Invasive Rainbow Trout (Oncorhynchus mykiss) Are Not Affected by Different Land Uses in a Multi-Use, Mediterranean Climate Landscape


1
Centro Interdisciplinario de Cambio Global, Pontificia Universidad Católica de Chile, Santiago 7820436, Chile
2
Departamento de Ecosistemas y Medio Ambiente, Pontificia Universidad Católica de Chile, Santiago 7820436, Chile
3
School of Geography, University of Lincoln, Lincoln, LN6 7TS, UK
*
Author to whom correspondence should be addressed.
Fishes 2018, 3(4), 37; https://doi.org/10.3390/fishes3040037
Submission received: 1 August 2018
/
Revised: 20 September 2018
/
Accepted: 21 September 2018
/
Published: 26 September 2018
Abstract
:Land use carries implications for freshwater fish conservation. Plantation forestry practices have been shown to have negative impacts on resident fish fauna, but little work has been conducted to assess these impacts on invasive vs. native fish populations. Ten headwater catchments in the Mediterranean climate zone of Chile were used to assess the impacts of land use (pine plantations vs. native forests) on fish condition (length-weight relationship) and abundance (catch-per-unit-effort (CPUE)) of the invasive trout Oncorhynchus mykiss and the threatened native catfish Nematogenys inermis. Negative impacts on trout condition were associated with a lack of canopy cover and river topology. The presence of N. inermis was associated with catchment factors less favourable to trout. Current environmental regulations and forestry management practices do not appear to create negative pressures on invasive trout from land use practices, despite expectations from the literature. Assessing how land use management regulations impact invasive and native fishes should be a part of species conservation and territorial planning.
1. Introduction
Humans impact landscapes and waterscapes in many different ways, including land cover composition, water quality degradation and water quantity diminution. In addition to impacts from landscape-scale changes, the introduction of exotic species adds additional pressures to changing local ecological communities, sometimes becoming invasive. Although altered ecosystems can function as a control for invasive species, very few studies have focused on this possibility. One reason for this lack of attention could be due to self-selection among ecologists to study natural ecological conditions over altered and invaded ones [1]. However, with our understandings of the current and future anthropogenic and climate change effects on river habitat [2,3,4], there is a need to study the management of altered and invaded spaces within a greater ecological conservation context. What is more, in data-deficient areas that are experiencing changes in land use and water use on top of the impacts of invasive species, the tacit paradigm of natural area conservation can actually hide the chances of using altered landscapes and riverscapes for conserving threatened and endangered freshwater species.
Throughout the Southern Hemisphere, trout species were introduced for sport fishing purposes from the end of the 19th Century and through to the 20th Century [5] and have become invasive in many countries, including Argentina, Australia, South Africa, India and Vietnam. In Chile, trout were introduced for sport fishing in 1890 [6], and presently, brown trout (Salmo trutta) or rainbow trout (Oncorhynchus mykiss) can be found in the mountainous reaches of all of Chile’s perennially flowing rivers [7], with trout displaying a seasonal range expansion and contraction that follows water temperature in Mediterranean climate river systems [8]. Since their introduction to Chile, trout have successfully outcompeted native fishes through habitat exclusion [9,10] and predation [11], decimating a freshwater fish fauna characterized by low diversity (46 species across the entire country), high endemism [7] and high levels of endangerment (all but three species listed as vulnerable or endangered) [12], with several species being the sole existing member of an entire family, such as Nematogenys inermis, the Chilean mountain catfish.
For ecologists and environmental managers in areas with trout invasions, conservation requires the determination of: (i) how to improve conditions for native species relative to that of trout, preferably with little additional cost; and (ii) how to manage trout, which are the primary sport fishing species. This challenge requires reassessing some conservation assumptions, based on local ecological contexts, especially about the responses to human activities, both those conducted with and without biological conservation in mind. It is in this context of reassessing conservation assumptions that we examine the effect of existing human landscape change on native fishes vs. trout. A net benefit for native species could mean an unexpected conservation pathway that utilizes the human landscape (often considered detrimental to native species) to provide comparative benefits to native species.
There are already examples of using altered ecosystems for conserving native ecologies [13]. In the aquatic context, the Great Lakes of North America have seen extensive land use change and exotic species introductions [14], including sea lamprey (Petromyzon marinus), a migratory species that parasitizes several lake and river fishes [15]. Here, there is a relative benefit to upstream ecosystems of using pre-existing dams near river mouths to block sea lamprey migration and avoid parasitism while removing upstream dams or providing fish passage to expand lamprey-protected habitat [16,17].
