
Response of the Zygopteran Community (Odonata: Insecta) to Change in Environmental Integrity Driven by Urbanization in Eastern Amazonian Streams




Abstract
:1. Introduction
2. Material and Methods
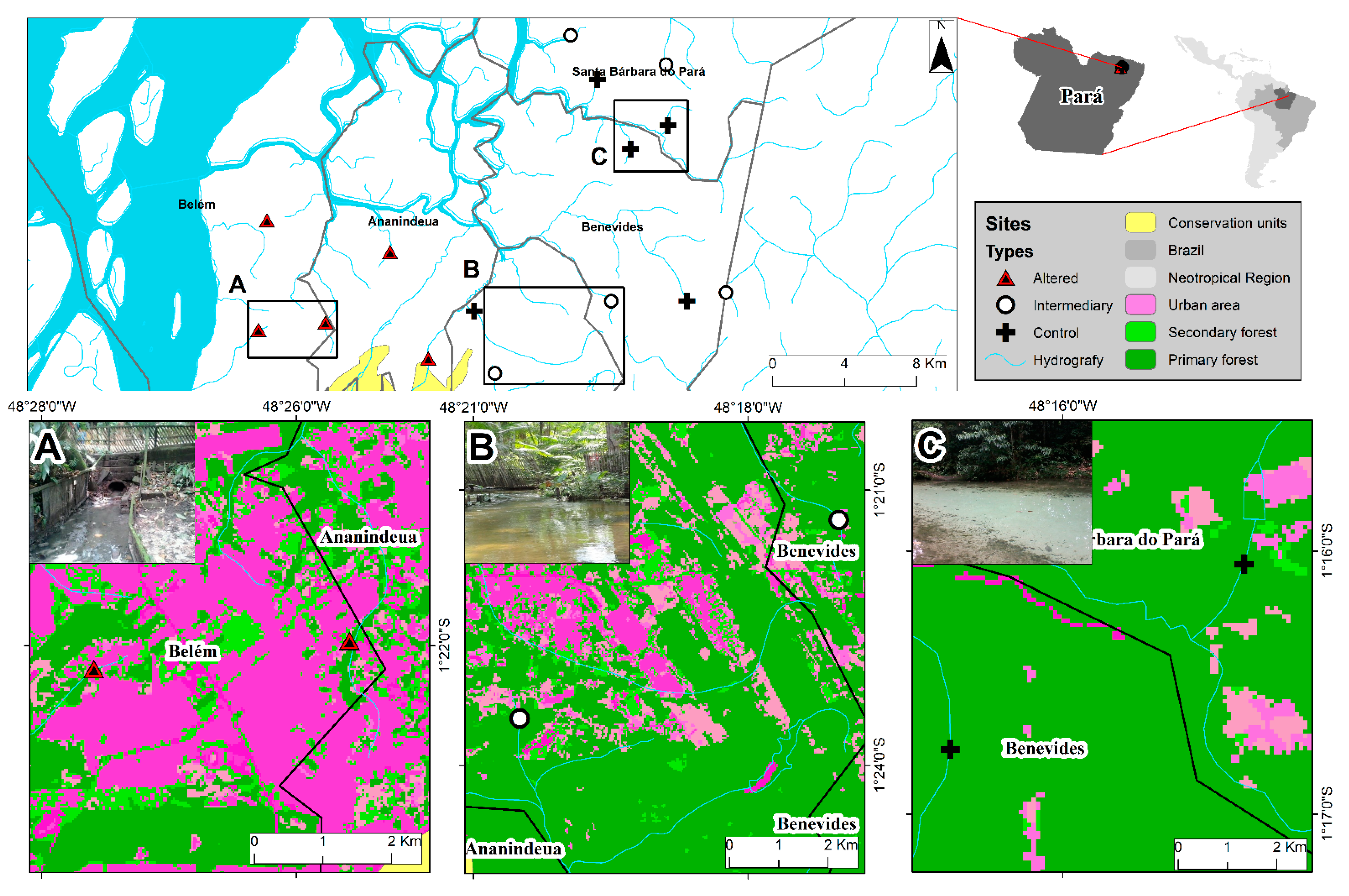
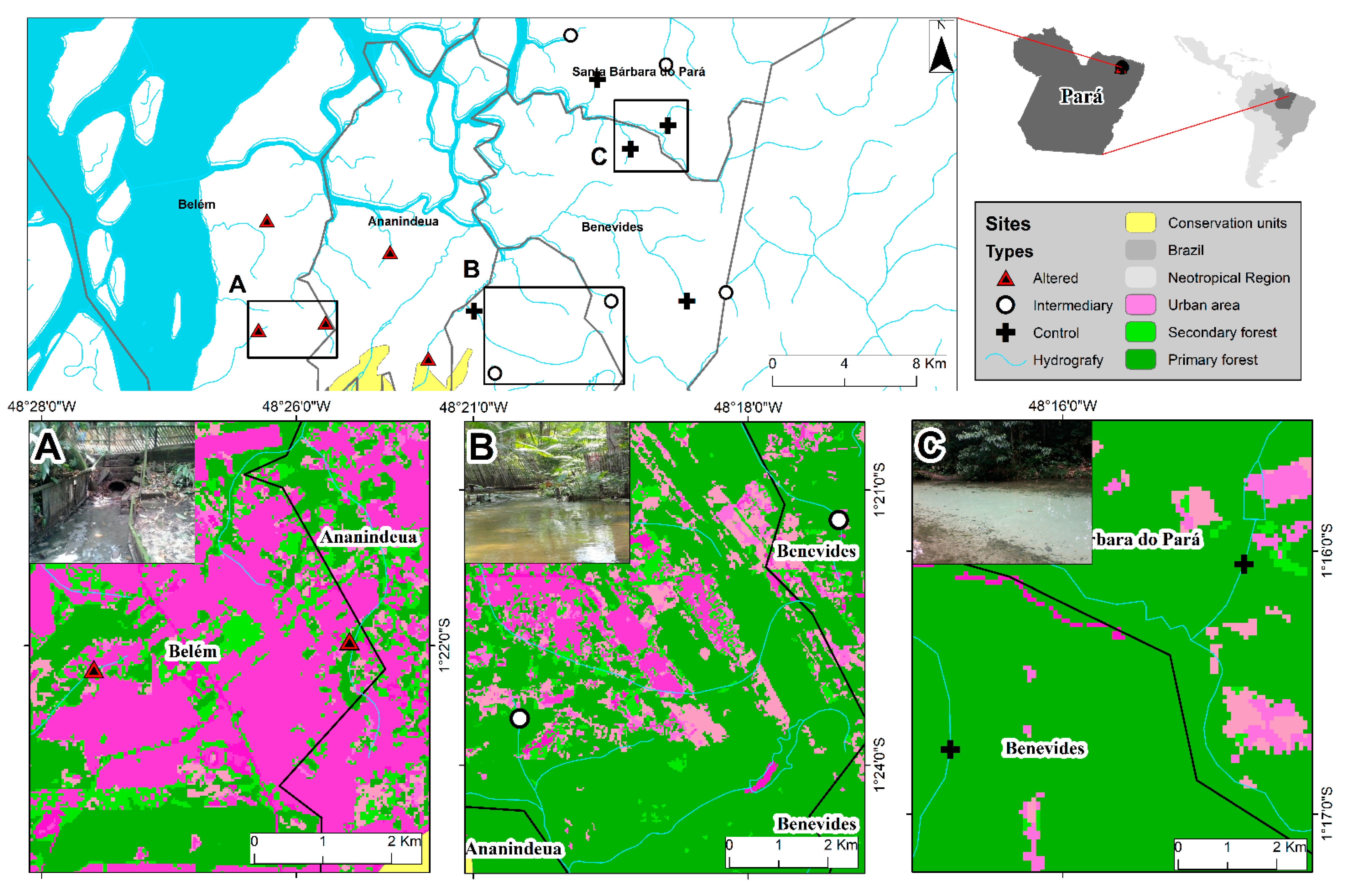
2.1. Study Area
2.2. Biological Sampling
2.3. Integrity Assessment of the Aquatic Environment
2.4. Biomass Measurements
2.5. Statistical Analyses
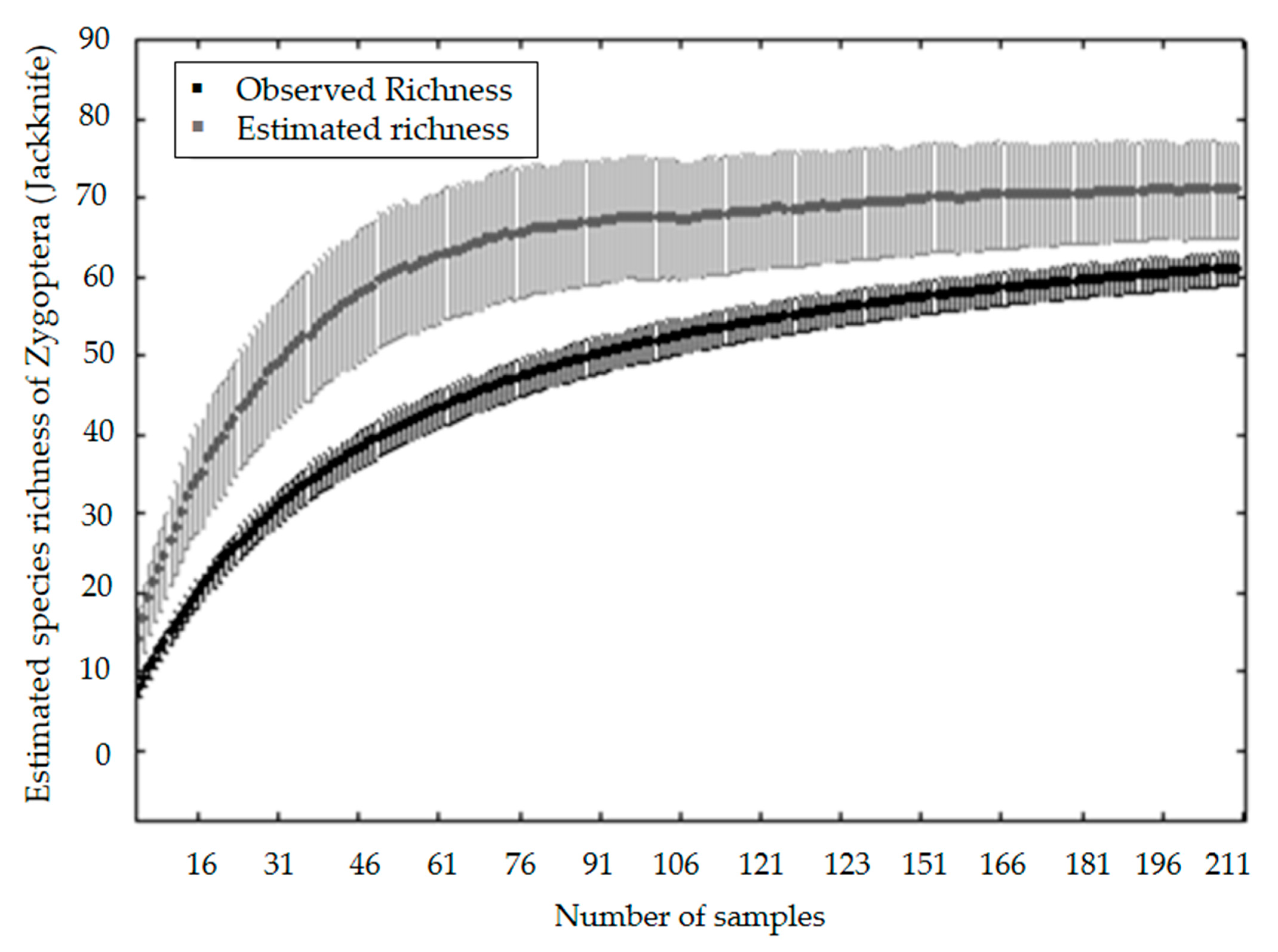
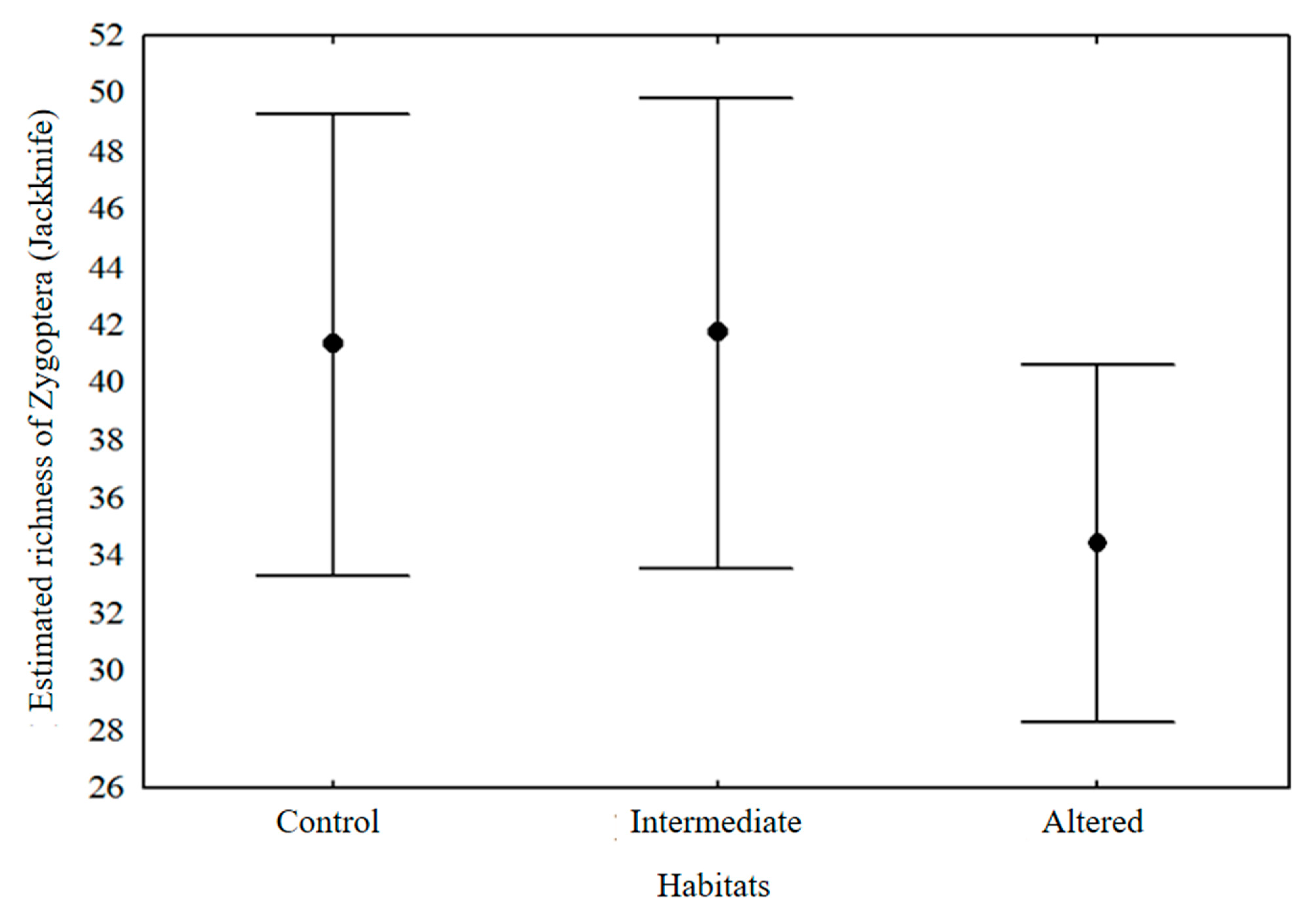
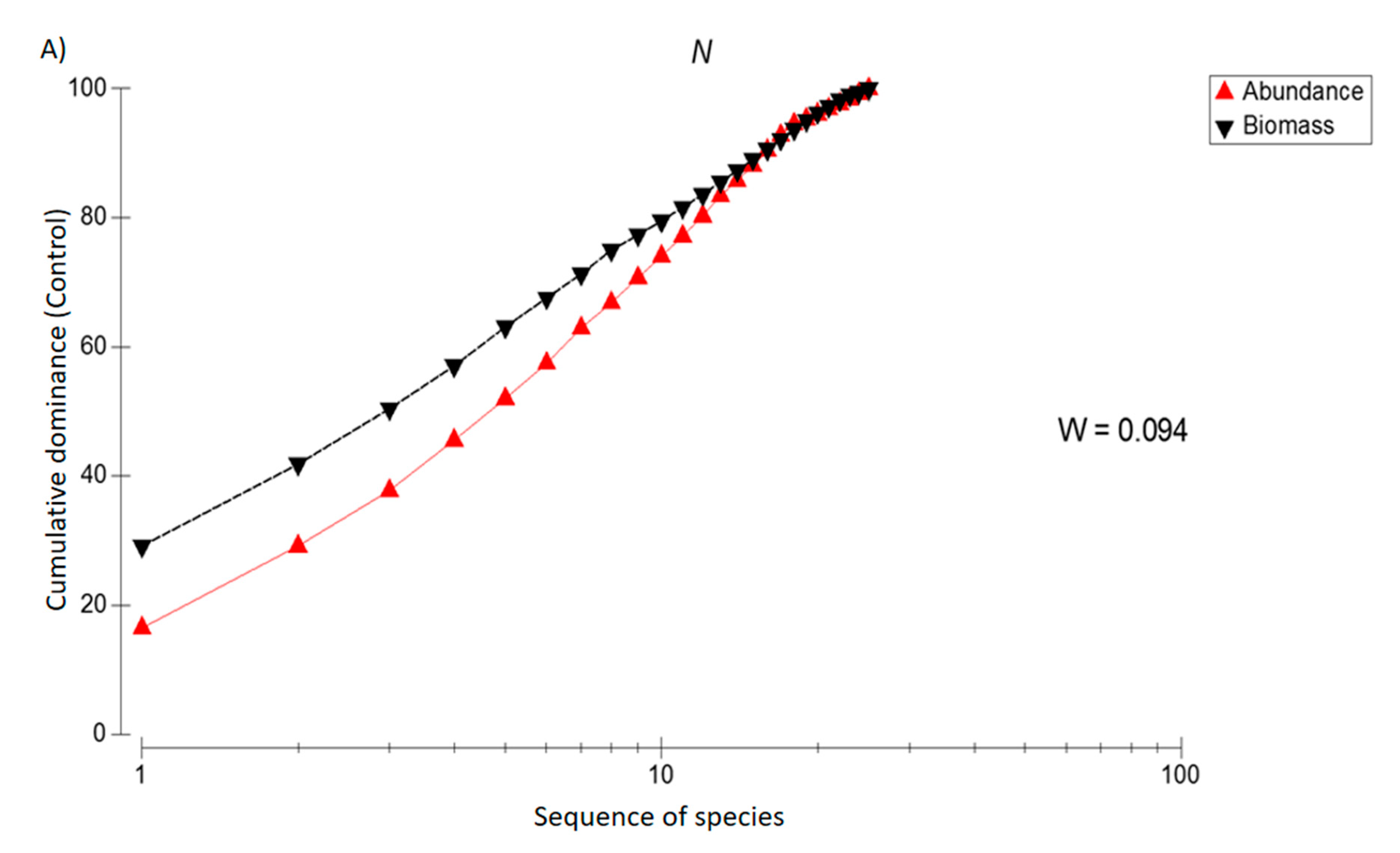
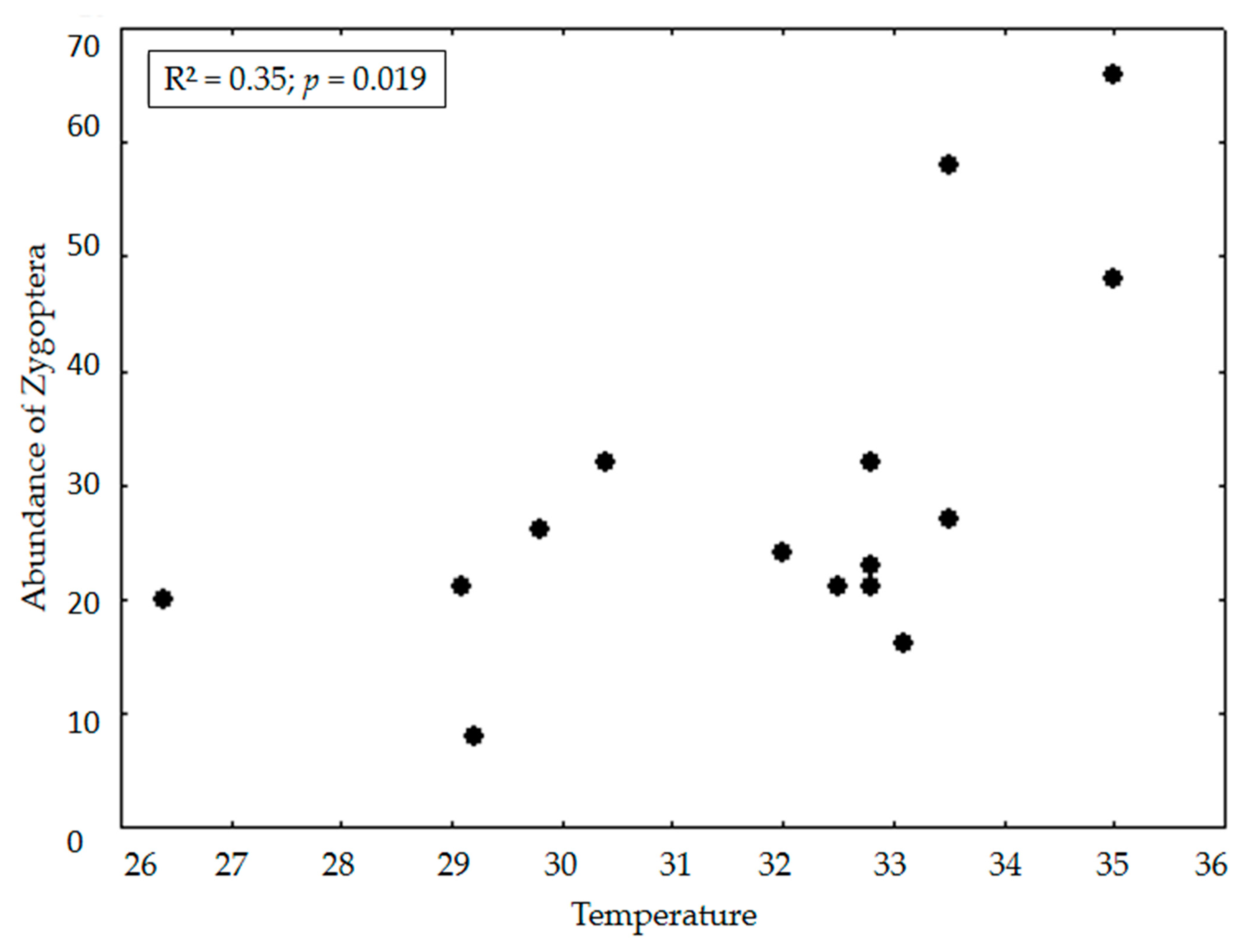
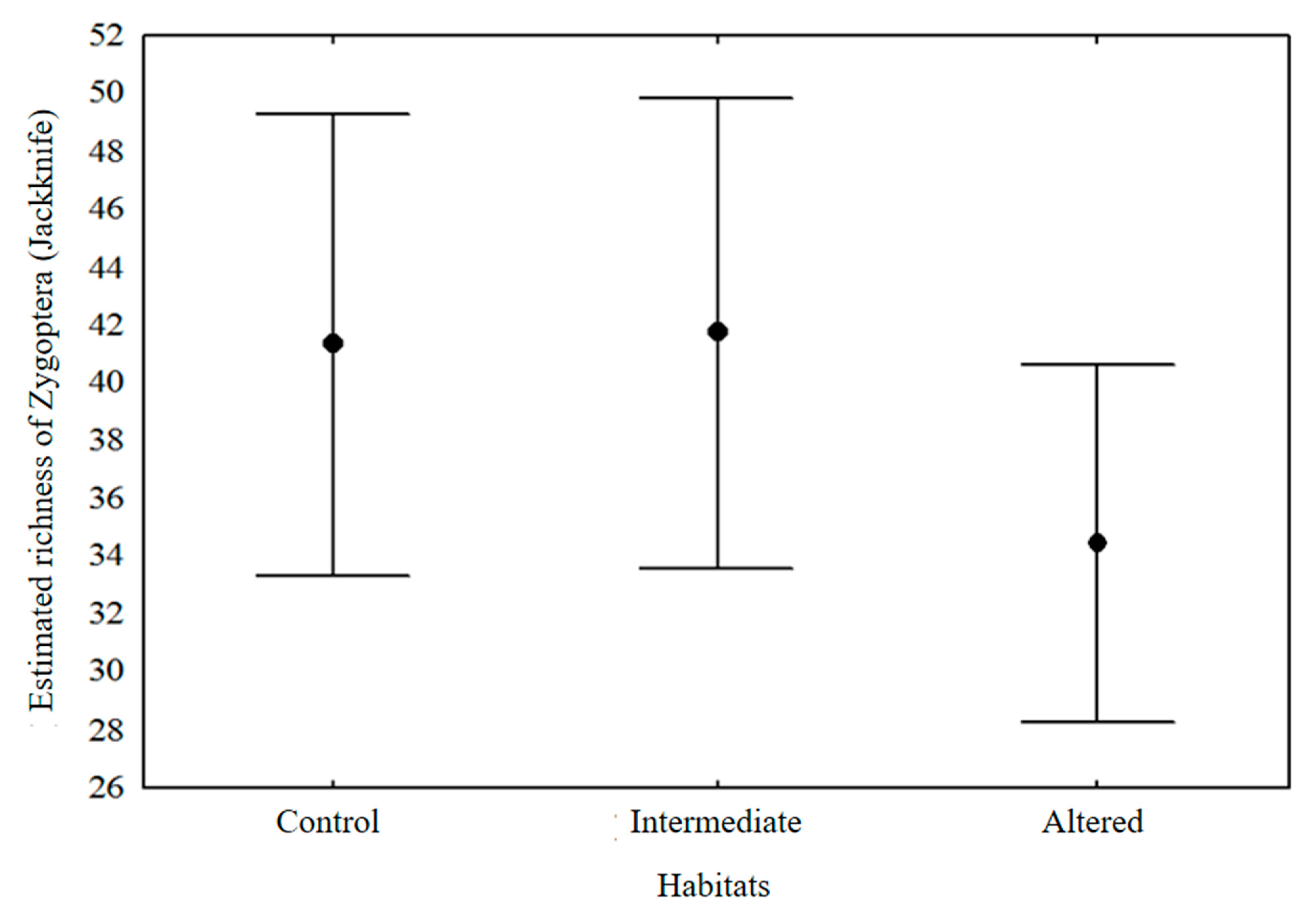
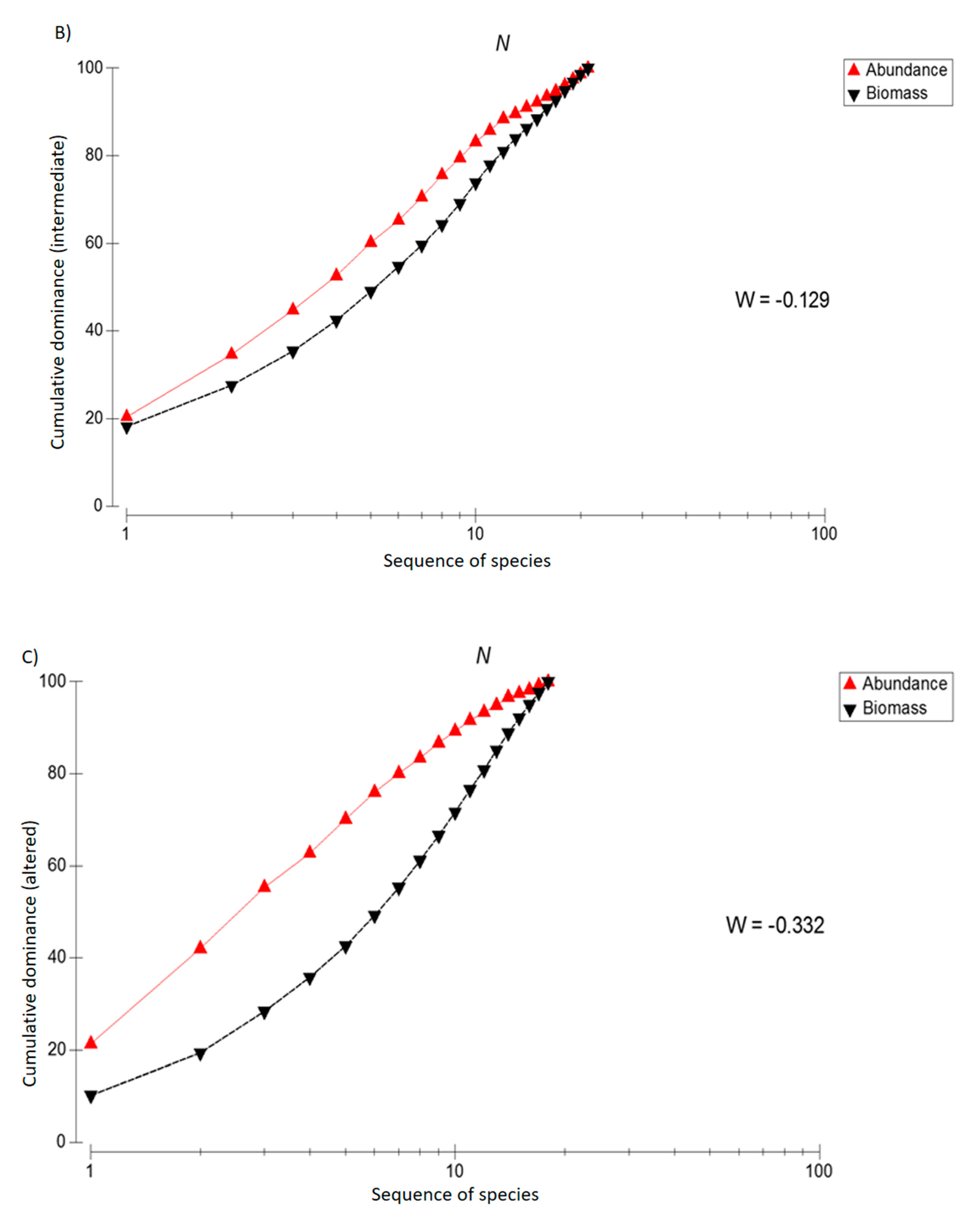
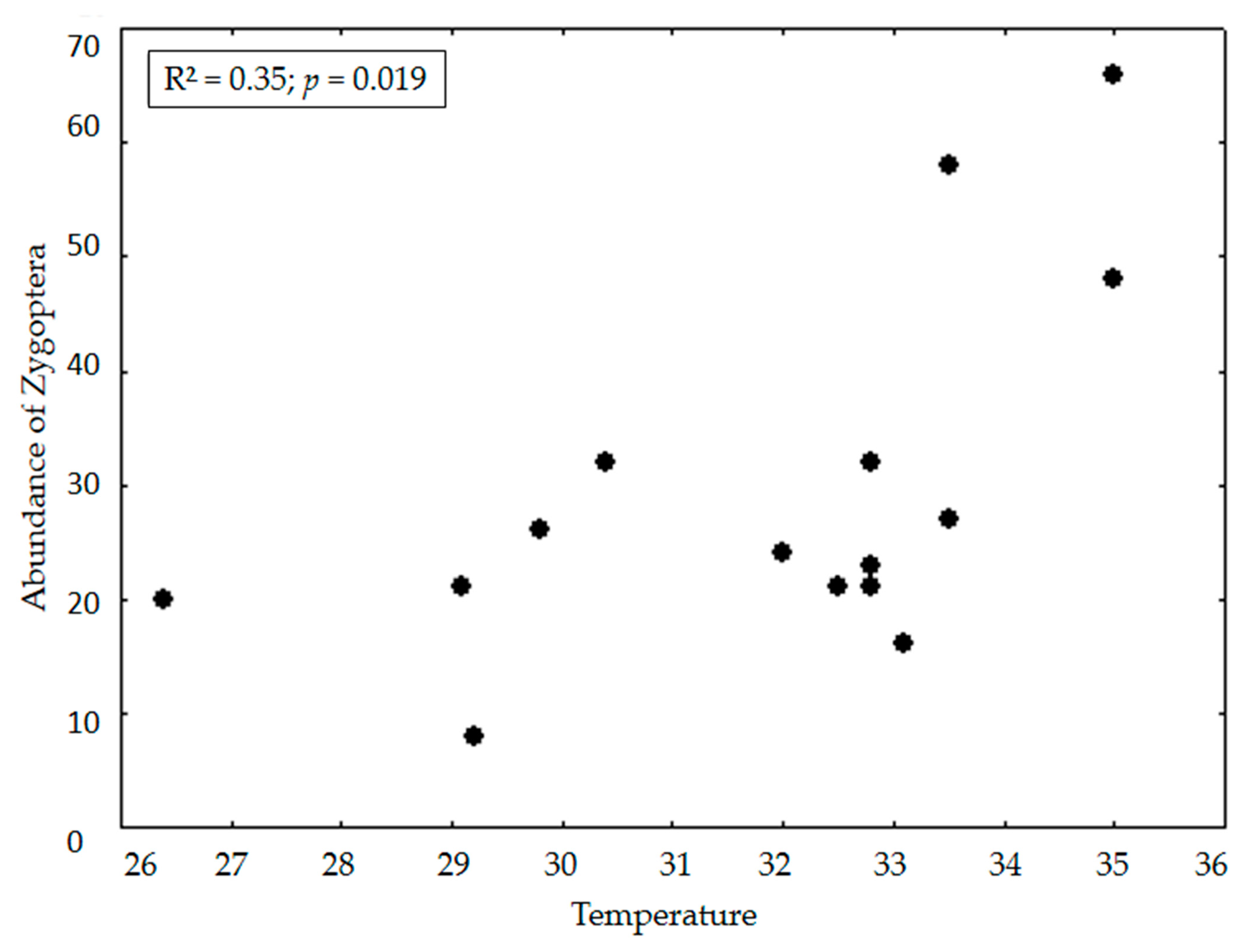
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Groups | Streams | HII |
|---|---|---|---|
| 1 | Altered | Parques dos Igarapés | 0.133 |
