of the manuscript. M.O.-H. provided resources necessary to conduct the experiment and contributed to the definition of the experimental protocol. K.M. provided resources necessary to conduct the experiment and coordinated ICP analyses and data quality and manuscript revision. E.N.R. provided resources necessary to conduct the experiment, contributed to defining procedures for statistical analysis, contributed in improving the final version of the manuscript, and manuscript review.
Funding
This work was supported by the USDA National Institute of Food and Agriculture and Hatch Appropriations under Project #PEN04723 and Accession #1020664.
Acknowledgments
The authors wish to thank Kamal Mahmoud for the assistance in performing the mineral analysis, Joel Mendez, Roger McGill, John Dunckelman, Alisheikh Atta, Gilma Castillo, and Thaisa Cantele for their technical assistance, and Seedway for providing the seeds used in this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McLean, E.; Cogswell, M.; Egli, I.; Wojdyla, D.; De Benoist, B. Worldwide prevalence of anaemia, WHO Vitamin and Mineral Nutrition Information System, 1993-2005. Public Health Nutr. 2009, 12, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Wessells, K.R.; Brown, K.H. Estimating the Global Prevalence of Zinc Deficiency: Results Based on Zinc Availability in National Food Supplies and the Prevalence of Stunting. PLoS ONE 2012, 7, e50568. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.D.D.; Welch, R.M. Food system strategies for preventing micronutrient malnutrition. Food Policy 2013, 42, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Clemens, S. Zn and Fe biofortification: The right chemical environment for human bioavailability. Plant Sci. 2014, 225, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Fraga, C.G. Relevance, essentiality and toxicity of trace elements in human health. Mol. Aspects Med. 2005, 26, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Bharadva, K.; Mishra, S.; Tiwari, S.; Yadav, B.; Deshmukh, U.; Elizabeth, K.E.; Banapurmath, C.R. Prevention of Micronutrient Deficiencies in Young Children: Consensus Statement from Infant and Young Child Feeding Chapter of Indian Academy of Pediatrics. Indian Pediatr. 2019, 56, 577–586. [Google Scholar] [CrossRef]
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; de Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J. Maternal and child undernutrition: Global and regional exposures and health consequences. Lancet 2008, 371, 243–260. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Mackenzie, G.G. Zinc, oxidant-triggered cell signaling, and human health. Mol. Aspects Med. 2005, 26, 245–255. [Google Scholar] [CrossRef]
- Johnson, F.; Giulivi, C. Superoxide dismutases and their impact upon human health. Mol. Aspects Med. 2005, 26, 340–352. [Google Scholar] [CrossRef]
- Hotz, C.; Brown, K.H. Contents International Zinc Nutrition Consultative Group (IZiNCG) Technical Document. Food Nutr. Bull. 2004, 25, S94–S200. [Google Scholar]
- Plum, L.M.; Rink, L.; Hajo, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, I. Enrichment of cereal grains with zinc: Agronomic or genetic biofortification? Plant Soil 2008, 302, 1–17. [Google Scholar] [CrossRef]
- St. Clair, S.B.S.; Lynch, J.P. The opening of Pandora’s Box: Climate change impacts on soil fertility and crop nutrition in developing countries. Plant Soil 2010, 335, 101–115. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortifying crops with essential mineral elements. Trends Plant Sci. 2005, 10, 586–593. [Google Scholar] [CrossRef] [PubMed]
- García-Casal, M.N. Planning and Implementing Food Fortification Programs to Combat Micronutrient Malnutrition: Iron. Food Nutr. Sci. 2014, 5, 880–888. [Google Scholar] [CrossRef] [Green Version]
- Murgia, I.; Arosio, P.; Tarantino, D.; Soave, C. Biofortification for combating “hidden hunger” for iron. Trends Plant Sci. 2012, 17, 47–55. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Clemens, S.; Williams, L.E.; Krämer, U.; Borg, S.; Schjørring, J.K.; Sanders, D. Zinc biofortification of cereals: Problems and solutions. Trends Plant Sci. 2008, 13, 464–473. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Physiological limits to zinc biofortification of edible crops. Front. Plant Sci. 2011, 2, 1–11. [Google Scholar] [CrossRef]
