
Species-Abundance Models for the Early Postfire Succession of Subalpine Shrub Grassland


Abstract
:1. Introduction
2. Materials and Methods
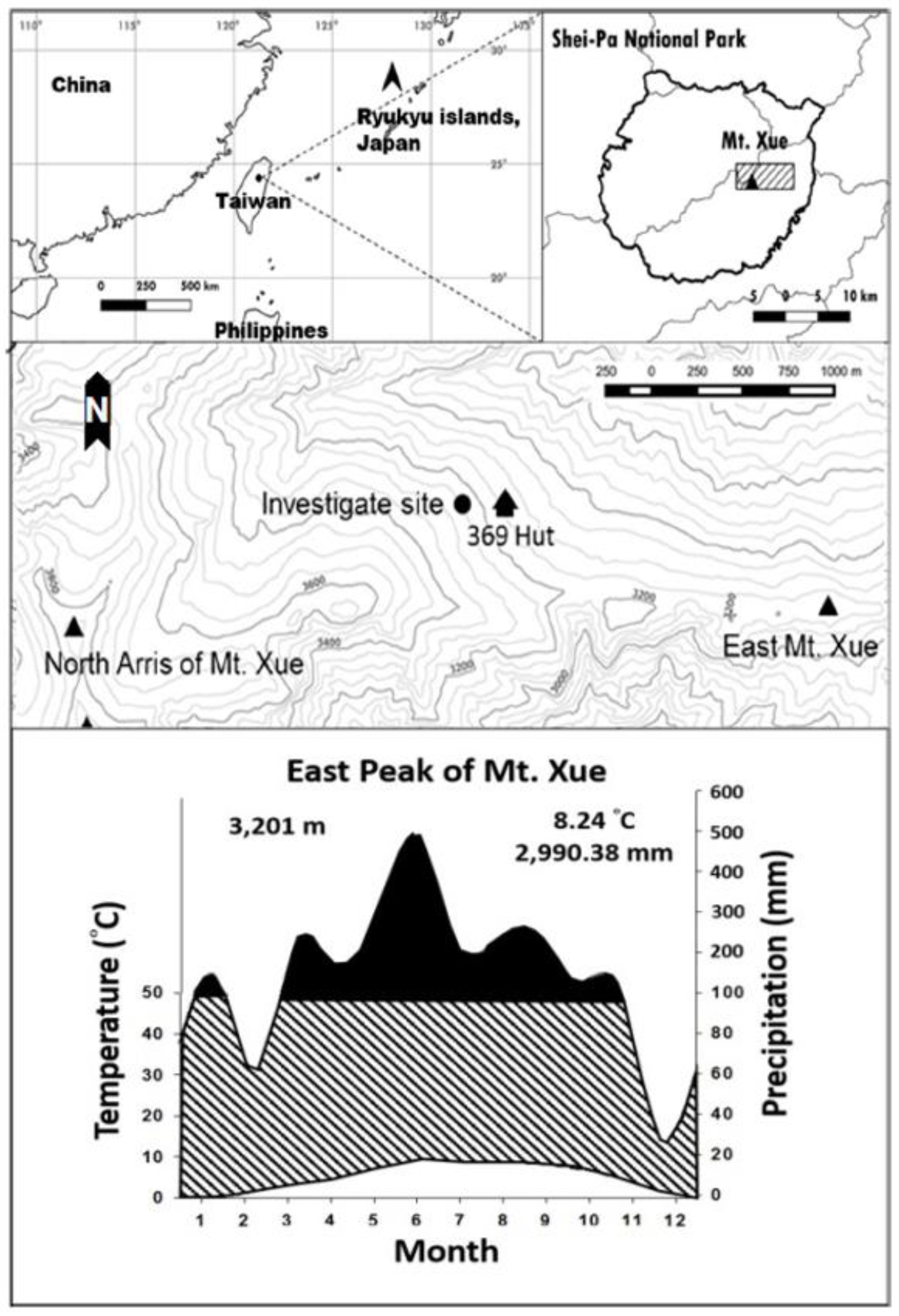
2.1. Study Area
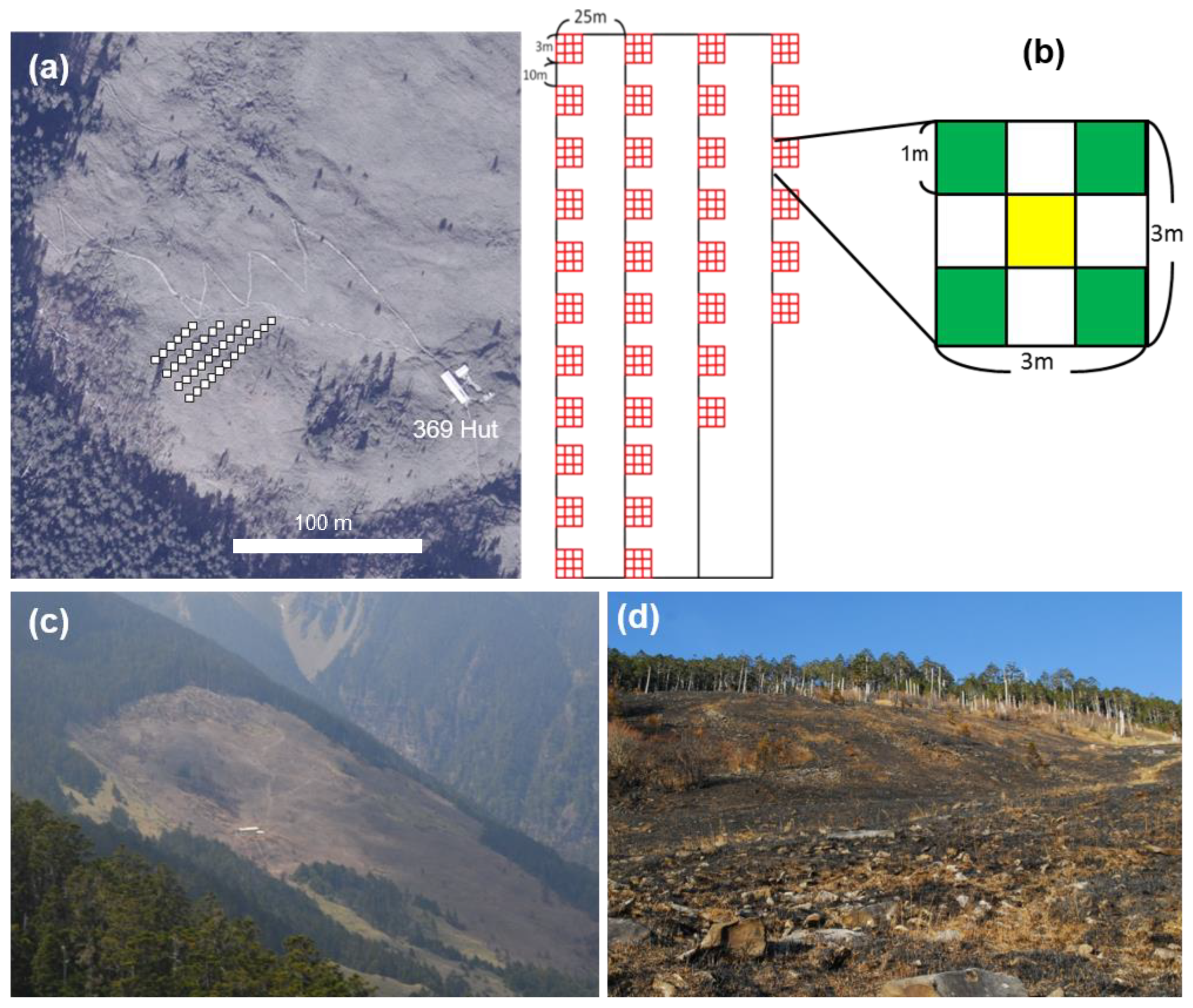
2.2. Sampling and Monitoring
2.3. Data Analysis
2.3.1. Species Diversity during Postfire Succession
2.3.2. Log-Normal Distribution of Species-Coverage Relationships
2.3.3. Species-Abundance Model Simulation for Postfire Subalpine Shrub Grasslands
- (1)
- Preemption Model
- (2)
- Broken-stick model
- (3)
- Log-normal model
- (4)
- Zipf model
- (5)
- Zipf–Mandelbrot Model
3. Results
3.1. Short-Term Change of Postfire Species Diversity
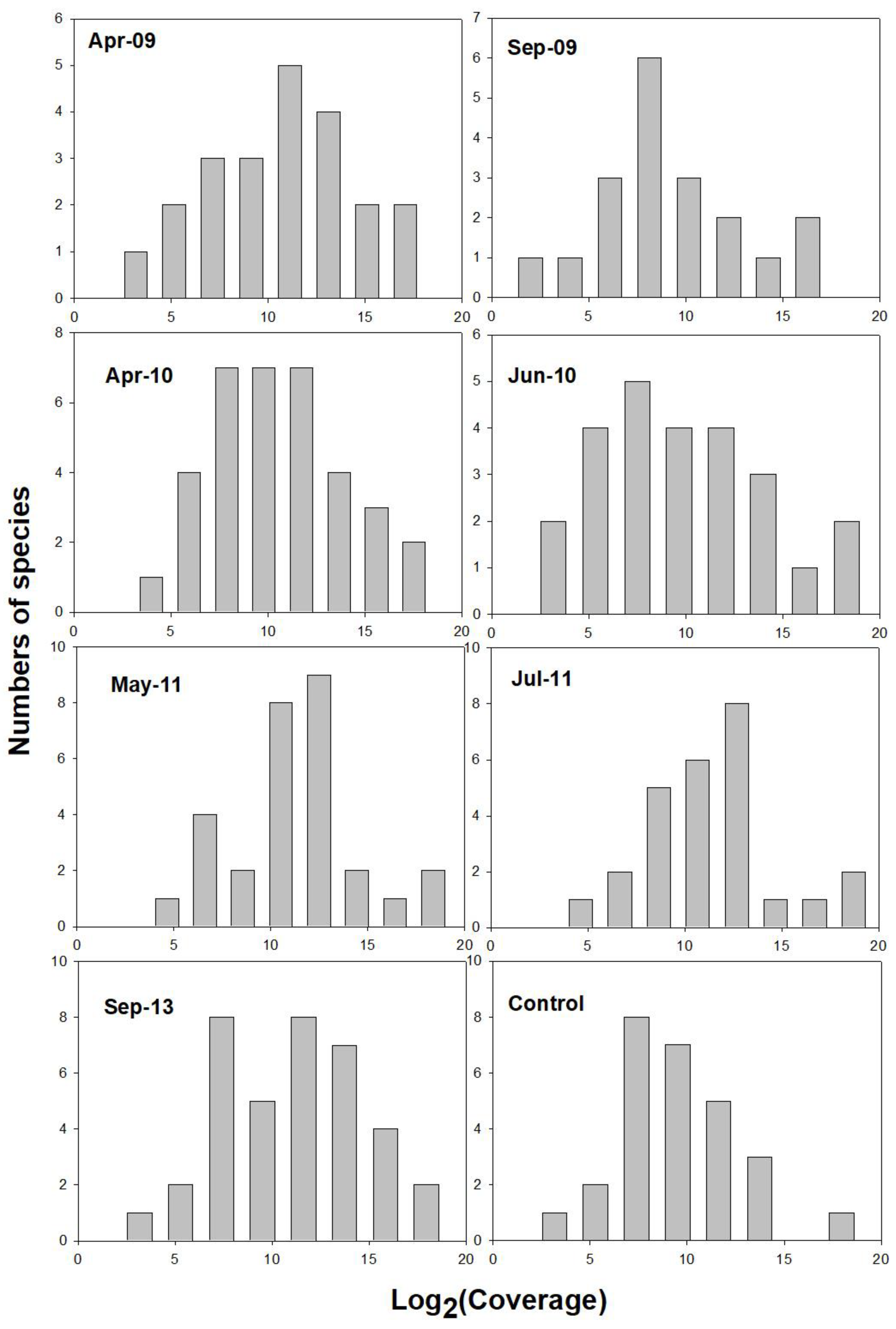
3.2. Species–Abundance Analysis of Postfire Plant Coverage
3.3. Species-Abundance Model Simulation for Postfire Subalpine Shrub Grasslands
4. Discussion
4.1. Short-Term Change of Postfire Species Diversity
4.2. Species-Abundance Series Analysis of Postfire Plant Coverage
4.3. Species-Abundance Model Simulation for Postfire Subalpine Shrub Grasslands
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fire Periods | Reference | Cause |
|---|---|---|
| 1903~1957 | [51] | unknown |
| 1957~1958 | ||
| 2008 | http://news.ltn.com.tw/news/life/paper/267022 (accessed on 16 December 2023) | artificial |
| 2014 | https://news.ltn.com.tw/news/society/paper/748572 (accessed on 16 December 2023) | artificial |
| 2019 | https://news.ltn.com.tw/news/life/breakingnews/2692421 (accessed on 16 December 2023) | artificial |
References