| 2 | Altered | Apeti | 0.175 |
| 3 | Altered | Águas Brancas | 0.172 |
| 4 | Altered | Paraíso dos Reis | 0.275 |
| 5 | Altered | Tenoné | 0.291 |
| 6 | Intermediate | Sítio Uruana | 0.508 |
| 7 | Intermediate | Muicuiambá | 0.536 |
| 8 | Intermediate | Pau D’Arco | 0.537 |
| 9 | Intermediate | Paar | 0.541 |
| 10 | Intermediate | Igarapé Neópolis | 0.68 |
| 11 | Control | Igarapé Coreia | 0.773 |
| 12 | Control | Cupuaçu | 0.715 |
| 13 | Control | Paricatuba | 0.743 |
| 14 | Control | Macumba | 0.812 |
| 15 | Control | Santa Maria | 0.941 |
References
- United Nations. World Urbanization Prospects: The 2018 Revision (ST/ESA/SER.A/420); Department of Economic and Social Affairs, Population Division: New York, NY, USA, 2019. [Google Scholar]
- De Long, D.C.J. Defining Biodiversity. Wildl. Soc. Bull. 1996, 24, 738–749. [Google Scholar]
- Walpole, M.; Almond, R.E.A.; Besançon, C.; Butchart, S.H.M.; Campbell-Lendrum, D.; Carr, G.M.; Collen, B.; Collette, L.; Davidson, N.C.; Dulloo, E.; et al. Tracking Progress Toward the 2010 Biodiversity Target and Beyond. Science 2009, 325, 1503–1504. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; Van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Monteiro-Júnior, C.D.S.M.; Juen, L.; Hamada, N. Analysis of urban impacts on aquatic habitats in the central Amazon basin: Adult odonates as bioindicators of environmental quality. Ecol. Indic. 2015, 48, 303–311. [Google Scholar] [CrossRef]
- Gordon, N.D.; McMahon, T.A.; Finlayson, B.L.; Gippel, C.J.; Nathan, R.J. Stream Hydrology: An Introduction for Ecologists, 2nd ed.; Wiley: Hoboken, NJ, USA, 1995. [Google Scholar]