- Dias, D.M.; de Castro Moreira, M.E.; Gomes, M.J.C.; Toledo, R.C.L.; Nutti, M.R.; Sant’Ana, H.M.P.; Martino, H.S.D. Rice and bean targets for biofortification combined with high carotenoid content crops regulate transcriptional mechanisms increasing iron bioavailability. Nutrients 2015, 7, 9683–9696. [Google Scholar] [CrossRef]
- Mukamuhirwa, F.; Tusiime, G.; Mukankusi, M.C. Inheritance of high iron and zinc concentration in selected bean varieties. Euphytica 2015, 205, 349–360. [Google Scholar] [CrossRef]
- Kumar, S.; Palve, A.; Joshi, C.; Srivastava, R.K.; Rukhsar. Crop biofortification for iron (Fe), zinc (Zn) and vitamin A with transgenic approaches. Heliyon 2019, 5, e01914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corguinha, A.P.B.; Carvalho, C.A.; de Souza, G.A.; de Carvalho, T.S.; Vieira, E.A.; Fialho, J.F.; Guilherme, L.R.G. Potential of cassava clones enriched with β-carotene and lycopene for zinc biofortification under different soil Zn conditions. J. Sci. Food Agric. 2019, 99, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.J.; Meenakshi, J.V.; Qaim, M.; Nestel, P.; Sachdev, H.P.S.; Bhutta, Z.A. Potential impacts of iron biofortification in India. Soc. Sci. Med. 2008, 66, 1797–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.B.; Hurrell, R.F. Improving iron, zinc and vitamin A nutrition through plant biotechnology. Curr. Opin. Biotechnol. 2002, 13, 142–145. [Google Scholar] [CrossRef]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Sec. 2017, 12, 49–58. [Google Scholar] [CrossRef]
- Sida-Arreola, J.P.; Sánchez-Chávez, E.; Ávila-Quezada, G.D.; Zamudio-Flores, P.B.; Acosta Muñíz, C.H. Iron biofortification and its impact on antioxidant system, yield and biomass in common bean. Plant, Soil Environ. 2015, 61, 573–576. [Google Scholar]
- Slamet-loedin, I.H.; Johnson-beebout, S.E.; Impa, S.; Tsakirpaloglou, N. Enriching rice with Zn and Fe while minimizing Cd risk. Front. Plant Sci. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Márquez-Quiroz, C.; De-La-cruz-Lázaro, E.; Osorio-Osorio, R.; Sánchez-Chávez, E. Biofortification of cowpea beans with iron: Iron’s influence on mineral content and yield. J. Soil Sci. Plant Nutr. 2015, 15, 839–847. [Google Scholar] [CrossRef]
- Wei, Y.; Shohag, M.J.I.; Wang, Y.; Lu, L.; Wu, C.; Yang, X. Effect of zinc sulfate fortification in germinated brown rice on seed zinc concentration, bioavailability, and seed germination. J. Agric. Food Chem. 2012, 60, 1871–1879. [Google Scholar] [CrossRef]
- Nilsson, J.; Olsson, K.; Engqvist, G.; Ekvall, J.; Olsson, M.; Nyman, M.; Åkesson, B. Variation in the content of glucosinolates, hydroxycinnamic acids, carotenoids, total antioxidant capacity and low-molecular-weight carbohydrates inBrassica vegetables. J. Sci. Food Agric. 2006, 86, 528–538. [Google Scholar] [CrossRef]
- Niyigaba, E.; Twizerimana, A.; Mugenzi, I.; Ngnadong, W.A.; Ye, Y.P.; Wu, B.M.; Hai, J.B. Winter wheat grain quality, zinc and iron concentration affected by a combined foliar spray of zinc and iron fertilizers. Agronomy 2019, 9, 250. [Google Scholar] [CrossRef]
- Ning, P.; Wang, S.; Fei, P.; Zhang, X.; Dong, J.; Shi, J.; Tian, X. Enhancing Zinc Accumulation and Bioavailability in Wheat Grains by Integrated Zinc and Pesticide Application. Agronomy 2019, 9, 530. [Google Scholar] [CrossRef]
- Dordas, C. Application of calcium and magnesium improves yield and essential oil yield of oregano (Origanum vulgare ssp. hirtum). Ital. J. Agron. 2008, 3, 417–418. [Google Scholar]
- Tzortzakis, N.G. Potassium and calcium enrichment alleviate salinity-induced stress in hydroponically grown endives. Hortic. Sci. 2010, 37, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Michailidi, E.; Tzortzakis, N. Physiological and biochemical responses of Lavandula angustifolia to salinity under mineral foliar application. Front. Plant Sci. 2018, 9, 1–23. [Google Scholar] [CrossRef]
- Sandberg, A.-S. Bioavailability of minerals in legumes. Br. J. Nutr. 2002, 88, 281–285. [Google Scholar] [CrossRef] [Green Version]
- Schlemmer, U.; Frølich, W.; Prieto, R.M.; Grases, F. Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis. Mol. Nutr. Food Res. 2009, 53, 330–375. [Google Scholar] [CrossRef]
- Zou, T.; Xu, N.; Hu, G.; Pang, J.; Xu, H. Biofortification of soybean sprouts with zinc and bioaccessibility of zinc in the sprouts. J. Sci. Food Agric. 2014, 94, 3053–3060. [Google Scholar] [CrossRef]
