
Feral Horses and Bison at Theodore Roosevelt National Park (North Dakota, United States) Exhibit Shifts in Behaviors during Drone Flights

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
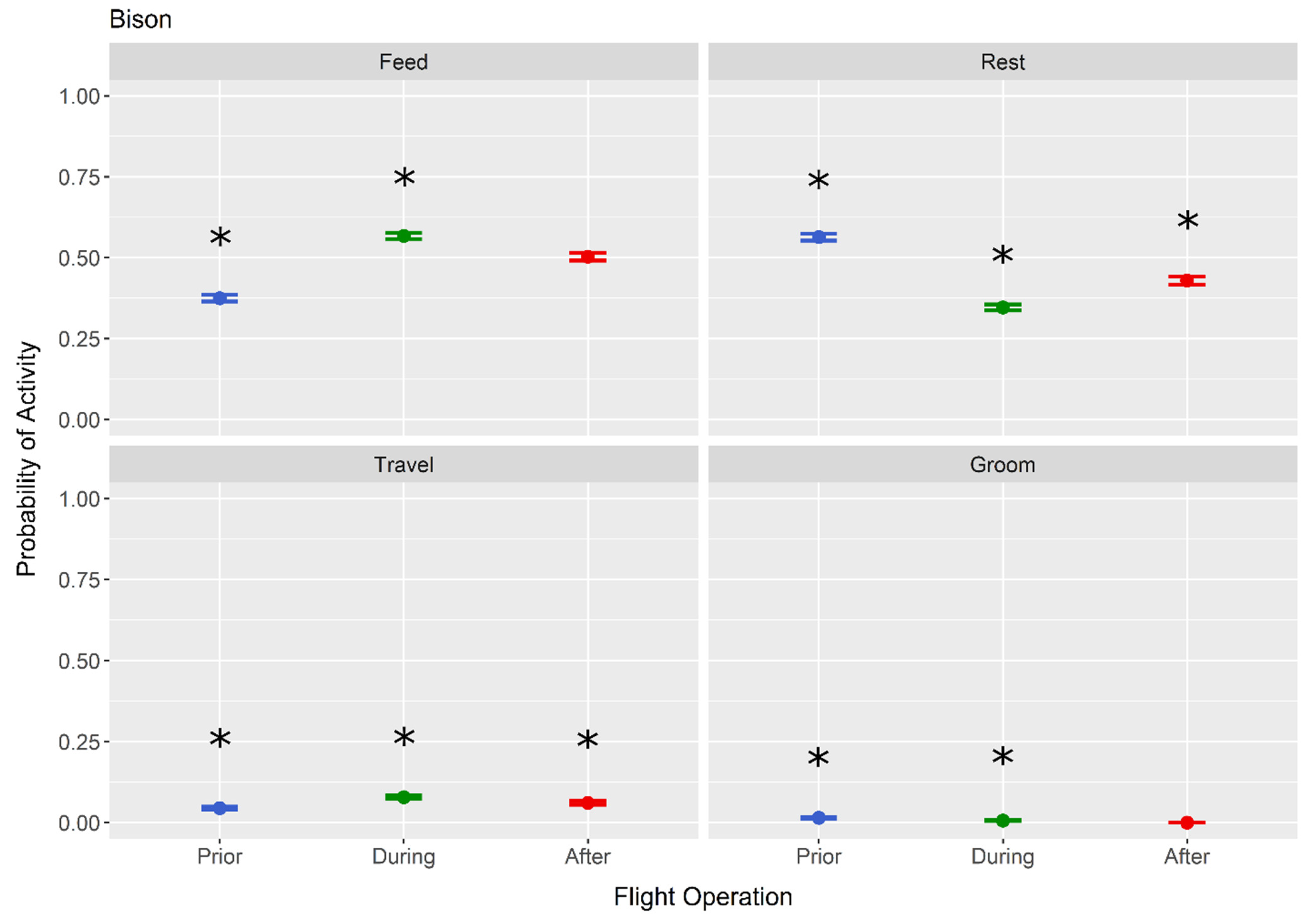
3. Results
4. Discussion
5. Future Work
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Estimate | Standard Error | p-Value | |
|---|---|---|---|
| Feeding | |||
| Pre | −5.46 | 0.41 | <0.001 |
| During | −1.65 | 0.06 | <0.001 |
| Post | −2.00 | 0.09 | <0.001 |
| Resting | |||
| Pre | 1.68 | 0.07 | <0.001 |
| During | −2.03 | 0.07 | <0.001 |
| Post | −0.99 | 0.06 | <0.001 |
| Vigilance | |||
| Pre | −1.82 | 0.07 | <0.001 |
| During | 0.34 | 0.04 | <0.001 |
| Post | −0.11 | 0.06 | 0.065 |
| Traveling | |||
| Pre | −4.85 | 0.30 | <0.001 |
| During | −1.85 | 0.06 | <0.001 |
| Post | −1.83 | 0.08 | <0.001 |
| Grooming | |||
| Pre | −5.17 | 0.35 | <0.001 |
| During | −5.69 | 0.40 | <0.001 |
| Post | −6.25 | 0.70 | <0.001 |
| Behavior | Estimate | Standard Error | p-Value |
|---|---|---|---|
| Feeding | |||
| Pre | −0.51 | 0.05 | <0.001 |
| During | 0.27 | 0.04 | <0.001 |
| Post | 0.01 | 0.05 | 0.786 |
| Resting | |||
| Pre | 0.26 | 0.04 | <0.001 |
| During | −0.63 | 0.04 | <0.001 |
| Post | −0.28 | 0.05 | <0.001 |
| Traveling | |||
| Pre | −3.06 | 0.11 | <0.001 |
| During | −2.46 | 0.07 | <0.001 |
| Post | −2.73 | 0.10 | <0.001 |
| Grooming | |||
| Pre | −4.19 | 0.20 | <0.001 |
| During | −5.06 | 0.22 | <0.001 |
| Post | −19.66 | 458.93 | 0.966 |
References
- Barnas, A.F.; Felege, C.J.; Rockwell, R.F.; Ellis-Felege, S.N. A Pilot (Less) Study on the Use of an Unmanned Aircraft System for Studying Polar Bears (Ursus Maritimus). Polar Biol. 2018, 41, 1055–1062. [Google Scholar] [CrossRef]
- Díaz-Delgado, R.; Mücher, S. Editorial of Special Issue “Drones for Biodiversity Conservation and Ecological Monitoring”. 2019. Available online: https://www.mdpi.com/2504-446X/3/2/47 (accessed on 30 March 2022).
- Linchant, J.; Lisein, J.; Semeki, J.; Lejeune, P.; Vermeulen, C. Are Unmanned Aircraft Systems (UAS s) the Future of Wildlife Monitoring? A Review of Accomplishments and Challenges. Mammal Rev. 2015, 45, 239–252. [Google Scholar] [CrossRef]