Just as there are examples of unexpected benefits from human alterations, there are examples of conservation interventions that have been designed for one ecological context causing unintended, negative consequences in other contexts. Therefore, the conservation utility of different human interventions changes based on ecological context, and the same conservation measures developed to protect trout in their native habitats [18,19,20] have helped spread invasive trout in Chile. Understanding the lessons learned about the causes for trout endangerment in their native habitats could be useful for finding a negative control on invasive trout in Chile, while offering comparative benefits for native species.
1.1. Negative Impacts of Forestry on Trout
Differences in land use across the landscape can alter fish community structure and dynamics by altering the local hydrology, water chemistry or sediment regimes [21,22,23,24]. Large-scale impacts from human land uses (such as agriculture or urbanization) can change these parameters beyond optimal or even tolerable levels for fish [25], especially highly specialized species like trout [26,27]. Plantation forestry in particular has shown negative impacts on brown trout [28], bull trout [29], lake trout [30] and rainbow trout [31], among others, showing the potential for forestry to act as a negative control for trout, as observed tangentially in plantations of exotic pine in southern Chile [32]. What is less known is the comparative impact of plantation forestry on native fishes.
Riparian buffers help ameliorate the impacts of otherwise significant land use effects [33]. In forestry plantations, wide riparian buffers help moderate water temperatures [34], improve water quality [35,36,37] and diminish impacts from siltation [38,39]. A well-maintained riparian buffer provides several benefits for trout in Mediterranean climate headwaters, where a lack of canopy cover could raise summer temperatures well above 18 °C, with potentially high sediment fluxes [40]. Trout preferentially seek out low-temperature streams with low levels of siltation [8], excluding native species. In contrast, the far more generalist native Chilean species, adapted to high fluctuations in water temperature and sediment transport characterizing river systems in Mediterranean climates [7,9], currently occupy areas that are less suitable for trout [41]. Since native fishes of the Mediterranean climate of Chile have a competitive advantage over trout in warmer rivers, this implies that existing landscape alterations that raise water temperatures and/or increase siltation could provide a comparative advantage to native fishes by providing less suitable trout habitat.
1.2. Measures of Trout Quality
The lack of fish monitoring in Chile means that estimates of fish abundance and physical condition are limited, even in conservation or planning contexts [42], necessitating the use of rapid assessment techniques to examine relative population sizes and structures. Since headwater systems throughout Central Chile are dominated by exotic trout [7] and trout have a significant ecological impact when present, rapid assessment tools can help determine the likely condition of a resident trout population. The condition indices of Fulton’s k are useful in estimating relative population sizes of invasive trout in Chilean oligotrophic lakes [43]. Although trout populations are regulated by density-dependant thinning and mortality [44], different streams express different factors that likely impact trout survival and growth (e.g., water temperature, river depth, substrate). This array of external controls can lead to significant differences in the ability of resident fish to gain weight, with populations from different catchments displaying a distinct condition. Thus, density-dependant population control is influenced by environmental factors, which limit trout condition as they approach environmental limits. Fish condition thus provides a means of assessing the negative impacts of land use and of population densities.
1.3. Central Chilean Headwaters
Mountain headwater systems of Chile’s Mediterranean zone have relatively steep slopes and flow over cobble and gravel beds, with some catchments containing small cascades. The water source for these streams is primarily from snow and glacier melt in the Andes and from rain along the coastal range [45]. Although air temperatures can rise above 20 °C, shading is often complete [40], helping buffer summer water temperatures [46]. Most of these headwater systems contain no notable sources of water pollution, and macroinvertebrate species that indicate good water quality (e.g., Ephemeroptera, Plecoptera and Trichoptera) are abundant. Given these factors, it is unsurprising to find trout throughout these headwaters. The objective of this paper is to assess differences in fish condition and abundance of native versus non-native fish species among ten headwater systems in Central Chile between catchments dominated by pine plantations and native forests. If pine plantations do exert a negative impact on trout, we expect to see a lower physical condition in trout populations in those streams.
2. Results
After sampling ten sites in mountain headwater streams in Central Chile, one site (C2-F), which was ecologically disconnected from the rest of the watershed due to a small waterfall downstream of the sampling site, contained no fish. Across all sites, only three species were found; O. mykiss were found in nine sites (N = 342), S. trutta in six sites (N = 72) and native mountain catfish, N. inermis, in two sites (N = 20, Table 1). Due to the low numbers of S. trutta across sites, possibly due to competitive exclusion by O. mykiss [47] and their lack of overlap with N. inermis, they were excluded from future assessments. The sampled sites were characterized by cool, relatively narrow streams with rock and gravel bottoms and heavy canopy cover provided by relatively wide riparian vegetative buffers, except for study site A1-F, which had cold water, but was a relatively wide, boulder-dominated stream with no canopy cover.
