Pyro-Ecophysiology: Shifting the Paradigm of Live Wildland Fuel Research


{kind=link}
Abstract
:1. Introduction
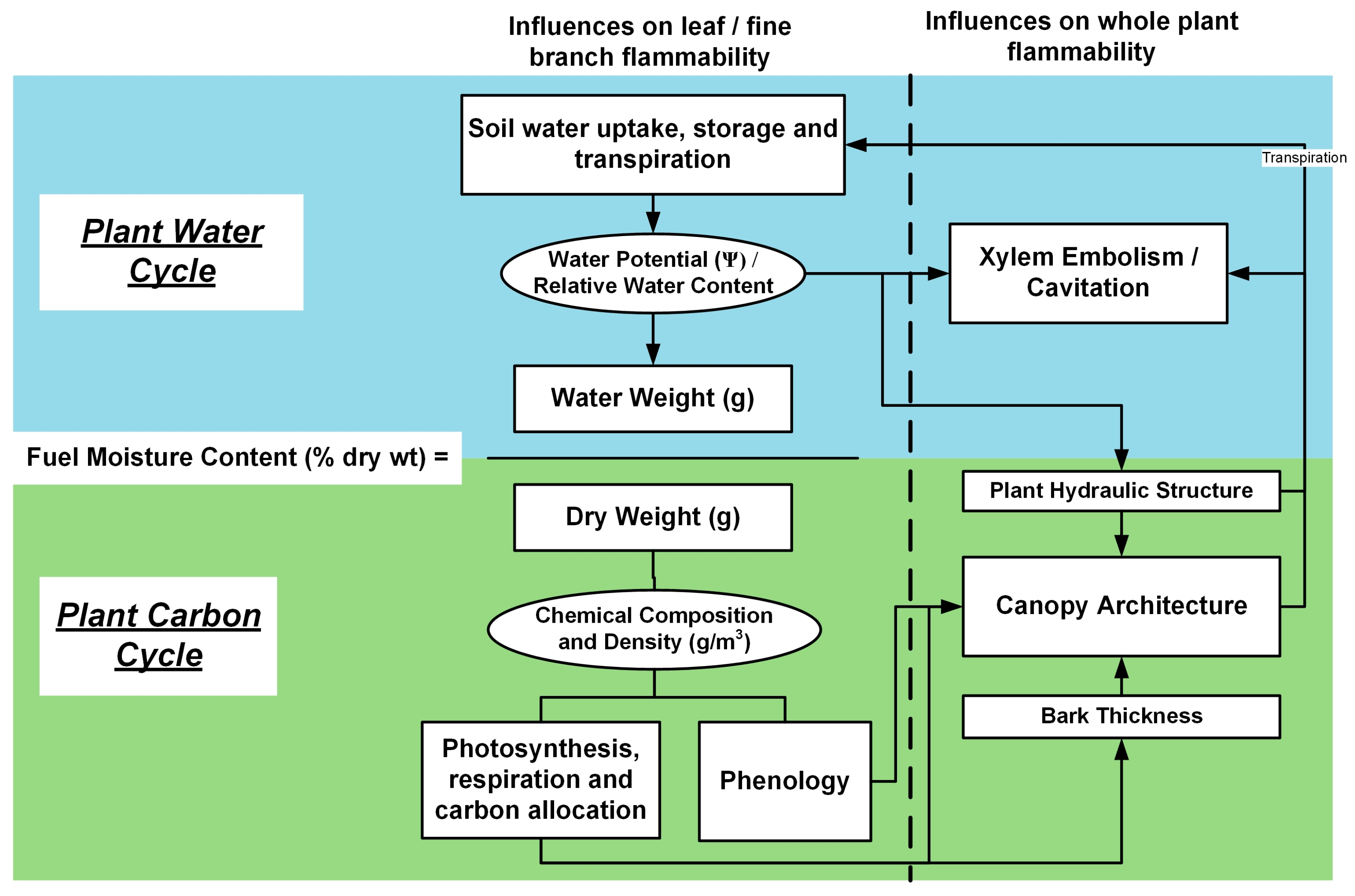
1.1. Leaf-Level Linkages between Physiology and Flammability
1.2. Whole-Plant Level Linkages between Physiology and Flammability
2. Summary: On the Need for a New Fire Science Discipline: Pyro-Ecophysiology
Author Contributions
Conflicts of Interest
References
- Gisborne, H.T. Measuring Forest-Fire Danger in Northern Idaho; US Dept. of Agriculture: Washington, DC, USA, 1928.
