

The Evolution of Human Social Behavior

Professor Emeritus, University of Oslo, NO-0316 Oslo, Norway
Encyclopedia 2024, 4(1), 430-443; https://doi.org/10.3390/encyclopedia4010029
Submission received: 4 January 2024
/
Revised: 19 February 2024
/
Accepted: 22 February 2024
/
Published: 27 February 2024
(This article belongs to the Section Behavioral Sciences)
{kind=link}
Abstract
:Social life is a key feature in humans; without it, language, science, and technology would not have appeared. The inclination to engage with others is also a main source of pleasure and pain and as such a key factor for quality of life. In this paper, I shall present current knowledge on the evolutionary trajectory leading to the four main types of relations: parent–child, pair-bonding, kinship, and social life (bonding between non-kin for purposes other than breeding). These relationships are not unique to humans; they have evolved independently multiple times across the animal kingdom. In our lineage, the origins of parent–child bonding may be traced back to the early amniotes some 320 Mya (million years ago). Pair-bonding and social life most likely evolved recently. Understanding how these affiliations are rooted in the brain, particularly the role of feelings, provides valuable insights that can help us improve society.
1. Introduction
Social behavior is arguably the most important human feature to understand. An evolutionary perspective offers insight that supplements the social sciences. Dobzhansky’s famous quote “Nothing in biology makes sense except in the light of evolution” [1] is therefore relevant.
Affiliations between individuals, collectively referred to as social behavior, have evolved independently in a wide range of animal lineages. The presence of affiliations is a question of balancing the benefits of solitary living against the compensations offered by associating with others. The various forms, such as parent–child, parental bonding, kinship, and larger constellations (referred to as social life), offer distinct advantages and disadvantages [2]. The main evolutionary advantage of the former two is reasonably obvious—to ensure necessary care for offspring—while social life can serve various functions. It offers, for example, increased vigilance and protection against predators, help in laying down prey, control of territory, and the construction of living quarters as in ants.
I shall focus on our branch of the evolutionary tree. I wish to present current knowledge as to the evolutionary rationale and history behind human affiliations. The neurobiology responsible will be covered briefly.
Affiliations vary from weak to strong, whether one considers differences between species or individuals. In the case of social life, bird colonies typically represent weak bonds with limited impact on behavior. In contrast, the bonds in eusocial animals such as ants and naked mole-rats control much of the observed behavior. There are also considerable variations as to what extent the neurobiology responsible is hard-wired or designed to be molded by experience. In social insects, it is presumably reasonably hard-wired, although even here the behavior is modulated by learning [3]. In the human brain, social behavior is soft-wired, as it is meant to develop and adapt throughout life.
How we relate to each other is arguably the most important factor in creating a society where the inhabitants flourish. If we understand the innate tendencies behind our social behavior and how these tendencies can be molded, we stand a better chance in this endeavor. There are several examples of a similar perspective in the literature [2,4,5,6], yet I believe the present text offers novel information and insight.
Feelings are an important factor in the human brain, not the least in connection with social behavior. In the following section, I discuss their role. In Section 3, I look at the evolution and neurobiology of various forms of affiliations. Section 4 deals with how our innate tendencies play out today. In the final section, I comment on the prospects for improving society.
2. The Role of Feelings
Social behavior requires a neurological strategy to manifest. In our lineage, feelings evolved as a key strategy used to motivate behavior. They were probably introduced in the early amniotes [7] to help the animal evaluate behavioral options in complex situations [8]. Amniotes include reptiles, birds, and mammals; their shared ancestors date back to some 320 Mya. As a behavioral strategy, feelings are soft-wired; they are designed to motivate rather than to drive actions, and the response is meant to be molded by experience.
Motivational neurobiology was probably present before the evolution of feelings and may have served as a neurological scaffold [8]. Feelings gradually gained in importance, but we have retained more instinctive behavioral propensities. Recently, we have evolved a cognitive capacity that can be used to overrule suggestions based on feelings, but they still have a considerable impact. It is somewhat difficult to differentiate between feelings and other motivators as the former can be acted on even when one is unaware of actually feeling anything.
I use the term feeling for any experience with a positive or negative “flavor”. Positive feelings are generally meant to direct behavior toward what is good for the genes in terms of survival and procreation, while negative feelings are there to help us avoid negative outcomes. For example, the acquisition of food or a mate is associated with pleasure, while freezing or being abandoned is painful. One way of categorizing feelings is to divide them into sensations and emotions depending on the source of the experience. Sensations are associated with input from sensory organs, while emotions typically involve interpersonal relations.
Feelings represent a potent way of pushing behavior in a (genetically) desired direction, but this whim of evolution had another unprecedented twist. Rather than just being a reproductive machinery for the genes, the individual can be happy or miserable. The capacity for pleasure and pain introduces a possible purpose for both the individual and society. Rather than measuring success in terms of the number of offspring, happiness can be formulated as what ought to be desired [9]. This idea has become widely popular as witnessed by efforts to measure happiness and direct policy toward improving the score, exemplified by the World Happiness Report [10] and the Happy Planet Index [11]. The ensuing discussion of affiliative behavior reflects the notion that society should aim to enhance the happiness of its inhabitants.
The brain can be described as a collection of functions, or modules, added and modified by the process of evolution since the first nervous systems evolved some 600 Mya [12,13]. Three modules are responsible for our capacity to feel pleasure and pain [14]: Two of these modules offer positive feelings in the form of rewards or pleasure: the seeking (or motivating) module is meant to direct behavior toward finding opportunities, while the liking (or consuming) module is there to ensure that the opportunities are utilized. The difference signifies two distinct types of situations where an urge to act is called for. For example, if you walk past a bakery, the smell activates the seeking module to make you enter; the taste activates the liking module to make sure you eat the cake. The third module represents negative feelings and is referred to as pain. Other brain modules, such as those processing sensory information, can activate either of these three mood modules. The level of happiness can be construed as the net output of the mood modules, that is, the pleasure minus the pain [9].
The neurobiology of the mood modules is partly characterized, each having a particular correlate in the brain [15,16]. The data suggest that roughly the same nerve circuits are involved regardless of the cause of the pleasure or the pain. Various parts of the limbic system (the amygdala, nucleus accumbens, thalamus, and hypothalamus) are assumed to serve a role in all three modules.
Feelings permeate human cognition. They are probably more or less constantly active and add a layer of good or bad to both thoughts and experiences. The layer is there to help sway behavior in particular directions. The positive or negative component can be too subtle to be consciously recognized, yet capable of guiding behavior [17]. Feelings act as invisible threads that help weave the fabric of our social interactions.
Although the evolutionary rationale is to benefit the genes, the actual behavior may not fit that description. For one, there is an element of learning in what elicits feelings. Even animals in the wild can learn to enjoy masturbation and end up spending more time on the activity than what serves their genes [18]. Two, the present environment offers pleasurable opportunities that evolution has not had time to adapt to. For example, heroin is a potent stimulator, and sex offers ample rewards even in the presence of contraceptives.