2.1. Oncorhynchus mykiss
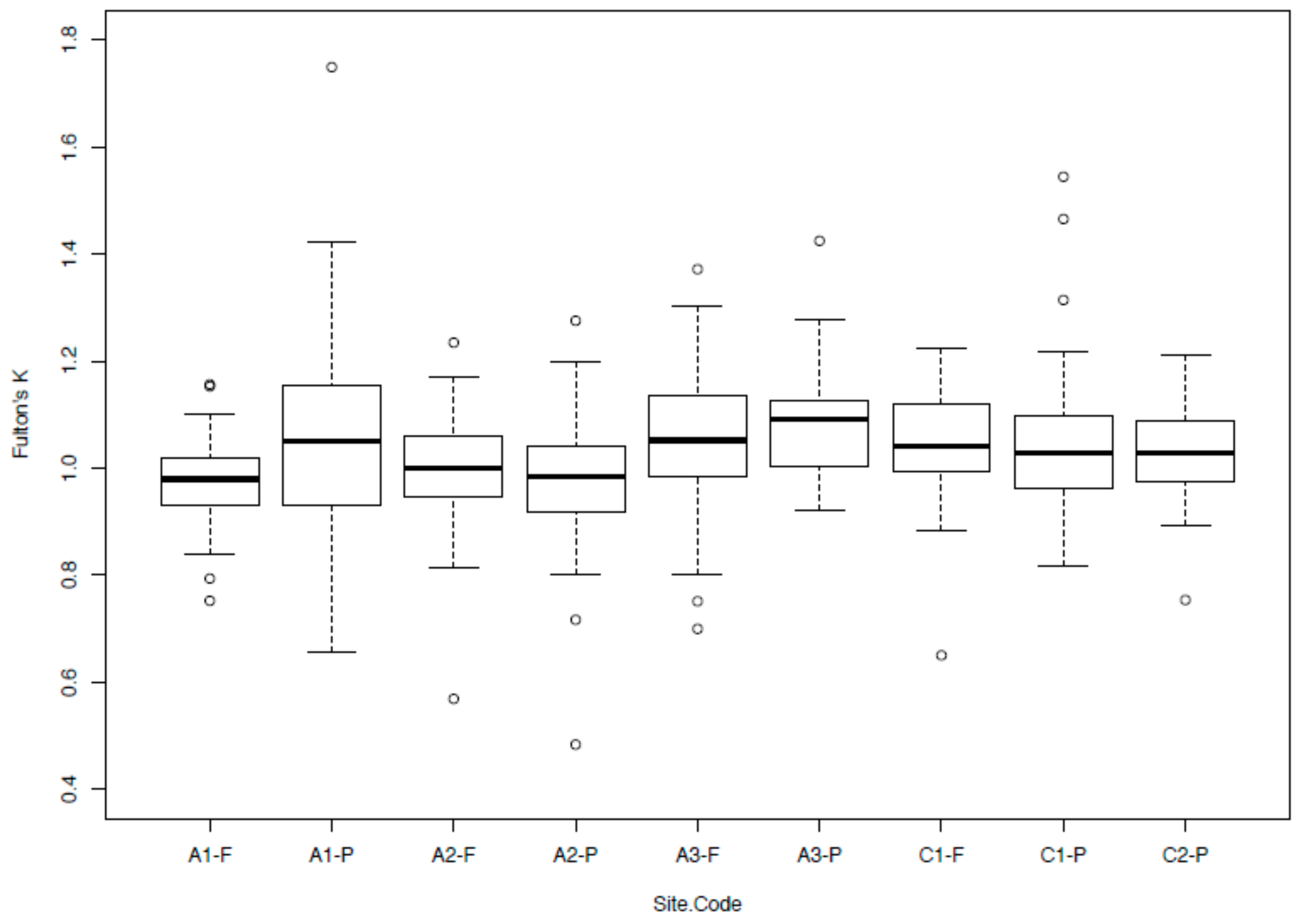
Non-young-of-year O. mykiss were found in nine of the ten sampling sites and ranged in size from 68 mm to 310 mm in total length (mean: 136 ± 1 mm) and 5–318 g in weight (mean: 32.2 ± 0.3 g). Estimated abundances of O. mykiss ranged from 13.6 ± 1.5–204.2 ± 9.6 fish/500 m2 (mean: 68.9 ± 4.4), but all populations exhibited characteristic linear relationships of log-transformed total length-to-weight fish condition measurements (R2 > 0.90) that were all highly significant (p < 0.0001, Table 1). The mean Fulton’s k between sites ranged from 0.97 (A2-P) to 1.08 (A1-P). Permutational multivariate analysis of variance (PERMANOVA) results indicate that fish condition differed significantly among catchments (F = 12.22, p < 0.0001, Figure 1), with A1-F showing a significantly lower condition than half of the other sites (A3-F, A3-P, C1-F and C1-P); A2-P had a significantly lower condition than three sites (A3-F, A3-P and C1-P); and A3-P showed a significantly higher condition than three sites (A1-F, A2-F and A2-P). The permutational analysis of covariance (pANCOVA) results indicate that coastal O. mykiss had a higher condition than Andean communities (p = 0.0306). Pine plantation O. mykiss were not significantly different from those found in native forests (p = 0.5942).