- Carvalho, F.G.; Roque, F.D.O.; Barbosa, L.; Montag, L.F.D.A.; Juen, L. Oil palm plantation is not a suitable environment for most forest specialist species of Odonata in Amazonia. Anim. Conserv. 2018, 21, 526–533. [Google Scholar] [CrossRef]
- Barthem, R.B.; Da Costa, M.C.; Cassemiro, F.; Leite, R.G.; Da Silva, N.J. Diversity and Abundance of Fish Larvae Drifting in the Madeira River, Amazon Basin: Sampling Methods Comparison. Biodivers. Dyn. Balance Planet 2014, 2014, 137–158. [Google Scholar] [CrossRef] [Green Version]
- IBGE—Instituto Brasileiro de Geografia e Estatística, IBGE Discloses Municipal Population Estimates for 2019: Social Statistics. Available online: https://agenciadenoticias.ibge.gov.br/en/agencia-press-room/2185-news-agency/releases-en/25283-ibge-divulga-as-estimativas-da-populacao-dos-municipios-para-202 (accessed on 29 August 2019).
- Siqueira, G.W.; Aprile, F. Avaliação de risco ambiental por contaminação metálica e material orgânico em sedimentos da bacia do Rio Aurá, Região metropolitana de Belém-PA. Acta Amaz. 2013, 43, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Monteiro-Júnior, C.; Juen, L.; Hamada, N. Effects of urbanization on stream habitats and associated adult dragonfly and damselfly communities in central Brazilian Amazonia. Landsc. Urban Plan. 2014, 127, 28–40. [Google Scholar] [CrossRef]
- Monteiro-Júnior, C.D.S.M.; Couceiro, S.R.M.; Hamada, N.; Juen, L. Effect of vegetation removal for road building on richness and composition of Odonata communities in Amazonia, Brazil. Int. J. Odonatol. 2013, 16, 135–144. [Google Scholar] [CrossRef]
- Couceiro, S.R.; Hamada, N.; Forsberg, B.R.; Pimentel, T.P.; Luz, S.L.B. A macro invertebrate multimetric index to evaluate the biological condition of streams in the Central Amazon region of Brazil. Ecol. Indics 2012, 18, 118–125. [Google Scholar] [CrossRef]
- Moreyra, A.K.; Padovesi-Fonseca, C. Environmental effects and urban impacts on aquatic macroinvertebrates in a stream of central Brazilian Cerrado. Sustain. Water Resour. Manag. 2015, 1, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Valente-Neto, F.; Roque, F.D.O.; Rodrigues, M.E.; Juen, L.; Swan, C.M. Toward a practical use of Neotropical odonates as bioindicators: Testing congruence across taxonomic resolution and life stages. Ecol. Indic. 2016, 61, 952–959. [Google Scholar] [CrossRef]
- Carvalho, F.G.; Pinto, N.S.; Júnior, J.M.B.D.O.; Juen, L. Effects of marginal vegetation removal on Odonata communities. Acta Limnol. Bras. 2013, 25, 10–18. [Google Scholar] [CrossRef] [Green Version]
- IUCN 2020, The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org (accessed on 3 May 2020).