The intensity of feelings presumably depends primarily on two factors (not considering personal and situational variation). One is the importance of the behavior in question for the genes, and the other is at what time in our evolutionary history the behavioral propensity was added to the brain. It seems likely that more recent functions to a larger extent rely on feelings. This statement reflects the idea that feelings gradually took over as a strategy for behavioral decisions but also that the evolution of cognitive capacity required stronger incentives to ensure behavior in line with genetic obligations.
3. Affiliations
3.1. Parent–Child
The most widespread affiliation to occur in nature is that of parent–child. While in many species the offspring must fend for itself, it is reasonably common to introduce caring behavior. More often the mother is the provider, but in some species, including certain titi monkeys [19], the father is the main caregiver.
Parental care (after the eggs hatch) is rare in fish, amphibians, and reptiles, while universally present in birds and mammals [20]. The trait may have been present in the ancestral amniotes, or it may have evolved independently at a later stage. Lactation is a certain sign of maternal care. We do not know when lactating evolved in our lineage, but the trait was most likely present in the shared ancestors of present mammals. The monotremes split with marsupials and eutherian mammals some 170 Mya [21], suggesting that maternal bonding was introduced before that date. If nascent feelings first appeared some 320 Mya [7], the bonding should involve positive emotions. Parental care in mammals (and birds) is generally more demanding in terms of the time and resources used compared to that of lower vertebrates, suggesting that strong incentives would be required.
The term love implies positive feelings associated with affiliations; thus, the first love, evolutionary speaking, was that between the mother and her offspring. The bond has evolved to be one of the strongest behavioral forces in the mammalian lineage.
There are considerable data as to the neurobiology of love and attachment [22,23,24]. Briefly, the neurotransmitters/hormones oxytocin and vasopressin play an important role. Dopaminergic circuits are also involved, but this may be due more to their role in the associated rewards rather than the actual sense of attachment. The ventral tegmental area of the midbrain is presumably active, as are the nucleus accumbens and ventral pallidum of the limbic system. Love dampens the activity in circuits associated with fear and negative emotions, possibly because attachment requires the individual to subdue the natural dread of other individuals.
3.2. Pair-Bonding
Birds and mammals reproduce sexually, meaning there is a mother and a father, but sexual reproduction does not require parental bonding. Less than 10% of mammalian species display this form of affiliation [25]; in the remaining species, the act of copulation is the only prominent shared behavior. The act does not require any distinct attachment, as it is cared for by the sexual drive. In contrast, some 90% of bird species form couples [26].
The question of whether the father should bond with the mother depends on what serves the genes best. If the required parental care is too demanding to be handled by one individual, evolution moves toward bonding; on the other hand, if resources are scarce so that two adults are unlikely to find sustenance if staying together, bonding is disfavored. The male has generally more to gain by solitary life, as he then stands a better chance of impregnating other females. In mammals, the female invests more in the early stages of the creation of progeny and is therefore more inclined to continue her engagement in the absence of a male. In birds, the balance leans in favor of couple formation due to the extensive care required for eggs and chicks.
In pair-bonding mammals, the relationships rarely last for a lifetime, and the degree of sexual fidelity is relatively low compared to birds [25,27]. The penchant for pair-bonding in humans probably reflects this observation, as copulations outside of marriage are common in most cultures [28].
Marriage can be defined as a culturally recognized union for the (biological) purpose of raising offspring. The union may involve more than two individuals, but for simplicity, I refer to it as pair-bonding. The potency of feelings such as falling in love and romantic love reflects our innate inclination to bond with the opposite sex, as does the observation that almost all cultures recognize some form of marriage [28]. One notable exception is the Mosuo of Southern China [29]. The exception testifies to the flexibility of the human brain; most innate behavioral tendencies can be overturned given the right environment.
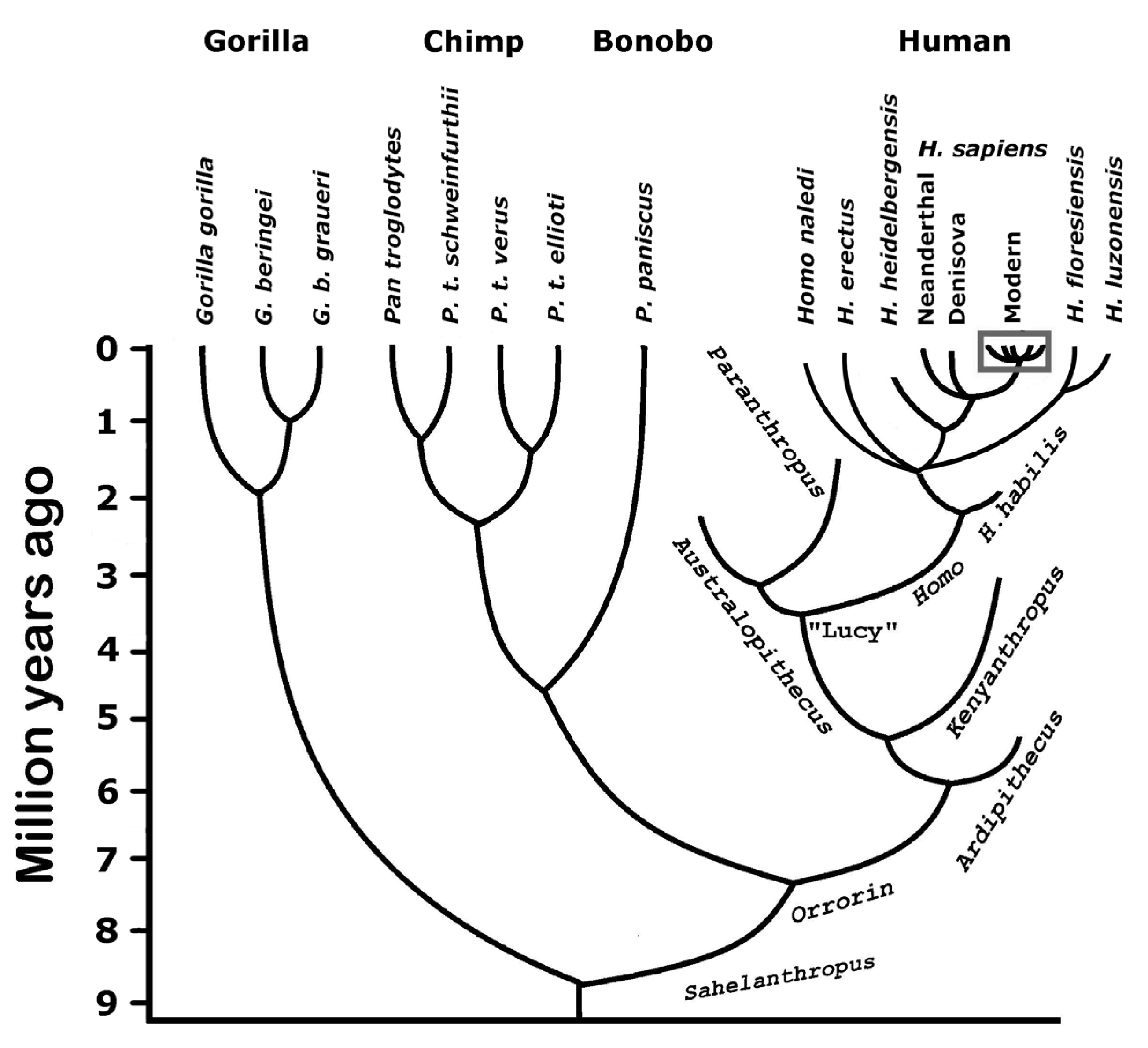
It is less clear when pair-bonding evolved in our lineage (see Figure 1). Our closest relatives, the two species of chimpanzees (chimps and bonobos), do not share this feature. Gorillas on the other hand, which split with our branch just before the split with the chimpanzees, do form polygynous bonding, a practice that is relatively common in humans as well [28]. It is conceivable that the trait was lost in the chimpanzees. As to the other extant apes, bonding is either more superficial or absent [30]. Most likely, the feelings involved in forming couples evolved relatively late in our lineage. That, combined with their importance for genes, explains the strong rewards.