2.2. Nematogenys inermis
The two populations of N. inermis were encountered in pine plantation catchments and ranged in size from 109–290 mm (mean: 178 ± 5 mm) and 7–252 g (mean: 66.6 ± 1.8 g). One was located on the Coastal Range (C1-P, 21.8 ± 1.9 fish/500 m2), and the other was in the Andes (A1-P, 12.6 ± 0.5 fish/500 m2). Due to the low number of individuals at A1-P, cross-site assessment was not possible. Combining the two populations produced a statistically-significant linear relationship (Table 1).
2.3. Body Condition Index
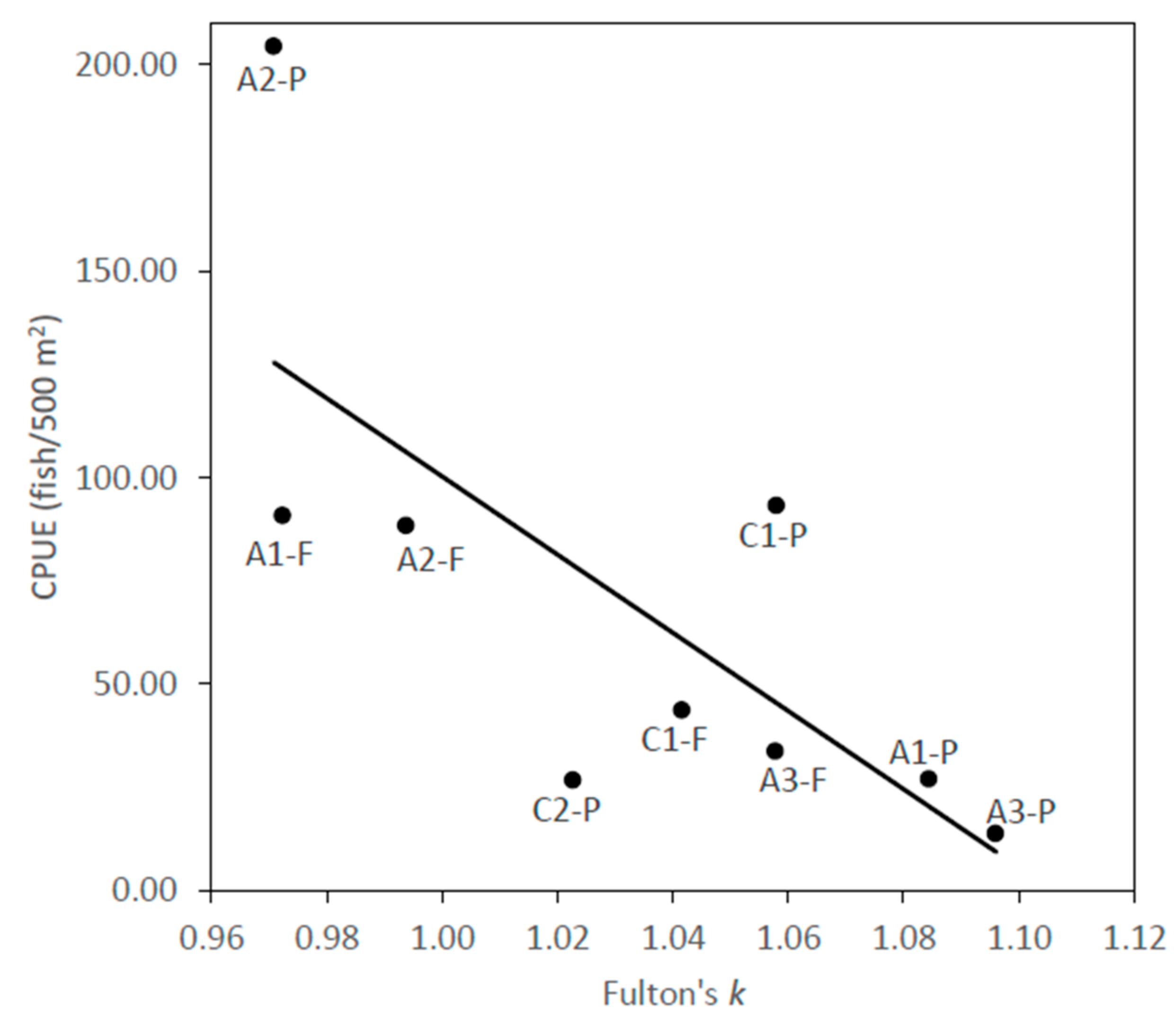
There were no statistically-significant differences in the relationship between condition (k) and O. mykiss catch per unit effort (CPUE) between pine plantation and native forest sites (p = 0.5942), so all sites were combined, which produced a statistically-significant relationship (F = 7.61, p = 0.0281, R2 = 0.521, Figure 2). The significant reduction of overall CPUE with increasing general condition of O. mykiss indicates the extent to which the physical contexts found across the sites were optimal for O. mykiss, which dominated the catch totals across all sites.