- Lingyun, Y.; Jian, W.; Chenggang, W.; Shan, L.; Shidong, Z. Effect of Zinc Enrichment on Growth and Nutritional Quality in Pea Sprouts. J. Food Nutr. Res. 2016, 4, 100–107. [Google Scholar]
- Park, S.A.; Grusak, M.A.; Oh, M.M. Concentrations of minerals and phenolic compounds in three edible sprout species treated with iron-chelates during imbibition. Hortic. Environ. Biotechnol. 2014, 55, 471–478. [Google Scholar] [CrossRef]
- Przybysz, A.; Wrochna, M.; Małecka-Przybysz, M.; Gawrońska, H.; Gawroński, S.W. Vegetable sprouts enriched with iron: Effects on yield, ROS generation and antioxidative system. Sci. Hortic. 2016, 203, 110–117. [Google Scholar] [CrossRef]
- Afify, A.E.M.M.R.; El-Beltagi, H.S.; El-Salam, S.M.A.; Omran, A.A. Bioavailability of iron, zinc, phytate and phytase activity during soaking and germination of white sorghum varieties. PLoS ONE 2011, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Han, B.Z.; Nout, M.J.R.; Hamer, R.J. Effect of soaking and phytase treatment on phytic acid, calcium, iron and zinc in rice fractions. Food Chem. 2009, 115, 789–794. [Google Scholar] [CrossRef]
- Frei, M.; Tetteh, R.N.; Razafindrazaka, A.L.; Fuh, M.A.; Wu, L.B.; Becker, M. Responses of rice to chronic and acute iron toxicity: Genotypic differences and biofortification aspects. Plant Soil 2016, 408, 149–161. [Google Scholar] [CrossRef]
- Tomasi, N.; Pinton, R.; Dalla Costa, L.; Cortella, G.; Terzano, R.; Mimmo, T.; Scampicchio, M.; Cesco, S. New “solutions” for floating cultivation system of ready-to-eat salad: A review. Trends Food Sci. Technol. 2014, 46, 267–276. [Google Scholar] [CrossRef]
- Di Gioia, F.; Avato, P.; Serio, F.; Argentieri, M.P. Glucosinolate profile of Eruca sativa, Diplotaxis tenuifolia and Diplotaxis erucoides grown in soil and soilless systems. J. Food Compos. Anal. 2018, 69, 197–204. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, Â.; Calhelha, R.C.; Di Gioia, F.; Kolovou, P.; Barros, L.; Ferreira, I.C.F.R. Chemical composition and bioactive properties of Cichorium spinosum L. in relation to nitrate/ammonium nitrogen ratio. J. Sci. Food Agric. 2019. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C. Enhancing quality of fresh vegetables through salinity eustress and biofortification applications facilitated by soilless cultivation. Front. Plant Sci. 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Raviv, M.; Lieth, J.H. Significance of Soilless Culture in Agriculture. In Soilless Culture Theory and Practice; Raviv, M., Lieth, J.H., Eds.; Elsevier: Amsterdam, The Nederlands, 2007; pp. 1–11. [Google Scholar]
- Orsini, F.; Kahane, R.; Nono-Womdim, R.; Gianquinto, G. Urban agriculture in the developing world: A review. Agron. Sustain. Dev. 2013, 33, 695–720. [Google Scholar] [CrossRef]
- Bonasia, A.; Lazzizera, C.; Elia, A.; Conversa, G. Nutritional, biophysical and physiological characteristics of wild rocket genotypes as affected by soilless cultivation system, salinity level of nutrient solution and growing period. Front. Plant Sci. 2017, 8, 15. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Luo, Y.; Wang, Q. Assessment of vitamin and carotenoid concentrations of emerging food products: Edible microgreens. J. Agric. Food Chem. 2012, 60, 7644–7651. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xiao, Z.; Lin, L.-Z.; Lester, G.E.; Wang, Q.; Harnly, J.M.; Chen, P. Profiling polyphenols in five Brassica species microgreens by UHPLC-PDA-ESI/HRMS(n). J. Agric. Food Chem. 2013, 61, 10960–10970. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, F.; Santamaria, P. Microgreens - Novel Fresh and Functional Food to Explore All the Value of Biodiversity; Eco-logica Srl: Bari, Italy, 2015; ISBN 9788890928932. [Google Scholar]
- Ebert, A.W.; Wu, T.H.; Yang, R.Y. Amaranth sprouts and microgreens - A homestead vegetable production option to enhance food and nutrition security in the rural-urban continuum. In Proceedings of the Regional Symposium on Sustaining Small-Scale Vegetable Production and Marketing Systems for Food and Nutrition Security (SEAVEG2014), Bangkok, Thailand, 25–27 February 2014; pp. 233–244. [Google Scholar]
- Xiao, Z.; Codling, E.E.; Luo, Y.; Nou, X.; Lester, G.E.; Wang, Q. Microgreens of Brassicaceae: Mineral composition and content of 30 varieties. J. Food Compos. Anal. 2016, 49, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-scale vegetable production and the rise of microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Di Gioia, F.; Renna, M.; Santamaria, P. Sprouts, Microgreens and “Baby Leaf” Vegetables. In Minimally Processed Refrigerated Fruits and Vegetables. Food Engineering Series; Yildiz, F., Wiley, R., Eds.; Springer: Boston, MA, USA, 2017; pp. 403–432. [Google Scholar]