- Whitehead, K.; Hugenholtz, C.H.; Myshak, S.; Brown, O.; LeClair, A.; Tamminga, A.; Barchyn, T.E.; Moorman, B.; Eaton, B. Remote Sensing of the Environment with Small Unmanned Aircraft Systems (UASs), Part 2: Scientific and Commercial Applications. J. Unmanned Veh. Syst. 2014, 2, 86–102. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Carignan, V.; Bird, D.M. Measuring Habitat Quality for Least Bitterns in a Created Wetland with Use of a Small Unmanned Aircraft. Wetlands 2014, 34, 527–533. [Google Scholar] [CrossRef]
- Habel, J.C.; Teucher, M.; Ulrich, W.; Bauer, M.; Rödder, D. Drones for Butterfly Conservation: Larval Habitat Assessment with an Unmanned Aerial Vehicle. Landsc. Ecol. 2016, 31, 2385–2395. [Google Scholar] [CrossRef]
- Jiménez López, J.; Mulero-Pázmány, M. Drones for Conservation in Protected Areas: Present and Future. Drones 2019, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Nagy-Reis, M.; Dickie, M.; Calvert, A.M.; Hebblewhite, M.; Hervieux, D.; Seip, D.R.; Gilbert, S.L.; Venter, O.; DeMars, C.; Boutin, S. Habitat Loss Accelerates for the Endangered Woodland Caribou in Western Canada. Conserv. Sci. Pract. 2021, 3, e437. [Google Scholar] [CrossRef]
- Olsoy, P.J.; Shipley, L.A.; Rachlow, J.L.; Forbey, J.S.; Glenn, N.F.; Burgess, M.A.; Thornton, D.H. Unmanned Aerial Systems Measure Structural Habitat Features for Wildlife across Multiple Scales. Methods Ecol. Evol. 2018, 9, 594–604. [Google Scholar] [CrossRef]
- Wich, S.; Dellatore, D.; Houghton, M.; Ardi, R.; Koh, L.P. A Preliminary Assessment of Using Conservation Drones for Sumatran Orang-Utan (Pongo Abelii) Distribution and Density. J. Unmanned Veh. Syst. 2016, 4, 45–52. [Google Scholar] [CrossRef]
- Consi, T.R.; Patzer, J.R.; Moe, B.; Bingham, S.A.; Rockey, K. An Unmanned Aerial Vehicle for Localization of Radio-Tagged Sturgeon: Design and First Test Results. In Proceedings of the OCEANS 2015-MTS/IEEE Washington, Washington, DC, USA, 19–22 October 2015; pp. 1–10. [Google Scholar]
- Hui, N.T.; Lo, E.K.; Moss, J.B.; Gerber, G.P.; Welch, M.E.; Kastner, R.; Schurgers, C. A More Precise Way to Localize Animals Using Drones. J. Field Robot. 2021. Available online: https://kastner.ucsd.edu/wp-content/uploads/2021/04/admin/jfr21-animal_tracking.pdf (accessed on 30 March 2022).
- Saunders, D.; Nguyen, H.; Cowen, S.; Magrath, M.; Marsh, K.; Bell, S.; Bobruk, J. Radio-Tracking Wildlife with Drones: A Viewshed Analysis Quantifying Survey Coverage across Diverse Landscapes. Wildl. Res. 2022. Available online: https://www.researchgate.net/publication/358404185_Radio-tracking_wildlife_with_drones_a_viewshed_analysis_quantifying_survey_coverage_across_diverse_landscapes (accessed on 30 March 2022).