It is not obvious that human pair-bonding is required for bringing up children. Even in the Stone Age, a single mother was presumably helped by members of her tribe and therefore could manage without support from the father. In some matrilineal cultures, the mother’s parents and siblings take more responsibility than the father [31]. The evolutionary rationale for pair-bonding in our lineage may depend to some extent on the importance of giving each male the right to foster children.
In humans, the strategy for pair-bonding is based on two stages. The first is what we refer to as falling in love. This form of love is meant to gradually develop into old or romantic love. Romantic love is presumably based on the same circuitry as the love for children [22,24]. It seems likely that other forms of bonding as well are based on the template responsible for the original mother–child affiliation [32].
Falling in love is a possible exception. The function may have started as a subsidiary of the love module but evolved to be a somewhat unique human feature. It has characteristics shared with addiction and obsession. The similarities include the particular use of the neurotransmitter serotonin [33] and perhaps the effect of vasopressin on the anterior part of the cerebral cortex [34]. The resemblance makes sense when considering the role of falling in love. In our tribal past, evolution needed a mechanism to secure out-breeding. As most of social life was restricted to other tribal members, instant affection on the rare occasions of meeting an eligible person from a foreign tribe served this purpose. As indicated by the English term, it is a “fall” in the sense that the relationship is based on an uncritical acceptance of the partner. This evolutionary quirk may explain why addictions and obsessions are so common in humans: they are byproducts of the module responsible for this form of love. Falling in love may be one of the most recent modules added to our brain; as expected, it is backed by strong rewards.
![Encyclopedia 04 00029 g001]()
Figure 1.
Our branch of the evolutionary tree. The figure attempts to present a consensus in a field with considerable differences in opinion, particularly as to branching points. There are two extant species of gorillas, and one of them has two subspecies. Chimpanzees are also divided into two species, but one of them has four subspecies. In the human branch, there is only one species alive today (box). More than 30 species belonging to the human branch have been proposed [35], but the figure only shows a selection. Until recently, there probably were at least eight species or subspecies present. Little is known about ancestral species of gorillas and chimpanzees.
Figure 1.
Our branch of the evolutionary tree. The figure attempts to present a consensus in a field with considerable differences in opinion, particularly as to branching points. There are two extant species of gorillas, and one of them has two subspecies. Chimpanzees are also divided into two species, but one of them has four subspecies. In the human branch, there is only one species alive today (box). More than 30 species belonging to the human branch have been proposed [35], but the figure only shows a selection. Until recently, there probably were at least eight species or subspecies present. Little is known about ancestral species of gorillas and chimpanzees.

3.3. Kinship
Kinship, or extended family, implies that the members of a group recognize relatedness beyond that of parents–offspring. The recognition is expressed as a tendency to affiliate and share resources more willingly with related individuals. The evolutionary rationale is covered by the concept of inclusive fitness, also referred to as kin selection [36]. It implies that it helps my genes to help someone who carries a share of my genes such as children and siblings. Relevant behavior is catered to by a variant of love.
In non-human species, kin generally implies siblings and those in a direct line of descent, and there is a tendency for same-sex affiliations (particularly among females) to be stronger than relations with the opposite sex [37]. Cousins and beyond have limited relevance outside human societies. In humans, kin typically includes those who have a stake in helping blood relatives, such as in-laws, and the importance of cultural traditions may lead to variants of kin recognition that are not exactly as expected from the principle of kin selection [38].
Kinship tends to be either female-centered (matrilineal) or male-centered (patrilineal) [39]. The most likely cause of this distinction is that one sex typically moves away for the sake of avoiding inbreeding. In traditional cultures, male-centered kinship is more common, and the same was likely the case with Neanderthals [40]. In modern societies, the distinction is less relevant as all children tend to move away from their parents.
The response to the problem of inbreeding is still a part of human life. It is revealed not only in cultural traditions, such as sexual taboos, but also in our genes. As to the latter, children who grow up as siblings are programmed to develop an aversion to sexual or romantic relations with each other [41]. Apparently, growing up together is what matters, not genetic relatedness.
The various options for constellations of kin have led to considerable cultural variation [31]. Who is treated as a relative differs; for example, only some cultures recognize second cousins. The restrictions and obligations involved also vary. Most cultures have kinship-based rules defining who one can have sexual/romantic liaisons with, but some allow cousin marriage while others do not.
3.4. Social Life
Social life is favored by evolution if it improves fitness, that is, the benefits of the genes should exceed the costs [42]. The level of cohesion and collaboration differs. Herbivores often gather in flocks due to the safety in numbers. Being together simply implies more vigilance and less chance for a particular individual to be killed by a predator. Carnivores that hunt together, such as wolves, require further collaborative effort, which means stronger bonds and more elaborate social relations.
The genetic benefits are not always obvious. Altruism is reasonably common, even though it implies that the provider loses fitness in the process of helping others [2]. Three mechanisms are proposed to explain this form of unselfish behavior:
- Inclusive fitness is primarily about relatives, but the associated feelings can “spill over” to non-kin.
- Reciprocity means that I help you today because I expect that you will help me later [43]. The principle stands strong in humans, where it forms the basis for commitment and obligations, but it is relevant for other species as well. The reciprocity can be indirect in that the return is given by a third party. Our concern about reputation is related to indirect reciprocity.
- Group selection is somewhat controversial [44]. It implies that natural selection can act not only on the individual but on a tribe or a troop. The idea is that the stronger groups outperform competing groups. The benefit will likely affect the genes of an individual because if the group thrives, his or her progeny will eventually benefit. I believe group selection was instrumental in establishing the foundation of human social life and perhaps the use of religion as a tool to promote prosocial behavior [6,45].