- Di Gioia, F.; De Bellis, P.; Mininni, C.; Santamaria, P.; Serio, F. Physicochemical, agronomical and microbiological evaluation of alternative growing media for the production of rapini (Brassica rapa L.) microgreens. J. Sci. Food Agric. 2017, 97, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; Pascale, S. De Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Renna, M.; Di Gioia, F.; Leoni, B.; Mininni, C.; Santamaria, P. Culinary Assessment of Self-Produced Microgreens as Basic Ingredients in Sweet and Savory Dishes. J. Culin. Sci. Technol. 2017, 15, 126–142. [Google Scholar] [CrossRef]
- Petropoulos, S.; Di Gioia, F.; Ntatsi, G. Vegetable Organosulfur Compounds and their Health Promoting Effects. Curr. Pharm. Des. 2017, 23, 2850–2875. [Google Scholar] [CrossRef]
- Anderson, D.L.; Henderson, L.J. Comparing Sealed Chamber Digestion with Other Digestion Methods Used for Plant Tissue Analysis. Agron. J. 1988, 80, 549. [Google Scholar] [CrossRef]
- Ozores-Hampton, M.; Di Gioia, F.; Sato, S.; Simonne, E.; Morgan, K. Effects of nitrogen rates on nitrogen, phosphorous, and potassium partitioning, accumulation, and use efficiency in seepage-irrigated fresh market tomatoes. HortScience 2015, 50, 1636–1643. [Google Scholar] [CrossRef]
- Hanlon, E.A.; Gonzalez, J.S.; Bartos, J.M. Mehlich 1 Extractable P, Ca, Mg, Mn, Cu and Zn; University of Florida: Gainesville, FL, USA, 1997. [Google Scholar]
- Plank, C.O. Plant analysis reference procedures for the southern region of the United States. South Coop. Ser. Bull. 1992, 368, 68. [Google Scholar]
- Munter, R.C.; Halverson, T.L.; Anderson, R.D. Quality assurance for plant tissue analysis by ICP-AES. Commun. Soil Sci. Plant Anal. 1984, 15, 1285–1322. [Google Scholar] [CrossRef]
- Ramezani, M.; Seghatoleslami, M.; Mousavi, G.; Sayyari-Zahan, M.H. Effect of salinity and foliar application of iron and zinc on yield and water use efficiency of Ajowan (Carum copticum). Int. J. Agric. Crop Sci. 2012, 4, 421–426. [Google Scholar]
- Moghimipour, Z.; Sourestani, M.M.; Ansari, N.A.; Ramezani, Z. The effect of foliar application of zinc on essential oil content and composition of holy basil [Ocimum sanctum] at first and second harvests. J. Essent. Oil-Bear. Plants 2017, 20, 449–458. [Google Scholar] [CrossRef]
- Hassanpouraghdam, M.; Gohari, G.; Tabatabaei, S.; Dadpour, M.; Shirdel, M. NaCl salinity and Zn foliar application influence essential oil composition of basil (Ocimum basilicum L.). Acta Agric. Slov. 2011, 97, 4–9. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Asgari Lajayer, H.; Hadian, J. Influence of copper and zinc on growth, metal accumulation and chemical composition of essential oils in sweet basil (Ocimum basilicum L.). J. Med. Plants 2016, 15, 132–144. [Google Scholar]
- Ahl, H.A.H.S.; Omer, E.A. Effect of spraying with zinc and / or iron on growth and chemical composition of coriander (Coriandrum sativum L.) harvested at three stages of development. J. Med. Food Plants 2009, 1, 30–46. [Google Scholar]
- Hanif, M.A.; Nawaz, H.; Ayub, M.A.; Tabassum, N.; Kanwal, N.; Rashid, N.; Saleem, M.; Ahmad, M. Evaluation of the effects of Zinc on the chemical composition and biological activity of basil essential oil by using Raman spectroscopy. Ind. Crops Prod. 2017, 96, 91–101. [Google Scholar] [CrossRef]
- Rietra, R.P.J.J.; Heinen, M.; Dimkpa, C.O.; Bindraban, P.S. Effects of nutrient antagonism and synergism on yield and fertilizer use efficiency. Commun. Soil Sci. Plant Anal. 2017, 48, 1895–1920. [Google Scholar] [CrossRef]
- Gupta, N.; Ram, H.; Kumar, B. Mechanism of Zinc absorption in plants: Uptake, transport, translocation and accumulation. Rev. Environ. Sci. Biotechnol. 2016, 15, 89–109. [Google Scholar] [CrossRef]
- Ricachenevsky, F.K.; de Araújo Junior, A.T.; Fett, J.P.; Sperotto, R.A. You shall not pass: Root vacuoles as a symplastic checkpoint for metal translocation to shoots and possible application to grain nutritional quality. Front. Plant Sci. 2018, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; The National Academies Press: Washington, DC, USA, 2001; ISBN 0309072794.