- Coop, J.D.; Massatti, R.T.; Schoettle, A.W. Subalpine vegetation pattern three decades after stand-replacing fire: Effects of landscape context and topography on plant community composition, tree regeneration, and diversity. J. Veg. Sci. 2010, 21, 472–487. [Google Scholar] [CrossRef]
- Turner, M.G.; Braziunas, K.H.; Hansen, W.D.; Harvey, B.J. Short-interval severe fire erodes the resilience of subalpine lodgepole pine forests. Proc. Natl. Acad. Sci. USA 2019, 116, 11319–11328. [Google Scholar] [CrossRef] [PubMed]
- Bashirzadeh, M.; Abedi, M.; Shefferson, R.P.; Farzam, M. Post-Fire Recovery of Plant Biodiversity Changes Depending on Time Intervals since Last Fire in Semiarid Shrublands. Fire 2023, 6, 103. [Google Scholar] [CrossRef]
- Tilman, D. Resource Competition and Community Structure. In Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1982; Volume 17, pp. 1–296. [Google Scholar]
- Lu, K.C. Effects of Wildfires on the Main Forest Ecosystems in Taiwan. Ph.D. Thesis, National Chung-Hsing University, Taichung, Taiwan, 1989. [Google Scholar]
- Lai, G.S. Dynamic Structures of Ecotones between Subalpine Coniferous forests and Grasslands in Taiwan. Ph.D. Thesis, National Chung-Hsing University, Taichung, Taiwan, 1992. [Google Scholar]
- Lai, G.S. The fire ecology of subalpine in Taiwan. In Seminar of Fire Ecology and Management; Forestry Bureau: Taipei, Taiwan, 2003; pp. 49–52. [Google Scholar]
- Overbeck, G.E.; Müller, S.C.; Pillar, V.D.; Pfadenhauer, J. Small-scale dynamics after fire in South Brazilian humid subtropical grassland. J. Veg. Sci. 2005, 16, 655–664. [Google Scholar] [CrossRef]
- Wang, W.; Tsai, S.T.; Chiu, C.A.; Hsu, C.K.; Tzeng, H.Y.; Lu, K.C. Species and life-form diversity along the altitudinal gradient on the Mt. Shei Eastern Trail. Q. J. For. Res. 2013, 5, 139–152. [Google Scholar]