It is common for monkeys to socialize, but humans and chimpanzees are the only apes with strong social commitments [30]. This observation suggests that social life evolved in our lineage after the split with the gorillas some eight Mya (Figure 1). One would therefore expect that our social propensities are based largely on the use of the mood modules.
The factors discussed above were presumably instrumental in forming the various conscious and subconscious and emotional and instinctive modules that are associated with human social affiliations. Our capacity for empathy and compassion is part of this behavioral repertoire, and concepts such as morals and norms reflect manifestations. The extensive list of relevant emotions testifies to the importance of collaboration in our species, as does our capacity for language. Social life may have evolved late, but it has become exceedingly complex and important.
Although our capacity for gregariousness and language is in our genes, both functions need to develop after birth. How we socialize and how we speak must fit with the norms of the community we live in. The genes specify at what point in life the most important part of this development should take place, that is, we have critical periods or “learning windows”. In the case of language, the period may start around birth and fade during adolescence [46]; as to social skills, the period starts a couple of years after birth but lasts through adolescence [47]. Those who grow up with minimal social contact tend to remain egocentric and deficient in language [48].
Human social behavior has one rather unique feature: we combine social life with family constellations. Pair-bonding offers obvious advantages to rearing a child, but as indicated in Section 3.2, another factor may have been equally important in establishing this feature. Extensive collaboration requires that all males have a chance to mate. If an alpha male controlled all the females, evolution would be unlikely to allow unrelated males to forsake their genes and serve the leader. Pair-bonding solved the problem. In chimps, there is typically an alpha male with a primary right to copulate, but they do not depend on the same level of cooperation.
Evolution found a different solution for bonobos. Here, group amicability and the chance to procreate are achieved by everyone having sex with everyone else [49]. The positive feelings associated with sex presumably help establish good relations, and all the males are given a chance to pass on their genes. As anyone could have fathered babies, they are treated well. Not surprisingly, bonobos are known as the most amicable of primates.
The use of sex for bonding is not unique to bonobos. The same rationale is most likely behind our propensity to have intercourse regardless of whether the female is in heat, but in humans, the main function appears to be pair-bonding. As pointed out in Section 3.2, we are not a highly monogamous species; thus, sex may serve a wider group coherence as well.
The prevalence of homosexual behavior in both humans and several other mammals is an intriguing observation [50]. As to humans, homosexuality is common in traditional cultures and is typically regarded as normal or acceptable behavior [28]. It appears even in modern cultures despite condemnation. As the activity does not result in conception, the likely evolutionary rationale (if any) is to promote collaboration within the genders [51].
Some 25 Mya, the TRPC2 gene was lost in the lineage leading to apes and humans due to a mutation [52]. The removal of TRPC2 function has been associated with same-sex sexual behavior in mice [53], and the behavior is particularly common in the primate species lacking the gene. Other (unknown) factors may have been responsible for the selection favoring the mutation, but the loss could still facilitate homosexuality and thereby promote gender cooperation at a later stage in evolution.
We typically refer to positive social connections beyond family as friendship. This form of attachment is built on neurobiology related to that of the love for spouse and children [32], which means it is fair to state that you “love” your friends. Both love and the mood modules testify to the frugality of evolution. Novel brain functions are not constructed from scratch, but rather built by modifying existing circuitry. As pointed out above, the three mood modules cater to all situations where positive or negative feelings are required to instigate behavior. Similarly, the love module caters to various forms of affiliation.
3.5. Hostility
Evolution alleviated the problem of selfish behavior within the tribe with the measures described in Section 3.1, Section 3.2, Section 3.3 and Section 3.4, but the potential for conflicts between tribes remains. By forming social coherence within a group, evolution also gave us a tendency to differ between us and them—between friend and foe. The measures set up to ensure cooperation do not apply to strangers. We may give an outsider the benefit of the doubt, he/she could prove to be an ally, but the threshold for aggression is low.
Humans can presumably maintain personal relations with some 100–200 individuals [54], a number that in the Stone Age included one’s tribe as well as members of cooperating tribes. As long as the population density was low, they would rarely meet with hostile outsiders; thus, in the absence of conflicts, relationships were primarily genial. Then, the density increased. The biological success of our species implies a population expansion that most likely started long before the invention of agriculture. More people competing for resources implies a higher probability of inter-group conflicts [55]. Eventually, the situation gave rise to large constellations of people in communities and cities. Here, the lack of close ties between the members caused within-community conflicts as well, a problem that has been referred to as the ”tragedy of the commons” [56]. The evils of hostility are not solved, but in modern times we have some countermeasures, including law enforcement and international treaties [57].
Aggressive behavior can benefit your genes; consequently, hostility and violence should elicit brain rewards. As witnessed by the popularity of fighting sports and the description of heaven (Valhalla) in Norse mythology, they do. Consequently, it is not obvious that your level of happiness is best served by “turning the other cheek”. As a fight can damage both parties, evolution did add restraints, but these are easily lifted. There is plenty of violence in the world.
In addition to countermeasures such as those referred to above, there is one encouraging factor: compassion is likely to do more for your level of happiness than aggression. Both forms of behavior offer rewards, but kindness may produce more potent rewards. Hostility is an ancient feature of the brain. While the need to fight for resources dates back to early animals, our social tendencies evolved recently. Based on the idea that late evolutionary instigations rely to a larger extent on feelings, the modules associated with geniality are likely to activate stronger rewards. In short, we are happier hugging than hitting each other. Moreover, violence fosters retaliation while compassion is returned with kindness.
On the other hand, feelings are just one of several mechanisms employed to instigate behavior [8], which means we do not necessarily choose the options that maximize happiness. Even if hugging does more for your score, the urge to hit can win the day. When someone “steps on your toes”, the response tends to be somewhat like pushing a button. Anger is deep-rooted and consequently requires a considerable effort to restrain.
There is another obstacle to avoiding violence. Individual hostility can to some extent be curbed by laws and morals, but when the violence is on behalf of your group, our prosocial tendencies boost the willingness to engage. Fighting a shared enemy is not only socially acceptable but often encouraged. A soldier can harvest both the rewards connected with personal anger and the rewards of doing something for the group. What seems obvious is that warfare reduces the overall happiness in the world.
4. Present Society
4.1. The History of Homo Sapiens
During the Middle Paleolithic period, 300-50 Kya (thousand years ago), there were probably eight or more species and subspecies of the genus Homo (Figure 1). Only one is still present. It may be that our ancestors helped cause the extinction of the other species, either due to subtle differences in mental capacity (including social behavior) or simply by being the more aggressive variant.