- De Dorlodot, S.; Lutts, S.; Bertin, P. Effects of ferrous iron toxicity on the growth and mineral composition of an interspecific rice. J. Plant Nutr. 2005, 28, 1–20. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Pannico, A.; Kyriacou, M.C.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Iron biofortification of red and green pigmented lettuce in closed soilless cultivation impacts crop performance and modulates mineral and bioactive composition. Agronomy 2019, 9, 290. [Google Scholar] [CrossRef]
- Cecílio Filho, A.B.; Mendoza Cortez, J.W.; de Sordi, D.; Urrestarazu, M. Common Chicory Performance as Influenced by Iron Concentration in the Nutrient Solution. J. Plant Nutr. 2015, 38, 1489–1494. [Google Scholar] [CrossRef]
- Assimakopoulou, A. Effect of iron supply and nitrogen form on growth, nutritional status and ferric reducing activity of spinach in nutrient solution culture. Sci. Hortic. 2006, 110, 21–29. [Google Scholar] [CrossRef]
- Hernández-Castro, E.; Trejo-Téllez, L.I.; Gómez-Merino, F.C.; Rodríguez-Mendoza, M.N.; Sánchez-García, P.; Robledo-Paz, A. Bioaccumulation of iron, selenium, nitrate, and proteins in chard shoots. J. Soil Sci. Plant Nutr. 2015, 15, 694–710. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Santamaria, P. Contribution of Leafy Vegetables to Dietary Nitrate Intake and Regulations. In Nitrate in Leafy Vegetables: Toxicity and Safety Measures; Umar, S., Naser, A., Nafees, A., Eds.; I.K. International Publishing House: New Delhi, India, 2013; pp. 1–16. [Google Scholar]
- Greer, F.R.; Shannon, M. Infant methemoglobinemia: The role of dietary nitrate in food and water. Pediatrics 2005, 116, 784–786. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Ayala, O.; Santamaria, P. Agronomic, physiological and quality response of romaine and red oak-leaf lettuce to nitrogen input. Ital. J. Agron. 2017, 12, 47–58. [Google Scholar] [CrossRef]
- Di Gioia, F.; Gonnella, M.; Buono, V.; Ayala, O.; Cacchiarelli, J.; Santamaria, P. Calcium cyanamide effects on nitrogen use efficiency, yield, nitrates, and dry matter content of lettuce. Agron. J. 2017, 109, 354–362. [Google Scholar] [CrossRef]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef]
Figure 1.
Relationship between single shoot fresh weight (A), fresh yield (B), zinc plant tissue concentration (C) and nutrient solution zinc concentration for arugula, red cabbage, and red mustard. Equations and coefficient of determination (R2) are reported in Table S1.
Figure 1.
Relationship between single shoot fresh weight (A), fresh yield (B), zinc plant tissue concentration (C) and nutrient solution zinc concentration for arugula, red cabbage, and red mustard. Equations and coefficient of determination (R2) are reported in Table S1.

Figure 2.
Arugula, red cabbage, and red mustard microgreens zinc (Zn) concentration in response to nutrient solution Zn level (0, 5, 10 mg L−1). Vertical bars represent the mean ± SE of three replications. Different letters indicate means that are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
Figure 2.