3. Discussion
The mountain headwater streams of Chile’s Mediterranean climate rivers feature many physical characteristics that are preferred by trout (i.e., cool water temperatures, shade to provide temperature stability, relatively high dissolved oxygen levels and good water quality). The presence of O. mykiss at all sites with fish is yet more confirmation of their widespread presence in Central Chilean mountain headwaters. Examining the environmental factors found at the pine plantation sites with resident N. inermis, a threatened endemic species, allows for assessing whether its presence is associated with particular environmental factors and differences in the fish condition of co-resident O. mykiss. Finally, the application of the rapid assessment methodology of Arismendi et al. [43] can help assess the impacts of trout numbers on their relative condition.
3.1. Oncorhynchus mykiss
Although A1-F was the largest site, this community had a significantly lower condition than several other sites. This likely was caused by the lack of any canopy, leading to very low allochthonous carbon inputs. The main channel of A1-F is composed of large boulders and cobble, in a typical step-pool morphology [48], and although this may provide a lot of potential habitat, the relatively low productivity—and associated food-web dynamics—would mean that resident trout would have far less food available. The same effect on lowered condition is seen in A2-P, which had no canopy cover just upstream of the sampling site and very little allochthonous carbon in the channel.
O. mykiss populations from Coastal Range headwaters had a higher condition than those from the Andes. One reason for this difference can be explained by topology, with the catchments of the Coastal Range being located closer to larger river confluences (Figure 3), which allows these populations quicker access to larger catchments [49]. However, there were no significant differences in either slope or intercept when considering pine plantation and native forest streams. It is possible that topology has a strong levelling effect on the fish conditions between the pine plantations versus native forests in the Coastal Range; more assessments in either the Andes or Coastal ranges can help disentangle these effects.
3.2. Nematogenys inermis
The threatened N. inermis were caught alongside O. mykiss, indicating that the habitat requirements of N. inermis overlap with those of O. mykiss. Averaged across both sites, the abundance of O. mykiss (59.9 fish/500 m2) was much higher than N. inermis (17.2 fish/500 m2), indicating that N. inermis might be suffering some effect of competitive exclusion from O. mykiss [10].
In examining which physical features of the site were significantly different in C1-P and A1-P from the rest of the sites [50], likely factors are low slopes, long or deep pools and very low amounts of wood. The presence of N. inermis in these sites is supported by some of the scant literature about the species that describes the optimal habitat for this species. Juvenile N. inermis (60–180 mm) prefer deeper areas (0.4–0.7 m) with much vegetation, river velocities between 0.10 and 0.71 m/s and water temperatures between 13 °C and 33 °C. Adults (>190 mm) prefer burrows in the stream bank with temperatures ranging from 10–28 °C [51]. The sampled catfish were all found in physical conditions associated with their age-class, as described in the literature. However, the water temperatures of these sampling sites were low (Table 2), buffered by near-complete canopy cover, and falling at the low end of the range for both O. mykiss and N. inermis.
3.3. Body Condition Index
The statistically-significant relationship between O. mykiss condition and total CPUE (fish/500 m2) indicates that increasing numbers of trout are negatively correlated with fish condition (Figure 2), regardless of forest type, indicating that the populations of rainbow trout in these headwaters are self-regulating [44] and that physical contexts—such as land use—do not appear to exert a negative influence. This corresponds with our findings of a lack of difference between the physical condition of populations in pine plantations versus native forests.
This rapid assessment tool, first proposed for trout invasions of oligotrophic lakes [43], functions with the same tendency in these headwater streams, although with far less predictive power (R2 = 0.5117 vs. 0.95). The continued utility of this metric in a different habitat and climatic setting indicates that it could be used in rapid assessments of invasive trout in other ecosystems. However, the lower strength of the relationship in these headwater systems indicates that the heterogeneity of these sites is higher than what is found in southern Chilean oligotrophic lakes. It is also possible that the sampled individuals are using headwater systems as thermal refugia during the summer months and spend the remaining months in still more diverse conditions. Further assessment of the relationship between habitat factors and body condition in other Mediterranean climate headwaters can help elucidate how abiotic and biotic controls affect invasive trout populations.