- Fons, W.L. Analysis of fire spread in light forest fuels. J. Agric. Res. 1946, 72, 93–121. [Google Scholar]
- Anderson, H.E.; Rothermel, R.C. Influence of moisture and wind upon the characteristics of free-burning fires. Symp. (Int.) Combust. 1965, 10, 1009–1019. [Google Scholar] [CrossRef]
- Rothermel, R.C. A Mathematical Model for Predicting Fire Spread in Wildland Fuels; USDA Forests Service Research Paper; Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1972. [Google Scholar]
- Dimitrakopoulos, A.P.; Bemmerzouk, A.M. Predicting live herbaceous moisture content from a seasonal drought index. Int. J. Biometeorol. 2003, 47, 73–79. [Google Scholar] [PubMed]
- Viegas, D.X.; Piñol, J.; Ogaya, R. Estimating live fine fuels moisture content using meteorologically-based indices. Int. J. Wildland Fire 2001, 10, 223–240. [Google Scholar] [CrossRef]
- Xanthopoulos, G.; Wakimoto, R.H. A time to ignition - temperature - moisture relationship for branches of three western conifers. Can. J. For. Res. 1993, 23, 253–258. [Google Scholar] [CrossRef]
- Nelson, R.M. Prediction of diurnal change in 10-h fuel stick moisture content. Can. J. For. Res. 2000, 30, 1071–1087. [Google Scholar] [CrossRef]
- Viney, N.R. A review of fine fuel moisture modelling. Int. J. Wildland Fire 1991, 1, 215–234. [Google Scholar] [CrossRef]
- Bertin, N.; Tchamitchian, M.; Baldet, P.; Devaux, C.; Brunel, B.; Gary, C. Contribution of carbohydrate pools to the variations in leaf mass per area within a tomato plant. New Phytol. 1999, 143, 53–61. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Meinzer, F.C.; Marias, D.E.; Sevanto, S.; Jenkins, M.W.; McDowell, N.G. Linking nonstructural carbohydrate dynamics to gas exchange and leaf hydraulic behavior in pinus edulis and juniperus monosperma. New Phytol. 2015, 206, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Jolly, W.M.; Hadlow, A.M.; Huguet, K. De-coupling seasonal changes in water content and dry matter to predict live conifer foliar moisture content. Int. J. Wildland Fire 2014, 23, 480–489. [Google Scholar] [CrossRef]
- Jolly, W.M.; Hintz, J.; Linn, R.L.; Kropp, R.C.; Conrad, E.T.; Parsons, R.A.; Winterkamp, J. Seasonal variations in red pine (pinus resinosa) and jack pine (pinus banksiana) foliar physio-chemistry and their potential influence on stand-scale wildland fire behavior. For. Ecol. Manag. 2016, 373, 167–178. [Google Scholar] [CrossRef]
- Wever, L.A.; Flanagan, L.B.; Carlson, P.J. Seasonal and interannual variation in evapotranspiration, energy balance and surface conductance in a northern temperate grassland. Agric. For. Meteorol. 2002, 112, 31–49. [Google Scholar] [CrossRef]
- Simms, D.; Law, M. The ignition of wet and dry wood by radiation. Combust. Flame 1967, 11, 377–388. [Google Scholar] [CrossRef]
- Pellizzaro, G.; Cesaraccio, C.; Duce, P.; Ventura, A.; Zara, P. Relationships between seasonal patterns of live fuel moisture and meteorological drought indices for mediterranean shrubland species. Int. J. Wildland Fire 2007, 16, 232–241. [Google Scholar] [CrossRef]
- Barrs, H.D.; Weatherley, P.E. A re-examination of the relative turgidity technique for estimating water deficits in leaves. Aust. J. Biol. Sci. 1962, 15, 413–428. [Google Scholar] [CrossRef]