The lineage leading to modern humans split up with the lineages leading to Neanderthals and Denisovans some 600 Kya [58]. According to standard biological definition, the boundary of a species depends on the capacity to produce fertile offspring. There is substantial evidence for admixture between these three lineages [59], implying that our species goes back to before the split. The genetic changes in our lineage that occurred after the split tend to involve genes assumed to affect brain function [59]. Yet, the difference in mental capacity was most likely minor as indicated by the following: one, the Neanderthals had the largest brain of any Homo species [60]; two, they displayed artistic and symbolic behavior [61]; and three, they probably had language akin to ours [62].
The present populations of humans separated gradually starting some 250 Kya [63]. There is evidence of improved technology and a burst in creativity upon entering the Upper Paleolithic period 50 Kya. The use of agriculture came later, perhaps 20 Kya [64]. Before the Upper Paleolithic, our ancestors probably lived the way of life their genes were adapted to, that is, as hunter/gatherers in tribal groups and in areas of low population density. Drastic changes did not start until the Neolithic (12 Kya), and until recent times these changes involved only select populations. The earliest remains of cities date to 10 Kya [65]. The increase in the size of societies, and the concomitant change in livelihood, had considerable consequences for human interactions.
The changes in the Upper Paleolithic and Neolithic are too recent to have caused more than a minor impact on human genetic constitution. Compared to other species, we are relatively homogenous genetically [66]. There are mutations that can be used to distinguish present populations, for example in genes responsible for pigmentation and lactose tolerance, but the genetic variability within any particular subpopulation is much larger than the variability between the subpopulations [67]. It seems unlikely that genetic changes in the period after the split of the present populations have had a notable impact on the modules related to social behavior. In short, our innate tendencies were for all practical purposes the same then, the same in all present populations, and will remain the same in the foreseeable future.
4.2. Social Discords
Forcing animals to live under conditions that differ from their natural environment can cause aberrant behavior and health problems [68], and the same is likely the case in humans [69]. Both our bodies and brains are adaptable, yet we are not immune to the negative consequences of a suboptimal environment. For example, although evolution designed us to be omnivores, the present diet is responsible for an increase in cardiovascular diseases [70], and although we have an extremely malleable brain, the present way of life is likely responsible for the high prevalence of mental issues [71].
The differences between the environment we are adapted to live in and the present situation have been referred to as mismatches [70]. Most of the mismatches are probably beneficial to quality of life, but some contribute to an increased prevalence of disorders. These disorders, for example, anxiety and type 2 diabetes, are referred to as the diseases of civilization [72]. The term discord is used for causative mismatches [69].
Most likely there are examples of discord that affect social behavior. Here is a list of candidates:
- A high population density. The “us and them” inclination is troublesome in a world where conflicts between groups are common.
- Present societies require that you interact with many people you do not know. We have an innate tendency to fear strangers [73]; thus, the situation is expected to increase not only hostility but also stress and anxiety.
- The large number of people, combined with how society is organized, means more competition and concomitant stress.
- We have lost the close-knit social structure of the tribe, and the consequences are loneliness, insecurity, and a lack of belonging. Family and friends do not offer complete compensation for the tribal bonds.
- While the typical tribe presumably had a relatively flat social structure, in most countries there is a distinct hierarchy. The inhabitants frequently encounter dominant individuals such as teachers, police, or government officials.
- We can no longer fulfill our needs solely by personal activity but rely on external suppliers.
Social discords such as those suggested above imply a less-than-optimal way of molding the brain. The possible consequences include mental problems, reduced quality of life, and antisocial behavior.
5. Conclusions and Prospects
5.1. Promoting Social Behavior
The suggested discords are difficult to avoid, yet it may be possible to improve the situation. The main issue is, arguably, how to prevent undesirable effects on mental health and social behavior. Half the population of Western nations has a diagnosable mental disorder at some point in life [74]. A negative mental state is likely to cause antisocial behavior, while a happy and healthy mind fosters kindness [75,76]. We should try to establish conditions that cater to the human psyche and boost the genial propensities of the brain. A comparison of known societies suggests that it is possible to augment prosocial behavior. For example, there is considerable variation in the use of violence in traditional cultures [77], and Japan has an exceptionally low crime rate compared to other countries [78].
I have discussed how to mold the brain elsewhere [79]. The educational system may be the most important venue for creating a desirable mentality. The attempt to form the brains of future generations should start early (in kindergartens), as childhood is when the window of social learning is most open [47]. Most, if not all, countries issue guidelines for what children should learn, but I believe these guidelines can be improved.
There are supplementary options. One example is the science-based prosocial initiative that proposes strategies for boosting collaborative behavior in adults [6]. Another is the use of brain exercise techniques such as the Buddhist practice of compassion meditation [80].
Evolution tends to find countermeasures if excessive affiliation and compassion pose a threat to the genes [81]. For example, in most species, mothers can be triggered to care for any baby, but in species such as sheep, where there is a distinct risk of spending resources on an unrelated offspring, mechanisms evolved to restrict affiliative behavior. The ewe will only care for a lamb she learns to recognize by smell at the time of birth. Fortunately, evolution did not need to impose distinct restraints on who a human can affiliate with. Consequently, it is possible to stimulate cohesion in groups that are much larger than the number of affiliates we can maintain personal relations with. For example, most countries have a level of national pride and unity.
The traditional way of life in Japan may reflect an extreme form of coherence. It has been suggested that offices and factories are organized as “tribal units” and the country as a “super-tribe” [82], that is, they depend more on morals than the law to enforce proper behavior. Responsibilities may stand stronger when based on morals. For one, we have an innate inclination to follow ethical rules while laws are a novel invention; and two, it is perhaps easier to escape law enforcement than a guilty conscience. Religion may enhance the impact of morals. We likely have a predisposition for religious beliefs that can be used to promote prosocial behavior, such as, for example, by combining rules of conduct with the tenet that “God can see you”. Science may help us make the most of this aspect of human mentality [45].
The term fundamental attribution error reflects the idea that we tend to underestimate the power of the environment in determining behavior but instead think of people as either good or bad [83]. Our innate social inclinations suggest that people are kind if they live in a suitable environment.
5.2. Aiming for Happiness
Using happiness as an aim has a double advantage. It is, arguably, what the individual should strive for, and it is a key feature for the task of enhancing society. A happy person is more likely to be caring and productive [84,85].
Research on happiness typically concludes that how we affiliate with others is the most important factor [86,87,88,89]. The strong emotions (and concomitant pleasures and pains) associated with social behavior presumably explain this finding. Consequently, it is vital to enhance positive and subdue negative relations. For that purpose, we need to stimulate people’s emotional intelligence—the ability to perceive, understand, manage, and express emotions in a positive way.
The bias, in the direction that geniality is more conducive to happiness than hatred and misconduct (see Section 3.5), appears to be a fortunate twist of evolution. The bias is supported by research; for example, giving offers stronger rewards than receiving a similar value or service [90], and volunteering work boosts well-being [91]. Another fortuitous twist is that evolution did not need to install restrictions on altruistic behavior because in the Stone Age excessive care for strangers was not a problem. The rewards of compassion and solidarity can therefore be called upon for any purpose. Our measure of free will [92], combined with these rewards, can be employed to promote behavior that is not in the interest of the genes but to the benefit of both the individual and society. It is difficult to motivate people to behave counter to what serves their well-being; thus, the point is to convince them that geniality is in their best interest. In short, biological constraints do not block the creation of a more convivial society.