- Oliveira, L.F. (Ed.) Livro Vermelho da Fauna Brasileira Ameaçada de Extinção; ICMBio/MMA: Sao Paulo, Brazil, 2018. [Google Scholar]
- Oliveira-Junior, J.M.B.; Juen, L. The Zygoptera/Anisoptera Ratio (Insecta: Odonata): A New Tool for Habitat Alterations Assessment in Amazonian Streams. Neotrop. Entomol. 2019, 48, 552–560. [Google Scholar] [CrossRef]
- Bastos, R.C.; Brasil, L.S.; Oliveira-Junior, J.M.B.; Carvalho, F.G.; Lennox, G.D.; Barlow, J.; Juen, L. Morphological and phylogenetic factors structure the distribution of damselfly and dragonfly species (Odonata) along an environmental gradient in Amazonian streams. Ecol. Indic. 2021, 122. [Google Scholar] [CrossRef]
- De Marco, P., Jr.; Latini, A.O.; Resende, D.C. Thermoregulatory constraints on behavior: Patterns in a Neotropical dragonfly assemblage. Neotrop. Entomol. 2005, 34, 155–162. [Google Scholar] [CrossRef]
- Reis, E.F.; Pinto, N.S.; Carvalho, F.G.; Juen, L. Efeito da integridade ambiental sobre a Assimetria Flutuante em Erythrodiplax basalis (Libellulidae: Odonata)(Kirby). EntomoBrasilis 2011, 4, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Pereira, D.F.; de Oliveira, J.M., Jr.; Juen, L. Environmental changes promote larger species of Odonata (Insecta) in Amazonian streams. Ecol. Indic. 2019, 98, 179–192. [Google Scholar] [CrossRef]
- Yemane, D.; Field, J.G.; Leslie, R.W. Exploring the effects of fishing on fish assemblages using Abundance Biomass Comparison (ABC) curves. ICES J. Mar. Sci. 2005, 62, 374–379. [Google Scholar] [CrossRef] [Green Version]
- Warwick, R.M.; Clarke, K.R. Relearning the ABC: Taxonomic changes and abundance/biomass relationships in disturbed benthic communities. Mar. Biol. 1994, 118, 739–744. [Google Scholar] [CrossRef]
- De Marco, P., Jr.; Batista, J.D.; Cabette, H.S.R. Community Assembly of Adult Odonates in Tropical Streams: An Ecophysiological Hypothesis. PLoS ONE 2015, 10, e0123023. [Google Scholar] [CrossRef] [Green Version]
- Juen, L.; De Oliveira-Junior, J.M.B.; Shimano, Y.; Mendes, T.P.; Cabette, H.S.R. Composição e riqueza de Odonata (Insecta) em riachos com diferentes níveis de conservação em um ecótone Cerrado-Floresta Amazônica. Acta Amaz. 2014, 44, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Dutra, S.; De Marco, P. Bionomic differences in odonates and their influence on the efficiency of indicator species of environmental quality. Ecol. Indic. 2015, 49, 132–142. [Google Scholar] [CrossRef]
- Calvão, L.B.; Juen, L.; de Oliveira, J.M.B., Jr.; Batista, J.D.; De Marco Júnior, P. Land use modifies Odonata diversity in streams of the Brazilian Cerrado. J. Insect Conserv. 2018, 22, 675–685. [Google Scholar] [CrossRef]
- Nogueira, D.S.; Montag, L.; Nogueira, D.S.; Calvão, L.B.; Fogaça, L.; Montag, D.A. Little effects of reduced-impact logging on insect communities in eastern Amazonia Little effects of reduced-impact logging on insect communities. Environ. Monit. Assess. 2016. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; De Marco, P.; Dias-silva, K.; Pereira, R.; Gontijo, C.; Santos, P.; Allan, T.; Hughes, R.M.; Juen, L. Limnologica Effects of human disturbance and riparian conditions on Odonata (Insecta) assemblages in eastern Amazon basin streams. Limnologica 2017, 66, 31–39. [Google Scholar] [CrossRef]
- Embrapa—Empresa Brasileira de Pesquisa Agropecuária, Sistema Brasileiro de Classificação de Solos. Available online: https://www.embrapa.br/solos/sibcs (accessed on 29 August 2020).
- Nessimian, J.L.; Venticinque, E.M.; Zuanon, J.; De Marco, P.; Gordo, M.; Fidelis, L.; D’arc Batista, J.; Juen, L. Land use, habitat integrity, and aquatic insect assemblages in Central Amazonian streams. Hydrobiologia 2008, 614, 117–131. [Google Scholar] [CrossRef]
- Cezário, R.R.; Firme, P.P.; Pestana, G.C.; Vilela, D.S.; Juen, L.; Cordero-Rivera, A.; Guillermo, R. Sampling Methods for Dragonflies and Damselflies. Meas. Arthropod Biodivers. 2020, 2020, 223–240. [Google Scholar]
- Mccauley, S.J.; Davis, C.J.; Relyea, R.A.; Yurewicz, K.L.; Skelly, D.K.; Werner, E.E. Metacommunity patterns in larval odonates. Oecologia 2008, 158, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lencioni, F.A.A. Damselflies of Brazil: An Illustrated Identification Guide: II-Coenagrionidae Families; NHBS: São Paulo, Brazil, 2006. [Google Scholar]
- Belle, J.A. synopsis of the species of Phyllocycla Calvert, with descriptions of four new taxa and a key to the genera of neotropical Gomphidae (Odonata, Gomphidae). Tijdschr. Voor Entomol. 1988, 131, 73–102. [Google Scholar]
- Belle, J. Higher classification of the South-American Gomphidae (Odonata). Zool. Meded. 1996, 70, 297–324. [Google Scholar]
- Garrison, R. A Synopsis of the Genera Mnerasete Cowley, Bryoplathanon Gen. Nov., and Ormenoplebia Gen. Nov. (Odonata: Calopterygidae). Geography 2006, 2006, 90421531. [Google Scholar]
- Pessacq, P. Synopsis of Epipleoneura (Zygoptera, Coenagrionidae, “Protoneuridae”), with emphasis on its Brazilian species. Zootaxa 2014, 3872, 201. [Google Scholar] [CrossRef] [Green Version]
- Heckman, C.W. Encyclopedia of South American Aquatic Insects: Odonata—Anisoptera: Illustrated Keys to Known Families, Genera, and Species in South America; Springer: Berlin, Germany, 2006. [Google Scholar]
- Mendes, T.P.; Oliveira-Junior, J.M.B.; Cabette, H.S.R.; Batista, J.D.; Juen, L. Congruence and the Biomonitoring of Aquatic Ecosystems: Are Odonate Larvae or Adults the Most Effective for the Evaluation of Impacts. Neotrop. Entomol. 2017, 46, 631–641. [Google Scholar] [CrossRef]
- Brasil, L.S.; Luiza-Andrade, A.; Calvão, L.B.; Dias-Silva, K.; Faria, A.P.J.; Shimano, Y.; Oliveira-Junior, J.M.B.; Cardoso, M.N.; Juen, L. Aquatic insects and their environmental predictors: A scientometric study focused on environmental monitoring in lotic environmental. Environ. Monit. Assess. 2020, 192. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolation, extrapolation, and comparingincidence-based species accumulation curves. Ecology 2004, 85, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chiu, C.H. Nonparametric Estimation and Comparison of Species Richness. eLS 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Pearson: London, UK, 2010. [Google Scholar]
- Carvalho, F.G.; Oliveira-Junior, J.M.B.; Farias, A.P.J.; Juen, L. Use of the ABC curve as a method to detect the effect of anthropogenic change on the Odonata (Insecta) assembly. Interciencia 2013, 38, 516–522. [Google Scholar]
- Oksanen, J.; Blanchett, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Vegan: Community Ecology Package. R Package 2.0.3. Available online: http//CRAN.R-project.org/package=vega2013 (accessed on 29 August 2020).