- Bushaw, J.D.; Ringelman, K.M.; Rohwer, F.C. Applications of Unmanned Aerial Vehicles to Survey Mesocarnivores. Drones 2019, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, P.; Steen, K.A.; Jørgensen, R.N.; Karstoft, H. Automated Detection and Recognition of Wildlife Using Thermal Cameras. Sensors 2014, 14, 13778–13793. [Google Scholar] [CrossRef] [PubMed]
- Stander, R.; Walker, D.J.; Rohwer, F.C.; Baydack, R.K. Drone Nest Searching Applications Using a Thermal Camera. Wildl. Soc. Bull. 2021, 45, 371–382. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Stolper, R.; Van Essen, L.D.; Negro, J.J.; Sassen, T. Remotely Piloted Aircraft Systems as a Rhinoceros Anti-Poaching Tool in Africa. PLoS ONE 2014, 9, e83873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivares-Mendez, M.A.; Fu, C.; Ludivig, P.; Bissyandé, T.F.; Kannan, S.; Zurad, M.; Annaiyan, A.; Voos, H.; Campoy, P. Towards an Autonomous Vision-Based Unmanned Aerial System against Wildlife Poachers. Sensors 2015, 15, 31362–31391. [Google Scholar] [CrossRef] [PubMed]
- Chabot, D.; Bird, D.M. Wildlife Research and Management Methods in the 21st Century: Where Do Unmanned Aircraft Fit In? J. Unmanned Veh. Syst. 2015, 3, 137–155. [Google Scholar] [CrossRef] [Green Version]
- Christie, K.S.; Gilbert, S.L.; Brown, C.L.; Hatfield, M.; Hanson, L. Unmanned Aircraft Systems in Wildlife Research: Current and Future Applications of a Transformative Technology. Front. Ecol. Environ. 2016, 14, 241–251. [Google Scholar] [CrossRef]
- Whitehead, K.; Hugenholtz, C.H. Remote Sensing of the Environment with Small Unmanned Aircraft Systems (UASs), Part 1: A Review of Progress and Challenges. J. Unmanned Veh. Syst. 2014, 2, 69–85. [Google Scholar] [CrossRef]
- Barnas, A.; Newman, R.; Felege, C.J.; Corcoran, M.P.; Hervey, S.D.; Stechmann, T.J.; Rockwell, R.F.; Ellis-Felege, S.N. Evaluating Behavioral Responses of Nesting Lesser Snow Geese to Unmanned Aircraft Surveys. Ecol. Evol. 2018, 8, 1328–1338. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Craik, S.R.; Bird, D.M. Population Census of a Large Common Tern Colony with a Small Unmanned Aircraft. PLoS ONE 2015, 10, e0122588. [Google Scholar] [CrossRef] [Green Version]