Framing happiness as the primary aim can also help us alleviate environmental problems. The crucial point is that happiness does not require excessive consumption [93,94]. Lifetime contentment is linked to having a healthy mind and prioritizing pleasures that do not have obvious downsides [9]. For example, love presumably has fewer disadvantages than revenge, healthy eating is a better option than junk food, and rewards that can be harvested for free may serve you better than those you need to buy. It may be possible to create a world where everyone has an opportunity to flourish without causing excessive pollution and depletion of natural resources. The task would certainly be easier with a reduced world population.
5.3. Finale Remarks
Molding human minds in a desired direction is a considerable challenge; thus, the social setting of modern society is likely to continue to be an Achilles heel. Our technical achievements, such as landing a man on the moon and creating artificial intelligence, seem to be easy tasks compared to dealing with human nature. The aim should be to build a more compassionate, connected, and resilient social world. For that, we need a better understanding of the human psyche and a willingness to act.
The challenges are more likely to escalate than decline. The population on the planet is still increasing. Evolution is too slow to make appreciable changes in the genetic underpinning of social behavior, and it may not even be headed in the desired direction. The genetic constitution of humans will eventually change, but the forces that gave us our ability to understand and our capacity to socialize are not necessarily active today. The direction of evolution is a question of who is having the most children. If parental intelligence and friendliness do not correlate with the number of children fostered, evolution is headed away from these qualities.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Dobzhansky, T. Nothing in Biology Makes Sense except in the Light of Evolution. Am. Biol. Teach. 1973, 35, 125–129. [Google Scholar] [CrossRef]
- Wilson, E.O. Sociobiology: The New Synthesis; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Grüter, C.; Czaczkes, T.J. Communication in Social Insects and How It Is Shaped by Individual Experience. Anim. Behav. 2019, 151, 207–215. [Google Scholar] [CrossRef]
- Dugatkin, L.A. Cheating Monkeys and Citizen Bees: The Nature of Cooperation in Animals and Humans; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Grinde, B. An Evolutionary Perspective on the Importance of Community Relations for Quality of Life. Sci. World J. 2009, 9, 588–605. [Google Scholar] [CrossRef] [PubMed]
- Atkins, P.W.; Wilson, D.S.; Hayes, S.C. Prosocial: Using Evolutionary Science to Build Productive, Equitable, and Collaborative Groups; New Harbinger Publications: New York, NY, USA, 2019. [Google Scholar]
- Grinde, B. Did Consciousness First Evolve in the Amniotes? Psychol. Conscious. 2018, 5, 239–257. [Google Scholar] [CrossRef]
- Grinde, B. Consciousness: A Strategy for Behavioral Decisions. Encyclopedia 2023, 3, 60–76. [Google Scholar] [CrossRef]
- Grinde, B. The Biology of Happiness; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Helliwell, J.F.; Layard, R.; Sachs, J.D.; Neve, J.-E.D.; Aknin, L.B.; Wang, S. World Happiness Report 2023; Sustainable Development Solutions Network: New York, NY, USA, 2023. [Google Scholar]
- Abdallah, S.; Thompson, S.; Michaelson, J.; Marks, N.; Steuer, N. The Happy Planet Index 2.0; New Economic Foundation: London, UK, 2009. [Google Scholar]
- Bertolero, M.A.; Yeo, B.T.T.; Bassett, D.S.; D’Esposito, M. A Mechanistic Model of Connector Hubs, Modularity and Cognition. Nat. Hum. Behav. 2018, 2, 765–777. [Google Scholar] [CrossRef]
- Philipson, L. Functional Modules of the Brain. J. Theor. Biol. 2002, 215, 109–119. [Google Scholar] [CrossRef]
- Panksepp, J. Affective Neuroscience: The Foundations of Human and Animal Emotions; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Leknes, S.; Tracey, I. A Common Neurobiology for Pain and Pleasure. Nat. Rev. Neurosci. 2008, 9, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C.; Kringelbach, M.L. Neuroscience of affect: Brain Mechanisms of Pleasure and Displeasure. Curr. Opin. Neurobiol. 2013, 23, 294–303. [Google Scholar] [CrossRef]
- Dalgleish, T. The Emotional Brain. Nat. Rev. Neurosci. 2004, 5, 583–589. [Google Scholar] [CrossRef]
- Balcombe, J. Animal Pleasure and Its Moral Significance. Appl. Anim. Behav. Sci. 2009, 118, 208–216. [Google Scholar] [CrossRef]
- Spence-Aizenberg, A.; Di Fiore, A.; Fernandez-Duque, E. Social Monogamy, Male–Female Relationships, and Biparental Care in Wild Titi Monkeys (Callicebus discolor). Primates 2016, 57, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Royle, N.J.; Smiseth, P.T.; Kölliker, M. The Evolution of Parental Care; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Flannery, T.F.; Rich, T.H.; Vickers-Rich, P.; Ziegler, T.; Veatch, E.G.; Helgen, K.M. A Review of Monotreme (Monotremata) Evolution. Alcheringa 2022, 46, 3–20. [Google Scholar] [CrossRef]
- Shih, H.-C.; Kuo, M.-E.; Wu, C.W.; Chao, Y.-P.; Huang, H.-W.; Huang, C.-M. The Neurobiological Basis of Love: A Meta-Analysis of Human Functional Neuroimaging Studies of Maternal and Passionate Love. Brain Sci. 2022, 12, 830. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.; Kushnick, G. Proximate and Ultimate Perspectives on Romantic Love. Front. Psychol. 2021, 12, 573123. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, S.A.; Young, L.J. The Neurobiology of Love and Pair Bonding from Human and Animal Perspectives. Biology 2023, 12, 844. [Google Scholar] [CrossRef]
- Kleiman, D.G. Monogamy in Mammals. Q. Rev. Biol. 1977, 52, 39–69. [Google Scholar] [CrossRef]