3.4. Implications for Invasive Species and Conservation Management
One surprising finding from this study is that pine plantations do not have any significant negative effect on trout condition, despite the various findings in the literature suggesting otherwise [28,29,30,31,32]. This is likely due to the large, intact vegetated margins that were found across nearly all sites, which diminished the effects of thermal gain [34,35,36,37,38,39]. O. mykiss fish condition appears to be influenced by various environmental factors at different scales, with overall basin topology (Andean catchments are associated with lower condition) creating larger scale patterns of fish condition.
The lack of any negative effect of pine plantations on trout condition indicates that the habitat factors found in these streams are as sufficient for resident trout as those found in native forest streams. This means that trout can occupy and displace native fishes [9,10] in potentially all headwater systems throughout the Mediterranean zone, with shaded headwaters acting as seasonal thermal refugia [8], as well as providing exclusive areas for egg-laying and juvenile growth.
However, pine plantation sites were also the only sites with the native catfish, N. inermis, indicating that such human-modified landscapes can provide habitat for native fishes. Due to the lack of data on N. inermis, it is unknown how sampled fish condition compares with other populations. Still, N. inermis were found in sites matching their habitat requirements [51] within a human-altered landscape, indicating that any assumptions about this endemic species preferring natural habitats do not hold. Furthermore, it is possible that landscapes can be managed to optimize environmental characteristics for the preferences of native species (e.g., warmer waters, slower velocities). This means that areas not traditionally considered to be part of conservation efforts—such as pine plantations—can be used to serve a conservation purpose.
River management strategies to exclude trout require a variety of mechanisms. One potential mechanism is evaluating ways to utilize existing human-altered landscapes to create areas less suitable for invasive trout and more suitable for threatened and endangered native species. To address trout habitat management, one could artificially modify the channel by adding in-channel wood [52] to diminish local slopes, deepen channels and diminish water velocity. Another mechanism would be to diminish the canopy cover from riparian vegetation. In this study, the sites with trout having significantly lower Fulton’s k were both in sites with complete or a partial lack of canopy cover, which diminished the allochthonous carbon inputs, leading to lower numbers of prey species. Removal of canopy cover would also permit radiative heating [46], possibly increasing water temperatures to be higher than what trout can tolerate [20,26,27], but be well within the preferences for local native fishes [51]. Other mechanisms could capitalize on the thermal range limits of trout to upland streams [8] in this region to construct barriers to fish passage at or near the summer range limit for trout. These barriers would segregate native species from invasive trout, which could diminish the predatory pressure and competitive resource use that trout impose on native fishes [53], while also serving to provide water for the pine plantations. Assessments of any land-management mechanism would need to be conducted to monitor the effects of such alterations, since the alteration of one factor can have cascading ecosystem effects [54,55].
Beyond land management approaches, the application of fishing management could be applied, but such an approach would be limited to Southern Chile. Although trout can be found in all Andean basins across the length of the country, their presence is limited to increasing elevations as one moves equatorward (i.e., north). This means that access to trout streams by the public becomes ever more difficult as one moves north from Southern Chile, that trout streams become more relegated to mountain areas and that the number and quality of roads accessing them diminish. Even in the study area, trout fishing is not commonplace, since trout populations are seasonally restricted to mountain streams and road access is limited. In Southern Chile, where trout have little or no seasonal restriction to lowland systems and where trout fishing is far more commonplace, fishing management could be pursued, but it will require far more institutional-governance capacity in freshwater ecosystems than currently exists in the country [42].
The current situation of trout invasion speaks to the important role that land management, environmental regulation [40] and forestry management can play in unintentionally fostering the widespread creation and maintenance of trout habitat in headwater streams. Given that trout abundance is the only apparent control on invasive trout in these headwaters and that ecological controls from native species are next to nil [8,11,41], native species conservation measures should assess the ways to diminish trout habitat availability while maintaining—or expanding—habitat availability for native species. The hope generated from the literature of finding a negative control for trout in the form of plantation forestry was not realized in Central Chile, but this only raises questions for how to combine forestry management in new ways that could be beneficial to the particular ecological contexts required by these native species. The strong invasive impacts of trout in Southern Hemisphere Mediterranean climate rivers means that future assessments should examine the relative impacts of land management on native vs. exotic species in order to align land conservation goals with river conservation.