- Ryckman, M.D.; Kemink, K.; Felege, C.J.; Darby, B.; Vandeberg, G.S.; Ellis-Felege, S.N. Behavioral Responses of Blue-Winged Teal and Northern Shoveler to Unmanned Aerial Vehicle Surveys. PLoS ONE 2022, 17, e0262393. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Jenni-Eiermann, S.; Strebel, N.; Sattler, T.; Negro, J.J.; Tablado, Z. Unmanned Aircraft Systems as a New Source of Disturbance for Wildlife: A Systematic Review. PLoS ONE 2017, 12, e0178448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klaassen, M.; Bauer, S.; Madsen, J.; Tombre, I. Modelling Behavioural and Fitness Consequences of Disturbance for Geese along Their Spring Flyway. J. Appl. Ecol. 2006, 43, 92–100. [Google Scholar] [CrossRef] [Green Version]
- Griffin, L.P.; Brownscombe, J.W.; Gagné, T.O.; Wilson, A.D.; Cooke, S.J.; Danylchuk, A.J. Individual-Level Behavioral Responses of Immature Green Turtles to Snorkeler Disturbance. Oecologia 2017, 183, 909–917. [Google Scholar] [CrossRef] [PubMed]
- McEvoy, J.F.; Hall, G.P.; McDonald, P.G. Evaluation of Unmanned Aerial Vehicle Shape, Flight Path and Camera Type for Waterfowl Surveys: Disturbance Effects and Species Recognition. PeerJ 2016, 4, e1831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vas, E.; Lescroël, A.; Duriez, O.; Boguszewski, G.; Grémillet, D. Approaching Birds with Drones: First Experiments and Ethical Guidelines. Biol. Lett. 2015, 11, 20140754. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, F.; Nielsen, M.L.; Charlton, C.; Bejder, L.; Madsen, P.T. Southern Right Whales Show No Behavioral Response to Low Noise Levels from a Nearby Unmanned Aerial Vehicle. Mar. Mammal Sci. 2020, 36, 953–963. [Google Scholar] [CrossRef]
- Fiori, L.; Martinez, E.; Orams, M.B.; Bollard, B. Using Unmanned Aerial Vehicles (UAVs) to Assess Humpback Whale Behavioral Responses to Swim-with Interactions in Vava’u, Kingdom of Tonga. J. Sustain. Tour. 2020, 28, 1743–1761. [Google Scholar] [CrossRef]
- Landeo-Yauri, S.S.; Castelblanco-Martínez, D.N.; Hénaut, Y.; Arreola, M.R.; Ramos, E.A. Behavioural and Physiological Responses of Captive Antillean Manatees to Small Aerial Drones. Wildl. Res. 2021. Available online: https://www.publish.csiro.au/WR/WR20159 (accessed on 30 March 2022).
- Palomino-González, A.; Kovacs, K.M.; Lydersen, C.; Ims, R.A.; Lowther, A.D. Drones and Marine Mammals in Svalbard, Norway. Mar. Mammal Sci. 2021. Available online: https://www.researchgate.net/publication/350062485_Drones_and_marine_mammals_in_Svalbard_Norway (accessed on 30 March 2022).