- Su, H.J. Studies on the climate and vegetation types of the natural forests in Taiwan. (II). Altitudinal vegetation zones in relation to temperature gradient. Q. J. Chin. For. 1984, 17, 57–73. [Google Scholar]
- Wang, W.; Chiu, C.A.; Tsai, S.T.; Hsu, C.K.; Tzeng, H.Y.; Lu, K.C. Vegetation research along the East Trail of Mt. Shei. Q. J. For. Res. 2011, 32, 15–34. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Lu, F.Y.; Ou, C.H.; Lai, G.S. Plant succession and competition mechanism of Yushania niitakayamensis at alpine in Taiwan. Q. J. Chin. For. 1984, 17, 1–32. [Google Scholar]
- McGill, B.J.; Etienne, R.S.; Gray, J.S.; Alonso, D.; Anderson, M.J.; Benecha, H.K.; Dornelas, M.; Enquist, B.J.; Green, J.L.; He, F.; et al. Species abundance distributions: Moving beyond single prediction theories to integration within an ecological framework. Eco. Lett. 2007, 10, 995–1015. [Google Scholar] [CrossRef]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- MacArthur, R.H. On the relative abundance of bird species. Proc. Natl. Acad. Sci. USA 1957, 43, 293–295. [Google Scholar] [CrossRef]
- Zhang, J.T. Quantitative Ecology; Science Press: Beijing, China, 2004; pp. 1–370. [Google Scholar]
- Yin, Z.Y.; Ren, H.; Peng, S.L.; Guo, Q.F.; Zeng, L.H.; He, X. Dynamics and modeling of species abundance distribution during natural restoration of degraded hilly grassland in south China. Ecol. Environ. Sci. 2009, 1, 222–228. [Google Scholar] [CrossRef]
- Li, Q.; Tu, J.; Xiong, Z.P.; Lu, Z.X.; Liu, C.J. Survey of research on species abundance pattern. J. Yunnan Agric. Univ. 2011, 26, 117–123. [Google Scholar]