4. Materials and Methods
4.1. Study Area
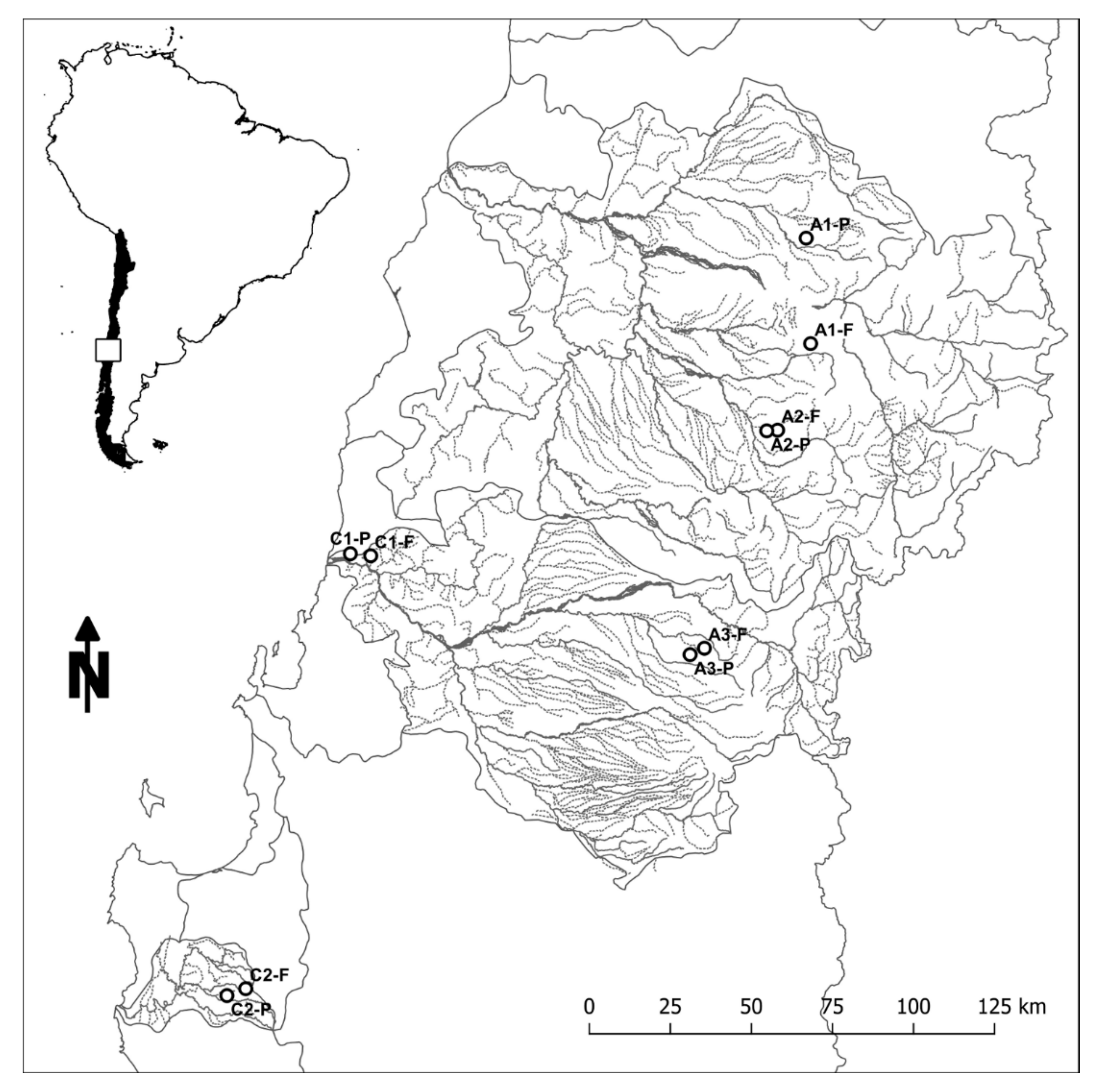
Seven Andean and four Coastal Range headwater catchments in Chile’s Mediterranean zone were selected for this study (Figure 3). Ten of the eleven study sites were selected to represent catchments paired along similar characteristics of slope, aspect, soils, drainage area, catchment orientation, climate, hydrologic regime and proximity (Table 2), allowing us to investigate differences caused by distinct land uses (i.e., native forest vs. pine plantations). For more information about the stream selection process and the physical characteristics of each site, see Mao et al. [50].
4.2. Field Sampling
Fish sampling was conducted between mid-December 2014 and mid-February 2015 (i.e., Southern Hemisphere summer). The length of electrofishing sampling sites ranged from 62–160 m, with the sampling length being at least 10-times the channel width and including at least two pool-riffle sequences.
Fish communities were sampled using upstream, double-pass electrofishing (HT-2000 Battery Backpack Electrofisher, Halltech Aquatic Research, Inc., Guelph, ON, Canada) with blocking nets. All fish were identified to the species level, measured for total length (mm) and weighed (g) before being returned to the river. Environmental variation was quantified using catchment variables derived in QGIS [56]. Local topography and channel-form were measured at the site, as were discharge, conductivity and water temperature. Abundances were standardised to unit-effort measurements (individuals-per-500 m2) Young-of-year were excluded from the assessment. Fish sampling and assessment protocols were approved by the Chilean Fisheries and Aquaculture Subsecretariat (SUBPESCA), Authorization Number 14.875.