- Ramos, E.A.; Maloney, B.; Magnasco, M.O.; Reiss, D. Bottlenose Dolphins and Antillean Manatees Respond to Small Multi-Rotor Unmanned Aerial Systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Smith, C.; Sykora-Bodie, S.; Pack, S.; Bloodworth, B.; Spradlin, T.; LeBoeuf, N. Assessment of Known Impacts of Unmanned Aerial Systems (UAS) on Marine Mammals: Data Gaps and Recommendations for Researchers in the United States. J. Unmanned Veh. Syst. 2016, 4, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Torres, L.G.; Nieukirk, S.L.; Lemos, L.; Chandler, T.E. Drone up! Quantifying Whale Behavior from a New Perspective Improves Observational Capacity. Front. Mar. Sci. 2018, 5, 319. [Google Scholar] [CrossRef] [Green Version]
- Bennitt, E.; Bartlam-Brooks, H.L.; Hubel, T.Y.; Wilson, A.M. Terrestrial Mammalian Wildlife Responses to Unmanned Aerial Systems Approaches. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ditmer, M.A.; Vincent, J.B.; Werden, L.K.; Tanner, J.C.; Laske, T.G.; Iaizzo, P.A.; Garshelis, D.L.; Fieberg, J.R. Bears Show a Physiological but Limited Behavioral Response to Unmanned Aerial Vehicles. Curr. Biol. 2015, 25, 2278–2283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunton, E.; Bolin, J.; Leon, J.; Burnett, S. Fright or Flight? Behavioural Responses of Kangaroos to Drone-Based Monitoring. Drones 2019, 3, 41. [Google Scholar] [CrossRef] [Green Version]
- Lu, V.; Xu, F.; Turghan, M.A. Przewalski’s Horses (Equus Ferus Przewalskii) Responses to Unmanned Aerial Vehicles Flights under Semireserve Conditions: Conservation Implication. Int. J. Zool. 2021, 2021, e6687505. [Google Scholar] [CrossRef]
- Schroeder, N.M.; Panebianco, A. Sociability Strongly Affects the Behavioural Responses of Wild Guanacos to Drones. Sci. Rep. 2021, 11, 20901. [Google Scholar] [CrossRef] [PubMed]
- National Park Service Foundation Document. Theodore Roosevelt National Park. Department of Interior, 2014; 64p. Available online: https://www.nps.gov/thro/learn/management/upload/Theodore-Roosevelt-National-Park-Foundation-Document-2014.pdf (accessed on 30 March 2022).
- Baker, D.L.; Powers, J.G.; Ransom, J.I.; McCann, B.E.; Oehler, M.W.; Bruemmer, J.E.; Galloway, N.L.; Eckery, D.C.; Nett, T.M. Reimmunization Increases Contraceptive Effectiveness of Gonadotropin-Releasing Hormone Vaccine (GonaCon-Equine) in Free-Ranging Horses (Equus Caballus): Limitations and Side Effects. PLoS ONE 2018, 13, e0201570. [Google Scholar] [CrossRef] [PubMed]
- Ransom, J.I.; Powers, J.G.; Garbe, H.M.; Oehler, M.W.; Nett, T.M.; Baker, D.L. Behavior of Feral Horses in Response to Culling and GnRH Immunocontraception. Appl. Anim. Behav. Sci. 2014, 157, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Hansen, K.; Mosley, J. Effects of Roundups on Behavior and Reproduction of Feral Horses. J. Range Manag. 2000, 53, 479–482. [Google Scholar] [CrossRef]
- Shult, M.J. American Bison Behavior Patterns at Wind Cave National Park; Iowa State University, 1972. Available online: https://core.ac.uk/download/pdf/38916419.pdf (accessed on 30 March 2022).