Arugula, red cabbage, and red mustard microgreens zinc (Zn) concentration in response to nutrient solution Zn level (0, 5, 10 mg L−1). Vertical bars represent the mean ± SE of three replications. Different letters indicate means that are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
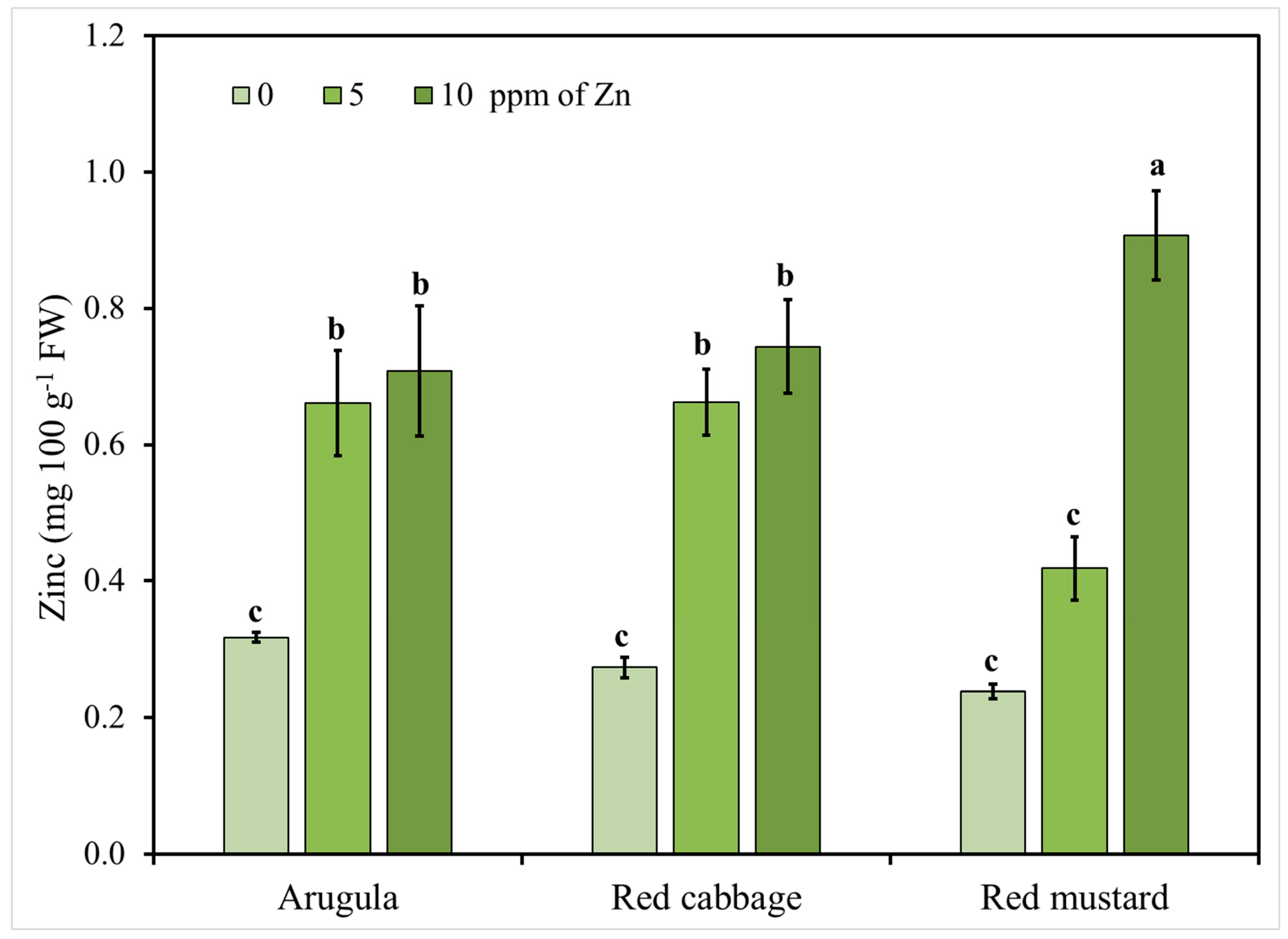
Figure 3.
Relationship between single shoot fresh weight (A), fresh yield (B), iron plant tissue concentration (C) and nutrient solution iron concentration for arugula, red cabbage, and red mustard. Equations and coefficient of determination (R2) are reported in Table S2.
Figure 3.
Relationship between single shoot fresh weight (A), fresh yield (B), iron plant tissue concentration (C) and nutrient solution iron concentration for arugula, red cabbage, and red mustard. Equations and coefficient of determination (R2) are reported in Table S2.

Figure 4.
Arugula, red cabbage, and red mustard microgreens iron (Fe) concentration in response to nutrient solution Fe level (0, 10, 20 mg L−1). Vertical bars represent the mean ± SE of three replications. Different letters indicate means that are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
Figure 4.
Arugula, red cabbage, and red mustard microgreens iron (Fe) concentration in response to nutrient solution Fe level (0, 10, 20 mg L−1). Vertical bars represent the mean ± SE of three replications. Different letters indicate means that are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Nutrient solution zinc (Zn) level effects on shoot population density, fresh yield, mean single shoot fresh weight, and dry matter accumulation of arugula, red cabbage, and red mustard microgreens 1.
Table 1.