- Gary, H.L. Seasonal and diurnal changes in moisture contents and water deficits of engelmann spruce needles. Bot. Gaz. 1971, 327–332. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Clausen, J.J. Changes in moisture contents and dry weights of buds and leaves of forest trees. Bot. Gaz. 1965, 126, 20–26. [Google Scholar] [CrossRef]
- Finney, M.A.; Cohen, J.D.; McAllister, S.S.; Jolly, W.M. On the need for a theory of wildland fire spread. Int. J. Wildland Fire 2013, 22, 25–36. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Pickett, B.M.; Isackson, C.; Wunder, R.; Fletcher, T.H.; Butler, B.W.; Weise, D.R. Flame interactions and burning characteristics of two live leaf samples1. Int. J. Wildland Fire 2009, 18, 865–874. [Google Scholar] [CrossRef]
- Schwilk, D.W. Flammability is a niche construction trait: Canopy architecture affects fire intensity. Am. Nat. 2003, 162, 725–733. [Google Scholar]
- Whittaker, R.H. Communities and Ecosystems; Macmillan: New York, NY, USA, 1970. [Google Scholar]
- Cohen, W.B.; Omi, P.N.; Kaufmann, M.R. Heating-related water transport to intact lodgepole pine branches. For. Sci. 1990, 36, 246–254. [Google Scholar]
- Tyree, M.T.; Sperry, J.S. Vulnerability of xylem to cavitation and embolism. Ann. Rev. Plant Biol. 1989, 40, 19–36. [Google Scholar] [CrossRef]
- Kavanaugh, K.L.; Dickinson, M.B.; Bova, A.S. A way forward for fire-caused tree mortality prediction: Modeling a physiological consequence of fire. Fire Ecol. 2010, 6, 80–94. [Google Scholar] [CrossRef]
- Pausas, J.G. Bark thickness and fire regime. Funct. Ecol. 2015, 29, 315–327. [Google Scholar] [CrossRef]
- Vines, R.G. Heat transfer through bark and the resistance of trees to fire. Aust. J. Bot. 1968, 16, 499–514. [Google Scholar] [CrossRef]
- Michaletz, S.T.; Johnson, E.A.; Tyree, M.T. Moving beyond the cambium necrosis hypothesis of post-fire tree mortality: Cavitation and deformation of xylem in forest fires. New Phytol. 2012, 194, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Nature. Available online: https://www.nature.com/subjects/ecophysiology (accessed on 31 January 2018).
- Smith, A.M.; Talhelm, A.F.; Johnson, D.M.; Sparks, A.M.; Kolden, C.A.; Yedinak, K.M.; Apostol, K.G.; Tinkham, W.T.; Abatzoglou, J.T.; Lutz, J.A. Effects of fire radiative energy density dose on pinus contorta and larix occidentalis seedling physiology and mortality. Int. J. Wildland Fire 2017, 26, 82–94. [Google Scholar] [CrossRef]
- Sparks, A.M.; Kolden, C.A.; Smith, A.; Boschetti, L.; Johnson, D.; Cochrane, M.A. Fire intensity impacts on post-fire response of temperature coniferous forest net primary productivity. Biogeosciences 2018. In Press. [Google Scholar]
- West, A.G.; Nel, J.A.; Bond, W.J.; Midgley, J.J. Experimental evidence for heat plume-induced cavitation and xylem deformation as a mechanism of rapid post-fire tree mortality. New Phytol. 2016, 211, 828–838. [Google Scholar] [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jolly, W.M.; Johnson, D.M. Pyro-Ecophysiology: Shifting the Paradigm of Live Wildland Fuel Research. Fire 2018, 1, 8. https://doi.org/10.3390/fire1010008
Jolly WM, Johnson DM. Pyro-Ecophysiology: Shifting the Paradigm of Live Wildland Fuel Research. Fire. 2018; 1(1):8. https://doi.org/10.3390/fire1010008
Chicago/Turabian StyleJolly, W. Matt, and Daniel M. Johnson. 2018. "Pyro-Ecophysiology: Shifting the Paradigm of Live Wildland Fuel Research" Fire 1, no. 1: 8. https://doi.org/10.3390/fire1010008