- Tsai, S.T.; Lu, K.C.; Tzeng, H.Y. Analyzing vegetation dynamics of the broad-leaved secondary forest by species abundance models at Mt. Showchu in the Hue-Sun Forest Station. Q. J. For. Res. 2011, 33, 9–22. [Google Scholar] [CrossRef]
- Huang, L.; Yang, H.; An, X.; Yu, Y.; Yu, L.; Huang, G.; Liu, X.; Chen, M.; Xue, Y. Species Abundance Distributions Patterns between Tiankeng Forests and Nearby Non-Tiankeng Forests in Southwest China. Diversity 2022, 14, 64. [Google Scholar] [CrossRef]
- Zang, Z.; Zeng, Y.; Wang, D.; Shi, F.; Dong, Y.; Liu, N.; Liang, Y. Species-Abundance Distribution Patterns of Plant Communities in the Gurbantünggüt Desert, China. Sustainability 2022, 14, 12957. [Google Scholar] [CrossRef]
- Antão, L.H.; Magurran, A.E.; Dornelas, M. The Shape of Species Abundance Distributions Across Spatial Scales. Front. Ecol. Evol. 2021, 9, 626730. [Google Scholar] [CrossRef]
- Cheng, J.J.; Mi, X.C.; Ma, K.P.; Zhang, J.T. Responses of species-abundance distribution to varying sampling scales in a subtropical broad-leaved forest. Biodivers. Sci. 2011, 19, 168–177. [Google Scholar] [CrossRef]
- Wilson, J.B.; Wells, T.C.E.; Trueman, C.; Jones, G.; Atkinson, M.D.; Crawley, M.J.; Dodd, M.E.; Silvertown, J. Are there assembly rules for plant species abundance: An investigation in relation to soil resources and successional trends. J. Eco. 1996, 84, 527–538. [Google Scholar] [CrossRef]
- Central Weather Bureau. 2022. Available online: https://www.cwb.gov.tw/V8/C/ (accessed on 31 December 2022).
- Ho, C.S. Geological Introduction of Taiwan; Central Geological Survey (MOEA): Taipei, Taiwan, 2003; pp. 1–163. [Google Scholar]