Nutrient solution zinc (Zn) level effects on shoot population density, fresh yield, mean single shoot fresh weight, and dry matter accumulation of arugula, red cabbage, and red mustard microgreens 1.
| Treatments | Shoot Population Density | Fresh | Shoot Fresh Weight | Dry |
|---|---|---|---|---|
| Yield | Matter | |||
| (shoot m−2) | (g m−2) | (mg shoot−1) | (g 100 g−1 FW) | |
| Zn (mg L−1) | ||||
| 0 | 22,060 | 1718 | 87.81 | 6.36 |
| 5 | 23,384 | 1716 | 83.29 | 6.42 |
| 10 | 21,363 | 1794 | 85.83 | 6.25 |
| 20 | 23,089 | 1679 | 80.13 | 6.56 |
| Specie (S) | ||||
| Arugula | 25,697 a | 1558 b | 55.32 c | 6.37 b |
| Red cabbage | 19,125 c | 1786 a | 105.08 a | 6.95 a |
| Red mustard | 22,599 b | 1837 a | 92.40 b | 5.87 c |
| p value | ||||
| Zn | 0.31 | 0.57 | 0.80 | 0.45 |
| S | 0.0001 | 0.02 | 0.0001 | 0.0003 |
| Zn × S | 0.71 | 0.19 | 0.13 | 0.45 |
1 Values presented are the average of three replications. Means followed by different letters within each column are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
Table 2.
Nutrient solution zinc (Zn) level effects on the concentration (mg 100 g−1 FW) of calcium (Ca), potassium (K), magnesium (Mg), phosphorous (P), copper (Cu), iron (Fe), and Zn in arugula, red cabbage, and red mustard microgreens 1.
Table 2.
Nutrient solution zinc (Zn) level effects on the concentration (mg 100 g−1 FW) of calcium (Ca), potassium (K), magnesium (Mg), phosphorous (P), copper (Cu), iron (Fe), and Zn in arugula, red cabbage, and red mustard microgreens 1.
| Treatments | Ca | K | Mg | P | Cu | Fe | Zn |
|---|---|---|---|---|---|---|---|
| Zn (mg L−1) | (mg 100 g−1 FW) | ||||||
| 0 | 124.78 b | 361.22 | 43.78 b | 64.89 | 0.05 b | 0.52 a | 0.28 b |
| 5 | 123.11 b | 332.33 | 43.67 b | 64.22 | 0.05 b | 0.44 ab | 0.58 b |
| 10 | 135.00 a | 348.44 | 46.44 ab | 64.56 | 0.05 b | 0.44 ab | 0.79 b |
| 20 | 142.22 a | 355.56 | 49.22 a | 67.44 | 0.07 a | 0.35 b | 6.94 a |
| Species (S) | |||||||
| Arugula | 113.75 b | 374.08 a | 36.75 b | 65.17 b | 0.07 a | 0.42 b | 1.91 |
| Red cabbage | 171.00 a | 301.50 b | 62.25 a | 69.33 a | 0.04 b | 0.53 a | 2.44 |
| Red mustard | 109.08 b | 372.58 a | 38.33 b | 61.33 c | 0.06 a | 0.35 c | 2.08 |
| p value | |||||||
| Zn | 0.01 | 0.72 | 0.01 | 0.38 | 0.02 | 0.01 | 0.0001 |
| S | 0.0001 | 0.003 | 0.0001 | 0.00 | 0.00 | 0.0001 | 0.06 |
| Zn × S | 0.25 | 0.37 | 0.12 | 0.03 | 0.52 | 0.23 | 0.02 |
1 Values presented are the average of three replications. Means followed by different letters within each column are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
Table 3.
Nutrient solution iron (Fe) level effects on shoot population density, fresh yield, mean single shoot fresh weight, and dry matter accumulation of arugula, red cabbage, and red mustard microgreens 1.
Table 3.
Nutrient solution iron (Fe) level effects on shoot population density, fresh yield, mean single shoot fresh weight, and dry matter accumulation of arugula, red cabbage, and red mustard microgreens 1.