- Saitoh, T.; Kobayashi, M. Appropriate Drone Flight Altitude for Horse Behavioral Observation. Drones 2021, 5, 71. [Google Scholar] [CrossRef]
- McDonnell, S.; Torcivia, C. Preliminary Proof of the Concept of Wild (Feral) Horses Following Light Aircraft into a Trap. Animals 2020, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Linklater, W.L.; Cameron, E.Z. Escape Behaviour of Feral Horses during a Helicopter Count. Wildl. Res. 2002, 29, 221–224. [Google Scholar] [CrossRef]
- Fortin, D.; Andruskiw, M. Behavioral Response of Free-Ranging Bison to Human Disturbance. Wildl. Soc. Bull. 2003, 31, 804–813. [Google Scholar]
- Borkowski, J.J.; White, P.J.; Garrott, R.A.; Davis, T.; Hardy, A.R.; Reinhart, D.J. Behavioral Responses of Bison and Elk in Yellowstone to Snowmobiles and Snow Coaches. Ecol. Appl. 2006, 16, 1911–1925. [Google Scholar] [CrossRef]
- Hardy, A.R. Bison and Elk Responses to Winter Recreation in Yellowstone National Park. Master’s Thesis, Montana State University-Bozeman, College of Letters & Science, 2001. Available online: https://www.snowmobileinfo.org/snowmobile-access-docs/Bison-elk-Responses-to-Winter-Recreation-in-Yellowstone_2001.pdf (accessed on 30 March 2022).
- Anderson, B.A. A Literature Review of the Effects of Helicopter Disturbance and Noise on Selected Wildlife Species; ABR, Inc.—Environmental Research & Services: Anchorage, Alaska, 2007; p. 230. [Google Scholar]
- McFarland, D.J. On the Causal and Functional Significance of Displacement Activities. Z. Für Tierpsychol. 1966, 23, 217–235. [Google Scholar] [CrossRef]
- Tinbergen, N. “Derived” Activities; Their Causation, Biological Significance, Origin, and Emancipation during Evolution. Q. Rev. Biol. 1952, 27, 1–32. [Google Scholar] [CrossRef]
- MacArthur, R.A.; Geist, V.; Johnston, R.H. Cardiac and Behavioral Responses of Mountain Sheep to Human Disturbance. J. Wildl. Manag. 1982, 46, 351–358. [Google Scholar] [CrossRef]
- Ellis-Felege, S.N.; Stechmann, T.J.; Hervey, S.D.; Felege, C.J.; Rockwell, R.F.; Barnas, A.F. Nesting Common Eiders (Somateria Mollissima) Show Little Behavioral Response to Fixed-Wing Drone Surveys. J. Unmanned Veh. Syst. 2021. Available online: https://cdnsciencepub.com/doi/full/10.1139/juvs-2021-0012 (accessed on 30 March 2022).
- Hodgson, J.C.; Koh, L.P. Best Practice for Minimising Unmanned Aerial Vehicle Disturbance to Wildlife in Biological Field Research. Curr. Biol. 2016, 26, R404–R405. [Google Scholar] [CrossRef] [Green Version]


| Behavior | Description |
|---|---|
| Horse | |
| Feeding | Active foraging with minimum travel |
| Resting | Head hanging, not attentive standing, absence of other behaviors |
| Vigilance | Head above, wither shoulders, and ears forward |
| Traveling | Walking and incidental grazing |
| Grooming | Licking and scratching |
| Bison | |
| Feeding | Active foraging |
| Resting | Standing or laying, head slung |
| Traveling | Active moving without foraging, walking, or running |
| Grooming | Rubbing, scratching with hoof |
| Wallowing | Rolling, dusting, grooming unless urinating and rolling in it |
| Agonistic | Pawing, wallowing with urination, rolling. Usually these behaviors occur in sequence in the presence of two or more males and are more common at the beginning of the breeding season |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenzi, J.; Felege, C.J.; Newman, R.; McCann, B.; Ellis-Felege, S.N. Feral Horses and Bison at Theodore Roosevelt National Park (North Dakota, United States) Exhibit Shifts in Behaviors during Drone Flights. Drones 2022, 6, 136. https://doi.org/10.3390/drones6060136
Lenzi J, Felege CJ, Newman R, McCann B, Ellis-Felege SN. Feral Horses and Bison at Theodore Roosevelt National Park (North Dakota, United States) Exhibit Shifts in Behaviors during Drone Flights. Drones. 2022; 6(6):136. https://doi.org/10.3390/drones6060136
Chicago/Turabian StyleLenzi, Javier, Christopher J. Felege, Robert Newman, Blake McCann, and Susan N. Ellis-Felege. 2022. "Feral Horses and Bison at Theodore Roosevelt National Park (North Dakota, United States) Exhibit Shifts in Behaviors during Drone Flights" Drones 6, no. 6: 136. https://doi.org/10.3390/drones6060136