- Yen, C.H. Soil properties of alpine in Xue Mountain. In Alpine Ecosystem Integrative Survey in Xue Mountain; Lu, K.C., Ou, C.H., Eds.; Shei-Pa National Park Headquarters: Miaoli, Taiwan, 2009; Chapter 3; pp. 1–30. [Google Scholar]
- Cheng, T.W.; Tzeng, H.Y.; Chiu, C.A.; Liu, S.Q.; Wang, C.M.; Tseng, Y.H. Studies on the life form of vascular plants along the East Trail of Xue Mountain. J. Nat. Park 2012, 22, 41–51. [Google Scholar]
- Wu, C.Y.; Tzeng, H.Y.; Qiu, Q.A.; Wang, C.M.; Liu, S.C.; Tseng, Y.H. Flowering phenology of East Xue Trail of Xue Mountain. Q. J. For. Res. 2013, 35, 223–240. [Google Scholar]
- Wilson, J.B. Methods for fitting dominance/diversity curves. J. Veg. Sci. 1991, 2, 35–46. [Google Scholar] [CrossRef]
- Tang, Z.Y.; Fang, J.Y. A review on the elevational patterns of plant species diversity. Sheng Wu Duo Yang Xing 2004, 12, 20–28. [Google Scholar] [CrossRef]
- Liu, T.S.; Su, H.J. Forest Ecology; The Commercial Press: Taipei, Taiwan, 1983; pp. 1–462. [Google Scholar]
- Midgley, J.J. Why the world’s vegetation is not totally dominated by resprouting plants; because resprouters are shorter than reseeders. Ecography 1996, 19, 92–95. Available online: https://www.jstor.org/stable/3683094 (accessed on 10 October 2023). [CrossRef]
- Hughes, R.G. Theories and models of species abundance. Am. Nat. 1986, 128, 879–899. Available online: https://www.jstor.org/stable/2461769 (accessed on 10 October 2023). [CrossRef]
- Motomura, I. A statistical treatment of associations. Jpn. J. Zool. 1932, 44, 379–383. Available online: https://cir.nii.ac.jp/crid/1570009750645470336 (accessed on 10 October 2023).