| Species (S) | Fe (mg L−1) | Shoot Population Density | Fresh Yield | Shoot Fresh Weight | Dry Matter |
|---|---|---|---|---|---|
| (shoot m−2) | (g m−2) | (mg shoot−1) | (g 100 g−1 FW) | ||
| Arugula | 0 | 29,582 a | 2125 a | 86.0 de | 5.61 e |
| 10 | 28,775 a | 1728 b | 66.6 ef | 6.37 e | |
| 20 | 24,968 b | 2039 ab | 80.9 de | 5.59 e | |
| 40 | 13,520 f | 503 d | 43.1 g | 9.56 c | |
| Red cabbage | 0 | 17,778 def | 2142 a | 140.0 a | 6.69 e |
| 10 | 16,861 ef | 1842 ab | 120.7 b | 7.33 de | |
| 20 | 16,961 ef | 1451 c | 95.8 cd | 8.53 cd | |
| 40 | 5740 g | 400 d | 91.9 cd | 19.32 a | |
| Red mustard | 0 | 22,738 bc | 1810 ab | 94.4 cd | 6.56 e |
| 10 | 22,033 bcd | 2171 a | 112.0 bc | 5.76 e | |
| 20 | 20,113 cde | 1875 ab | 101.3 bcd | 6.13 e | |
| 40 | 4484 g | 234 d | 50.3 fg | 14.10 b | |
| p value | |||||
| Fe | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| S | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| Fe × S | 0.10 | 0.001 | 0.004 | 0.0001 |
1 Values presented are the average of three replications. Means followed by different letters within each column are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
Table 4.
Nutrient solution iron (Fe) level effects on the concentration (mg 100 g−1 FW) of calcium (Ca), potassium (K), magnesium (Mg), phosphorous (P), copper (Cu), Fe, and zinc (Zn) in arugula, red cabbage, and red mustard microgreens 1.
Table 4.
Nutrient solution iron (Fe) level effects on the concentration (mg 100 g−1 FW) of calcium (Ca), potassium (K), magnesium (Mg), phosphorous (P), copper (Cu), Fe, and zinc (Zn) in arugula, red cabbage, and red mustard microgreens 1.
| Species (S) | Fe (mg L−1) | Ca | K | Mg | P | Cu | Fe | Zn |
|---|---|---|---|---|---|---|---|---|
| (mg 100 g−1 FW) | ||||||||
| Arugula | 0 | 119.41 d | 398.16 bc | 38.16 f | 56.80 d | 0.06 c | 0.49 d | 0.28 e |
| 10 | 124.73 d | 344.53 cde | 43.26 ef | 52.37 d | 0.07 c | 0.83 d | 0.41 e | |
| 20 | 119.40 d | 452.23 b | 41.26 ef | 58.21 d | 0.06 c | 0.81 d | 0.56 de | |
| 40 | 150.21 d | 427.07 bc | 56.99 e | 120.81 c | 0.13 bc | 11.07 c | 0.92 c | |
| Red cabbage | 0 | 184.02 c | 297.77 de | 69.52 d | 54.83 d | 0.10 c | 0.77 d | 0.32 e |
| 10 | 197.29 c | 307.91 de | 79.91 cd | 60.41 d | 0.09 c | 1.74 d | 0.55 de | |
| 20 | 207.57 c | 260.71 e | 86.14 bc | 64.07 d | 0.14 bc | 2.90 d | 1.21 b | |
| 40 | 326.83 a | 422.61 bc | 134.51 a | 202.09 a | 0.42 a | 44.85 a | 2.53 a | |
| Red mustard | 0 | 128.81 d | 384.48 bcd | 49.39 ef | 48.10 d | 0.06 c | 0.49 d | 0.25 e |
| 10 | 130.87 d | 475.06 b | 47.55 ef | 52.85 d | 0.07 c | 1.20 d | 0.41 e | |
| 20 | 134.58d | 465.28 b | 51.05 ef | 52.71 d | 0.07 c | 1.64 d | 0.85 cd | |
| 40 | 239.70 b | 566.39 a | 93.68 b | 161.28 b | 0.18 b | 32.33 b | 1.38 b | |
| p value | ||||||||
| Fe | 0.0001 | 0.01 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| S | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
| Fe × S | 0.0001 | 0.01 | 0.0004 | 0.0001 | 0.0001 | 0.0001 | 0.0001 | |
1 Values presented are the average of three replications. Means followed by different letters within each column are significantly different (p < 0.05) using Student-Newman-Keuls multiple range test.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Di Gioia, F.; Petropoulos, S.A.; Ozores-Hampton, M.; Morgan, K.; Rosskopf, E.N. Zinc and Iron Agronomic Biofortification of Brassicaceae Microgreens. Agronomy 2019, 9, 677. https://doi.org/10.3390/agronomy9110677
AMA Style
Di Gioia F, Petropoulos SA, Ozores-Hampton M, Morgan K, Rosskopf EN. Zinc and Iron Agronomic Biofortification of Brassicaceae Microgreens. Agronomy. 2019; 9(11):677. https://doi.org/10.3390/agronomy9110677
Chicago/Turabian StyleDi Gioia, Francesco, Spyridon A. Petropoulos, Monica Ozores-Hampton, Kelly Morgan, and Erin N. Rosskopf. 2019. "Zinc and Iron Agronomic Biofortification of Brassicaceae Microgreens" Agronomy 9, no. 11: 677. https://doi.org/10.3390/agronomy9110677
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.