- Lack, D. Ecological Adaptations for Breeding in Birds; Methuen and Company: London, UK, 1968. [Google Scholar]
- Lukas, D.; Clutton-Brock, T.H. The Evolution of Social Monogamy in Mammals. Science 2013, 341, 526–530. [Google Scholar] [CrossRef]
- Ford, C.S.; Beach, F.A. Patterns of Sexual Behavior; Harper: New York, NY, USA, 1951. [Google Scholar]
- Yong, J.C.; Li, N.P. Elucidating Evolutionary Principles with the Traditional Mosuo: Adaptive Benefits and Origins of Matriliny and “Walking Marriages”. Cult. Evol. 2022, 19, 22–40. [Google Scholar] [CrossRef]
- Strier, K.B. Primate Behavioral Ecology; Routledge: New York, NY, USA, 2016. [Google Scholar]
- Hasty, J.; Lewis, D.G.; Snipes, M.M. Introduction to Anthropology; OpenStax Rice University: Houston, TX, USA, 2022. [Google Scholar]
- Feldman, R. The Neurobiology of Human Attachments. Trends Cogn. Sci. 2017, 21, 80–99. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Akiskal, H.S.; Rossi, A.; Cassano, G.B. Alteration of the Platelet Serotonin Transporter in Romantic Love. Psychol. Med. 1999, 29, 741–745. [Google Scholar] [CrossRef]
- Pitkow, L.J.; Sharer, C.A.; Ren, X.; Insel, T.R.; Terwilliger, E.F.; Young, L.J. Facilitation of Affiliation and Pair-Bond Formation by Vasopressin Receptor Gene Transfer into the Ventral Forebrain of a Monogamous Vole. J. Neurosci. 2001, 21, 7392–7396. [Google Scholar] [CrossRef]
- Wood, B.; Boyle, E.K. Hominin Taxic Diversity: Fact or Fantasy? Am. J. Phys. Anthropol. 2016, 159, 37–78. [Google Scholar] [CrossRef]
- Levin, S.R.; Grafen, A. Inclusive Fitness Is an Indispensable Approximation for Understanding Organismal Design. Evolution 2019, 73, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.S.; De Moor, D.; Casanova, C.; Brent, L.J. Kinship Composition in Mammals. R. Soc. Open Sci. 2023, 10, 230486. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.T.; Steadman, L.B. Human Kinship as a Descendant-Leaving Strategy: A Solution to an Evolutionary Puzzle. J. Soc. Evol. Syst. 1997, 20, 39–51. [Google Scholar] [CrossRef]
- Fortunato, L. Lineal Kinship Organization in Cross-Specific Perspective. Philos. Trans. R. Soc. B 2019, 374, 20190005. [Google Scholar] [CrossRef] [PubMed]
- Skov, L.; Peyrégne, S.; Popli, D.; Iasi, L.N.; Devièse, T.; Slon, V.; Zavala, E.I.; Hajdinjak, M.; Sümer, A.P.; Grote, S. Genetic Insights into the Social Organization of Neanderthals. Nature 2022, 610, 519–525. [Google Scholar] [CrossRef]
- Lieberman, D.; Lobel, T. Kinship on the Kibbutz: Coresidence Duration Predicts Altruism, Personal Sexual Aversions and Moral Attitudes among Communally Reared Peers. Evol. Hum. Behav. 2012, 33, 26–34. [Google Scholar] [CrossRef]
- Krause, J.; Ruxton, G.D. Living in Groups; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Romano, A.; Saral, A.S.; Wu, J. Direct and Indirect Reciprocity among Individuals and Groups. Curr. Opin. Psychol. 2022, 43, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Smith, D. Cultural Group Selection and Human Cooperation: A Conceptual and Empirical Review. Evol. Hum. Sci. 2020, 2, e2. [Google Scholar] [CrossRef]
- Grinde, B. How Can Science Help Religion toward Optimal Benefit for Society? Zygon 2005, 40, 277–288. [Google Scholar] [CrossRef]
- Hensch, T.K. Critical Period Regulation. Annu. Rev. Neurosci. 2004, 27, 549–579. [Google Scholar] [CrossRef] [PubMed]
- Berk, L. Child Development; Pearson Higher Education: Sidney, OH, USA, 2015. [Google Scholar]
- Dombrowski, S.C.; Gischlar, K.L.; Mrazik, M.; Greer, F.W. Feral Children. In Assessing and Treating Low Incidence/High Severity Psychological Disorders of Childhood; Springer: New York, NY, USA, 2011; pp. 81–93. [Google Scholar]
- de Waal, F. Bonobo Sex and Society. Sci. Am. 2016, 25, 42–49. [Google Scholar]
- Poiani, A. Animal Homosexuality: A Biosocial Perspective; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Barron, A.B.; Hare, B. Prosociality and a Sociosexual Hypothesis for the Evolution of Same-Sex Attraction in Humans. Front. Psychol. 2020, 10, 2955. [Google Scholar] [CrossRef] [PubMed]
- Pfau, D.; Jordan, C.L.; Breedlove, S.M. The De-Scent of Sexuality: Did Loss of a Pheromone Signaling Protein Permit the Evolution of Same-Sex Sexual Behavior in Primates? Arch. Sex. Behav. 2021, 50, 2267–2276. [Google Scholar] [CrossRef] [PubMed]
- Stowers, L.; Holy, T.E.; Meister, M.; Dulac, C.; Koentges, G. Loss of Sex Discrimination and Male-Male Aggression in Mice Deficient for Trp2. Science 2002, 295, 1493–1500. [Google Scholar] [CrossRef]
- Hill, R.A.; Dunbar, R.I. Social Network Size in Humans. Hum. Nat. 2003, 14, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Peter, T. Population Dynamics and Internal Warfare: A Reconsideration. Soc. Evol. Hist. 2006, 5, 112–147. [Google Scholar]
- Rankin, D.J. Resolving the Tragedy of the Commons: The Feedback between Intraspecific Conflict and Population Density. J. Evol. Biol. 2007, 20, 173–180. [Google Scholar] [CrossRef]
- Pinker, S. The Better Angels of Our Nature; Penguin: London, UK, 2011. [Google Scholar]
- Prüfer, K.; De Filippo, C.; Grote, S.; Mafessoni, F.; Korlević, P.; Hajdinjak, M.; Vernot, B.; Skov, L.; Hsieh, P.; Peyrégne, S. A High-Coverage Neandertal Genome from Vindija Cave in Croatia. Science 2017, 358, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, N.K.; Shapiro, B.; Green, R.E. An Ancestral Recombination Graph of Human, Neanderthal, and Denisovan Genomes. Sci. Adv. 2021, 7, eabc0776. [Google Scholar] [CrossRef]
- Macciardi, F.; Martini, F. The Neanderthal Brain: Biological and Cognitive Evolution. In Updating Neanderthals; Romangnoli, F., Rivals, F., Benazzi, S., Eds.; Elsevier: New York, NY, USA, 2022; pp. 89–108. [Google Scholar]
- García-Diez, M. ‘Art’: Neanderthal Symbolic Graphic Behaviour. In Updating Neanderthals; Romangnoli, F., Rivals, F., Benazzi, S., Eds.; Elsevier: New York, NY, USA, 2022; pp. 251–260. [Google Scholar]