- Preston, F.W. The commonness and rarity of species. Ecology 1948, 29, 254–283. [Google Scholar] [CrossRef]
- May, R.M. Patterns of species abundance and diversity. In Ecology and Evolution of Communities; Lody, M.L., Diamond, J.M., Eds.; Harvard University Press: Cambridge, MA, USA, 1975; pp. 81–120. [Google Scholar]
- Barange, M.; Campos, B. Models of species abundance: A critique of and an alternative to the dynamics model. Mar. Ecol. Prog. Ser. 1991, 69, 293–298. Available online: https://digital.csic.es/bitstream/10261/281973/1/Barange_et_al_1991.pdf (accessed on 10 October 2023). [CrossRef]
- Frontier, S. Diversity and structure in aquatic ecosystems. Oceanogr. Mar. Biol. 1985, 23, 253–312. [Google Scholar]
- Moradizadeh, H.; Heydari, M.; Omidipour, R.; Mezbani, A.; Prévosto, B. Ecological effects of fire severity and time since fire on the diversity partitioning, composition and niche apportionment models of post-fire understory vegetation in semi-arid oak forests of Western Iran. Ecol. Eng. 2020, 143, 105694. [Google Scholar] [CrossRef]
- Bell, D.T. Ecological response syndromes in the flora of southwestern Western Australia: Fire resprouters versus reseeders. Bot. Rev. 2001, 67, 417–440. [Google Scholar] [CrossRef]
- Sritharan, M.S.; Scheele, B.C.; Blanchard, W.; Foster, C.N.; Werner, P.A.; Lindenmayer, D.B. Plant rarity in fire-prone dry sclerophyll communities. Sci. Rep. 2022, 12, 12055. [Google Scholar] [CrossRef] [PubMed]
- Vogl, R.J. Effects of fire on grasslands. In Fire and Ecosystems; Kozlowski, T.T., Ahlgren, C.E., Eds.; Academic: New York, NY, USA, 1974; pp. 139–194. [Google Scholar]
- Connell, J.H. Diversity in tropical rain forest and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Huston, M.A. A general hypothesis of species diversity. Am. Nat. 1979, 113, 81–101. Available online: https://www.jstor.org/stable/2459944 (accessed on 10 October 2023). [CrossRef]
- Denslow, J.S. Gap partitioning among tropical rainforest trees. Biotropica 1980, 12, 47–55. [Google Scholar] [CrossRef]
- Lin, C.C.; Chiou, C.R.; Chou, C.Y. Identifying and evaluating fire severity: A case study of the Wulin fire. Taiwan J. For. Sci. 2014, 20, 203–213. [Google Scholar] [CrossRef]
- Pan, C.C.; Tseng, Y.H.; Chiu, C.A.; Tzeng, H.Y. Phenology of Rhododendron pseudochrysanthum in Mt. Xue. Q. J. For. Res. 2013, 35, 71–86. [Google Scholar]
- Deng, N.; Song, Q.; Ma, F.; Tian, Y. Patterns and Driving Factors of Diversity in the Shrub Community in Central and Southern China. Forests 2022, 13, 1090. [Google Scholar] [CrossRef]
- Chiarucci, A.; Wilson, J.B.; Anderson, B.J.; De Dominicis, V. Cover versus biomass as an estimate of species abundance: Does it make a difference to the conclusions? J. Veg. Sci. 1999, 10, 35–42. [Google Scholar] [CrossRef]
- Chen, M.Y.; Shih, Y.Y. Studies on the effects of wildfires on the vegetation at Fansan and Shesan areas. Nat. Park J. 1999, 8, 155–165. [Google Scholar]