- Alves-Martins, F.; Brasil, L.S.; Juen, L.; De Marco, P.; Stropp, J.; Hortal, J. Metacommunity patterns of Amazonian Odonata: The role of environmental gradients and major rivers. PeerJ 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Heino, J. Environmental heterogeneity, dispersal mode, and co-occurrence in stream macroinvertebrates. Ecol. Evol. 2013, 3, 344–355. [Google Scholar] [CrossRef]
- Balvanera, P.; Pfisterer, A.B.; Buchmann, N.; He, J.S.; Nakashizuka, T.; Raffaelli, D.; Schmid, B. Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol. Lett. 2006, 9, 1146–1156. [Google Scholar] [CrossRef] [Green Version]
- Nagy, L.; Williams, W.; Tauxe, L.; Muxworthy, A.R.; Ferreira, I. Thermomagnetic recording fidelity of nanometer-sized iron and implications for planetary magnetism. Proc. Natl. Acad. Sci. USA 2019, 116, 1984–1991. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J.; French, G.C.; Harvey, I.F. Historical changes in the phenology of British Odonata are related to climate. Glob. Chang. Biol. 2007, 13, 933–941. [Google Scholar] [CrossRef]
- Gonçalves, C.S.; Braga, F. Diversidade de peixes na área de influência da UHE Mogi Guaçu e lagoas marginais, bacia do alto Rio Paraná, São Paulo, Brasil. Biota Neotrop. 2008, 2, 103–114. [Google Scholar] [CrossRef] [Green Version]
- De Marco, P., Jr.; Resende, D.C. Activity patterns and thermoregulation in a tropical dragonfly assemblage. Odonatologica 2002, 31, 129–138. [Google Scholar]
- Chang, X.; Zhai, B.; Wang, M.; Wang, B. Relationship between exposure to an insecticide and fluctuating asymmetry in a damselfly (Odonata, Coenagrionidae). Hydrobiologia 2007, 586, 213–220. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M.H. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar] [CrossRef]
- Fulan, J.Â.; Henry, R. Distribuição temporal de imaturos de Odonata (Insecta) associados Eichhornia azurea (Kunth) na Lagoa do Camargo, Rio Paranapanema, São Paulo. Rev. Bras. Entomol. 2007, 51, 224–227. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.A.; Barlow, J.; Chazdon, R.; Ewers, R.M.; Harvey, C.A.; Peres, C.A.; Sodhi, N.S. Prospects for tropical forest biodiversity in a human-modified world. Ecol. Lett. 2009, 12, 561–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]


| Number of Specimens by Treatment | ||||||
|---|---|---|---|---|---|---|
| Suborder | Family | Species | Altered | Control | Intermediare | Total Specimens |
| Zygoptera | Calopterygidae | Hetaerina sanguinea Selys, 1853 | 1 | 7 | 6 | 14 |
| Hetaerina sp. | – | 1 | 4 | 5 | ||
| Mnesarete aenea (Selys, 1853) | – | 3 | – | 3 | ||
| Mnesarete smaragdina (Selys, 1869) | – | 4 | 1 | 5 | ||
| Mnesarete sp. | – | 5 | 1 | 6 | ||
| Mnesarete williamsoni Garrison, 2006 | – | 10 | – | 10 | ||
| Coenagrionidae | Acanthagrion aepiolum Tennessen, 2004 | 26 | – | – | 26 | |
| Acanthagrion amazonicum Sjöstedt, 1918 | 9 | – | – | 9 | ||
| Acanthagrion kennedii Williamson, 1916 | 14 | 4 | 6 | 24 | ||
| Acanthagrion rubrifrons Leonard, 1977 | 3 | 2 | – | 3 | ||
| Acanthagrion sp. | 4 | – | – | 4 | ||
| Aeolagrion dorsale (Burmeister, 1839) | 1 | 2 | 6 | 9 | ||
| Argia fumigata Hagen in Selys, 1865 | – | 16 | 5 | 21 | ||
| Argia indicatrix Calvert, 1902 | 21 | – | 4 | 25 | ||
| Argia oculata Hagen in Selys, 1865 | – | 7 | 3 | 10 | ||
| Argia sp. | 2 | 3 | 1 | 6 | ||
| Argia sp.1 | – | 1 | - | 1 | ||
| Argia thespis Hagen in Selys, 1865 | 3 | 2 | 1 | 6 | ||
| Argia tinctipennis Selys, 1865 | – | 5 | 4 | 9 | ||
| Ischnura capreolus (Hagen, 1861) | 5 | – | – | 5 | ||
| Ischnura sp. | 9 | – | – | 9 | ||
| Dicteriadidae | Heliocharis amazona Selys, 1853 | - | 1 | – | 1 | |
| Lestidae | Lestes sp. | 1 | – | 1 | 2 | |
| Megapodagrionidae | Heteragrion icterops Selys, 1862 | – | 1 | – | 1 | |
| Oxystigma williamsoni Geijskes, 1976 | – | 3 | – | 3 | ||
| Polythoridae | Chalcopteryx radians Ris, 1914 | – | – | 3 | 3 | |
| Chalcopteryx rutilans (Rambur, 1842) | – | 3 | – | 3 | ||
| Chalcopteryx sp. | – | 1 | – | 1 | ||
| Protoneuridae | Epipleoneura kaxuriana Machado, 1985 | – | 4 | 3 | 7 | |
| Epipleoneura lamina Williamson, 1915 | – | – | 8 | 8 | ||
| Epipleoneura metallica Rácenis, 1955 | – | 21 | 11 | 32 | ||
| Epipleoneura solitaria De Marmels, 1989 | – | 8 | 1 | 9 | ||
| Epipleoneura sp. | 1 | 1 | 2 | 4 | ||
| Epipleoneura spatulata Rácenis, 1960 | – | – | 5 | 5 | ||
| Neoneura fulvicollis Selys, 1886 | – | – | 1 | 1 | ||
| Neoneura gaida Racenis, 1953 | 1 | – | 6 | 7 | ||
| Neoneura luzmarina De Marmels, 1989 | – | 11 | 16 | 27 | ||
| Neoneura sp. | – | – | 2 | 2 | ||
| Neoneura rubriventris Selys, 1860 | – | 1 | 1 | 2 | ||
| Protoneura scintilla Gloyd, 1939 | 1 | – | – | 1 | ||
| Protoneura sp. | 1 | 1 | 2 | 4 | ||
| Protoneura tenuis Selys, 1860 | – | 4 | 3 | 7 | ||
| Total specimens | 103 | 130 | 107 | 340 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brito, J.P.; Carvalho, F.G.; Juen, L. Response of the Zygopteran Community (Odonata: Insecta) to Change in Environmental Integrity Driven by Urbanization in Eastern Amazonian Streams. Ecologies 2021, 2, 150-163. https://doi.org/10.3390/ecologies2010008
Brito JP, Carvalho FG, Juen L. Response of the Zygopteran Community (Odonata: Insecta) to Change in Environmental Integrity Driven by Urbanization in Eastern Amazonian Streams. Ecologies. 2021; 2(1):150-163. https://doi.org/10.3390/ecologies2010008
Chicago/Turabian StyleBrito, Jhose Paixão, Fernando Geraldo Carvalho, and Leandro Juen. 2021. "Response of the Zygopteran Community (Odonata: Insecta) to Change in Environmental Integrity Driven by Urbanization in Eastern Amazonian Streams" Ecologies 2, no. 1: 150-163. https://doi.org/10.3390/ecologies2010008
APA StyleBrito, J. P., Carvalho, F. G., & Juen, L. (2021). Response of the Zygopteran Community (Odonata: Insecta) to Change in Environmental Integrity Driven by Urbanization in Eastern Amazonian Streams. Ecologies, 2(1), 150-163. https://doi.org/10.3390/ecologies2010008