- Benítez-Burraco, A. The Case of Neanderthal Language (s): A Multidisciplinary Approach. PsyArXiv 2023. [Google Scholar] [CrossRef]
- Bergström, A.; McCarthy, S.A.; Hui, R.; Almarri, M.A.; Ayub, Q.; Danecek, P.; Chen, Y.; Felkel, S.; Hallast, P.; Kamm, J. Insights into Human Genetic Variation and Population History from 929 Diverse Genomes. Science 2020, 367, eaay5012. [Google Scholar] [CrossRef] [PubMed]
- Snir, A.; Nadel, D.; Groman-Yaroslavski, I.; Melamed, Y.; Sternberg, M.; Bar-Yosef, O.; Weiss, E. The Origin of Cultivation and Proto-Weeds, Long before Neolithic Farming. PLoS ONE 2015, 10, e0131422. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.; Evans, J.; Lamb, A.; Baird, D.; Hodder, I.; Marciniak, A.; Larsen, C.S.; Knüsel, C.J.; Haddow, S.D.; Pilloud, M.A. Mobility and Kinship in the World’s First Village Societies. Proc. Natl. Acad. Sci. USA 2023, 120, e2209480119. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R. Human Genetics: The Basics; Taylor & Francis: New York, NY, USA, 2016. [Google Scholar]
- Owens, K.; King, M.-C. Genomic Views of Human History. Science 1999, 286, 451–453. [Google Scholar] [CrossRef]
- Moberg, G.; Mench, J. The Biology of Animal Stress; Moberg, G., Mench, J., Eds.; CABI: Oxfordshire, UK, 2000. [Google Scholar]
- Grinde, B. Can the Concept of Discords Help Us Find the Causes of Mental Diseases? Med. Hypotheses 2009, 73, 106–109. [Google Scholar] [CrossRef]
- Eaton, S.B.; Konner, M.; Shostak, M. Stone Agers in the Fast Lane: Chronic Degenerative Diseases in Evolutionary Perspective. Am. J. Med. 1988, 84, 739–749. [Google Scholar] [CrossRef]
- Grinde, B. An Approach to the Prevention of Anxiety-Related Disorders Based on Evolutionary Medicine. Prev. Med. 2005, 40, 904–909. [Google Scholar] [CrossRef]
- Williams, G.C.; Nesse, R.M. The Dawn of Darwinian Medicine. Q. Rev. Biol. 1991, 66, 1–22. [Google Scholar] [CrossRef]
- Marks, I. The Development of Normal Fear: A Review. J. Child Psychol. Psychiatry 1987, 28, 667–697. [Google Scholar] [CrossRef]
- Moffitt, T.E.; Caspi, A.; Taylor, A.; Kokaua, J.; Milne, B.J.; Polanczyk, G.; Poulton, R. How Common Are Common Mental Disorders? Evidence That Lifetime Prevalence Rates Are Doubled by Prospective Versus Retrospective Ascertainment. Psychol. Med. 2010, 40, 899–909. [Google Scholar] [CrossRef]
- Douglas, K. Playing Fair. New Sci. 2001, 169, 38–42. [Google Scholar]
- Fowler, J.H.; Christakis, N.A. Dynamic Spread of Happiness in a Large Social Network: Longitudinal Analysis over 20 Years in the Framingham Heart Study. BMJ 2008, 337, a2338. [Google Scholar] [CrossRef]
- Nivette, A.E. Violence in Non-State Societies: A Review. Br. J. Criminol. 2011, 51, 578–598. [Google Scholar] [CrossRef]
- Hino, K.; Schneider, R.H. Planning for Crime Prevention in Japan. Built Environ. 2013, 39, 114–139. [Google Scholar] [CrossRef]
- Grinde, B. How to Form the Human Brain. PsyArXiv Prepr. 2024. [Google Scholar] [CrossRef]
- Hofmann, S.G.; Grossman, P.; Hinton, D.E. Loving-Kindness and Compassion Meditation: Potential for Psychological Interventions. Clin. Psychol. Rev. 2011, 31, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Krasnegor, N.A.; Bridges, R.S. Mammalian Parenting; Oxford University Press: Oxford, UK, 1990. [Google Scholar]
- Clark, R. The Japanese Company; Tuttle Publishing: Tokyo, Japan, 2013. [Google Scholar]
- Berry, Z.; Frederickson, J. Explanations and Implications of the Fundamental Attribution Error: A Review and Proposal. J. Integr. Soc. Sci. 2015, 5, 44–57. [Google Scholar]
- DiMaria, C.H.; Peroni, C.; Sarracino, F. Happiness Matters: Productivity Gains from Subjective Well-Being. J. Happiness Stud. 2020, 21, 139–160. [Google Scholar] [CrossRef]
- Grinde, B. Social Behavior: Making the Best of the Human Condition. Mank. Q. 2000, 41, 193–210. [Google Scholar] [CrossRef]
- Layard, R. Happiness—Lessons from a New Science; Penguin: London, UK, 2005. [Google Scholar]
- Grinde, B.; Nes, R.B.; MacDonald, I.; Wilson, D.S. Quality of Life in Intentional Communities. Soc. Indic. Res. 2017, 137, 625–640. [Google Scholar] [CrossRef]
- Gallagher, E.N.; Vella-Brodrick, D.A. Social Support and Emotional Intelligence as Predictors of Subjective Well-Being. Personal. Individ. Differ. 2008, 44, 1551–1561. [Google Scholar] [CrossRef]
- Grinde, B. Role of Happiness When Evaluating Society. Encyclopedia 2022, 2, 230–236. [Google Scholar] [CrossRef]
- Moll, J.; Krueger, F.; Zahn, R.; Pardini, M.; de Oliveira-Souza, R.; Grafman, J. Human Fronto-Mesolimbic Networks Guide Decisions about Charitable Donation. Proc. Natl. Acad. Sci. USA 2006, 103, 15623–15628. [Google Scholar] [CrossRef] [PubMed]
- Borgonovi, F. Doing Well by Doing Good. The Relationship between Formal Volunteering and Self-Reported Health and Happiness. Soc. Sci. Med. 2008, 66, 2321–2334. [Google Scholar] [CrossRef]
- Grinde, B. An Evolutionary Perspective on Free Will and Self-Consciousness. Psychol. Conscious. 2022. [Google Scholar] [CrossRef]
- Lykken, D. Happiness: The Nature and Nurture of Joy and Contentment; St. Martin’s Griffin: New York, NY, USA, 2000. [Google Scholar]
- Veenhoven, R. Well-Being in the Welfare State: Level Not Higher, Distribution Not More Equitable. J. Comp. Policy Anal. 2000, 2, 91–125. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Grinde, B. The Evolution of Human Social Behavior. Encyclopedia 2024, 4, 430-443. https://doi.org/10.3390/encyclopedia4010029
AMA Style
Grinde B. The Evolution of Human Social Behavior. Encyclopedia. 2024; 4(1):430-443. https://doi.org/10.3390/encyclopedia4010029
Chicago/Turabian StyleGrinde, Bjørn. 2024. "The Evolution of Human Social Behavior" Encyclopedia 4, no. 1: 430-443. https://doi.org/10.3390/encyclopedia4010029