| Parameters | Survey Period | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Feb-09 | Apr-09 | Sep-09 | Apr-10 | Jun-10 | May-11 | Jul-11 | Sep-13 | Control | |
| Season 1 | 1 | 1 | 2 | 1 | 2 | 1 | 2 | 2 | 2 |
| Number of species | 5 | 13 | 24 | 24 | 36 | 27 | 33 | 29 | 27 |
| Coverage (%) | 0.21 | 0.93 | 25.91 | 13.14 | 43.95 | 42.47 | 48.45 | 64.80 | 100.00 |
| Phanerophytes (%) | 0.00 | 0.00 | 0.00 | 5.26 | 3.23 | 4.17 | 6.90 | 8.33 | 14.29 |
| Chamaephytes (%) | 0.00 | 18.18 | 22.73 | 26.32 | 16.13 | 20.83 | 17.24 | 8.33 | 9.52 |
| Hemicryptophytes (%) | 75.00 | 54.55 | 54.55 | 52.63 | 61.29 | 54.17 | 58.62 | 66.67 | 57.14 |
| Geophytes (%) | 25.00 | 27.27 | 22.73 | 15.79 | 19.35 | 20.83 | 17.24 | 16.67 | 19.05 |
| Survival strategy 2 | 100/0 | 92/8 | 79/21 | 81/19 | 62/38 | 70/30 | 63/37 | 61/39 | 41/59 |
| Ferns quotient | 6.25 | 4.55 | 2.27 | 2.63 | 2.42 | 3.13 | 2.59 | 4.17 | 5.71 |
| Gleason index | 1.01 | 2.62 | 4.83 | 4.83 | 7.24 | 5.43 | 6.64 | 5.84 | 7.32 |
| Shannon index | 0.44 | 0.57 | 1.10 | 0.96 | 1.11 | 0.87 | 1.08 | 1.03 | 1.08 |
| Evenness index | 0.27 | 0.22 | 0.35 | 0.30 | 0.31 | 0.26 | 0.31 | 0.31 | 0.33 |
| Survey Period | S 1 | Species-Abundance Models | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Broken Stick | Preemption | Log-Normal | Zipf | Zipf–Mandelbrot | |||||||
| Dev. | χ2 | Dev. | χ2 | Dev. | χ2 | Dev. | χ2 | Dev. | χ2 | ||
| Feb-09 | 1 | 0.29 | 0.25 | 0.82 | 2.04 | 0.01 | 0.01 | 0.00 * | 0.00 | 0.00 | 0.01 |
| Apr-09 | 1 | 1.15 | 1.27 | 2.42 | 4.48 | 0.03 * | 0.03 | 0.03 | 0.02 | 0.04 | 0.04 |
| Apr-10 | 1 | 25.33 | 26.39 | 36.14 | 48.65 2 | 6.11 | 7.58 | 7.04 | 8.68 | 4.82 * | 5.72 |
| May-11 | 1 | 111.30 | 124.90 | 135.84 | 186.24 | 12.60 | 15.03 | 13.99 | 16.50 | 8.59 * | 9.68 |
| Sep-09 | 2 | 30.67 | 30.36 | 44.70 | 54.26 | 4.22 | 4.50 | 7.12 | 6.87 | 3.84 * | 3.64 |
| Jun-10 | 2 | 66.53 | 67.15 | 64.37 | 69.46 | 11.22 | 11.64 | 17.01 | 16.94 | 9.47 * | 9.29 |
| Jul-11 | 2 | 104.47 | 25.62 | 115.11 | 24.23 | 6.70 | 0.25 | 7.98 | 0.93 | 4.49 * | 0.49 |
| Sep-13 | 2 | 157.85 | 68.51 | 184.94 | 71.86 | 25.43 | 1.94 | 27.32 | 6.40 | 18.12 * | 3.15 |
| Control | 2 | 90.03 | 143.08 | 107.12 | 217.77 | 4.07 | 15.38 | 3.29 * | 3.61 | 5.83 | 10.19 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Liao, M.-C.; Tzeng, H.-Y. Species-Abundance Models for the Early Postfire Succession of Subalpine Shrub Grassland. Fire 2024, 7, 21. https://doi.org/10.3390/fire7010021
Wang W, Liao M-C, Tzeng H-Y. Species-Abundance Models for the Early Postfire Succession of Subalpine Shrub Grassland. Fire. 2024; 7(1):21. https://doi.org/10.3390/fire7010021
Chicago/Turabian StyleWang, Wei, Min-Chun Liao, and Hsy-Yu Tzeng. 2024. "Species-Abundance Models for the Early Postfire Succession of Subalpine Shrub Grassland" Fire 7, no. 1: 21. https://doi.org/10.3390/fire7010021