4.3. Comparative Fish Condition Assessment
We followed the methods outlined by Lamothe et al. [57] to compare the differences in condition between sites and differences associated with large-scale factors. Briefly, we calculated Fultion’s k using the equation k = (W/TL3) × 100, where W is fish weight (g) and TL is fish length (mm). Using the adonis function in the vegan package, we conducted a PERMANOVA with 9999 permutations to assess differences in Fulton’s fish condition between sites. Then, we used pair-wise t-tests to conduct post-hoc assessments of between-site differences. Separate assessments were conducted for each species. In order to assess the impact of large-scale factors on the slope and y-intercept in length-weight linear-relationships, we log-transformed both length and weight and then conducted pANCOVAs using the aovp function in the lmPerm package with 9999 permutations that grouped the trout according to dominant land use (native forest vs. pine plantation) and topology (Andean catchment vs. Coastal catchment).
To generate the trout body condition index to examine the strength of self-regulation of trout populations, body condition (k) for O. mykiss was determined using the equation k = (W/TL3) × 100. The statistical significance and predictive strength were then compared against CPUE [43]. Catch per unit effort (CPUE) (n/500 m2) was calculated by using the estimated abundances for each sampling site [58] and then standardizing them to a constant area, based on field surveys [50]. All statistical assessments were conducted in R [59].
5. Conclusions
This study was based on the idea of examining whether pine plantations—often regarded as an impediment to conservation—could play a conservation role in Chile by having a negative impact on invasive trout, building on prior research. It focused on headwater streams of the Central Chilean uplands, where plantation forestry is widely practiced, trout are abundant and scientific information is scarce, to see whether existing land use patterns created detectable changes in the physical condition of resident fishes. This research interrogates little-questioned conservation norms of whether highly altered landscapes (pine plantations) can be conservative for fish. Trout abundance was found to be the only dominant mechanism in affecting population health (measured as body condition), indicating that current landscape management does not negatively impact trout, which is surprising, given prior research. We hypothesize that allowing greater solar gain to mid-latitude mountain streams can have a negative impact on resident trout, which can help open up stream habitat for native species. Such research can be pursued in other Southern Hemisphere Mediterranean systems with trout-invaded headwaters, such as those found in Chile, South Africa and Australia.
Author Contributions
Conceptualization, S.L. Funding acquisition, L.M. Investigation, F.U. Methodology, S.L. Project administration, L.M. Writing, original draft, S.L. Writing, review and editing, F.U. and L.M.
Funding
This research was funded by the Chilean National Science and Technology Commission (CONICYT), Grant Number USA2012-0011.
Acknowledgments
We thank Joaquin Lobato, Ernesto Cobo, Diego Silva and Francisco Novoa for helping collect the fish and habitat data. We also thank Victoria Madrid for helping assess catchment-based variables in GIS. We thank Dominique Alò for assistance with statistical assessments in R.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Reiners, W.A.; Reiners, D.S.; Lockwood, J.A. Traits of a good ecologist: What do ecologists think? Ecosphere 2013, 4, 1–22. [Google Scholar] [CrossRef]
- Justice, C.; White, S.M.; McCullough, D.A.; Graves, D.S.; Blanchard, M.R. Can stream and riparian restoration offset climate change impacts to salmon populations? J. Environ. Manag. 2017, 188, 212–227. [Google Scholar] [CrossRef] [PubMed]
- Carlson, A.K.; Taylor, W.W.; Schlee, K.M.; Zorn, T.G.; Infante, D.M. Projected impacts of climate change on stream salmonids with implications for resilience-based management. Ecol. Freshw. Fish 2017, 26, 190–204. [Google Scholar] [CrossRef]
- Santiago, J.M.; García de Jalón, D.; Alonso, C.; Solana, J.; Ribalaygua, J.; Pórtoles, J.; Monjo, R. Brown trout thermal niche and climate change: Expected changes in the distribution of cold-water fish in central Spain. Ecohydrology 2016, 9, 514–528. [Google Scholar] [CrossRef]
- Crawford, S.S.; Muir, A.M. Global introductions of salmon and trout in the genus Oncorhynchus: 1870–2007. Rev. Fish Biol. Fish. 2008, 18, 313–344. [Google Scholar] [CrossRef]
- Arismendi, I.; Penaluna, B.E.; Dunham, J.B.; García de Leaniz, C.; Soto, D.; Fleming, I.A.; Gomez-Uchida, D.; Gajardo, G.; Vargas, P.V.; León-Muñoz, J.; et al. Differential invasion success of salmonids in southern Chile: Patterns and hypotheses. Rev. Fish Biol. Fish. 2014, 24, 919–941. [Google Scholar] [CrossRef]
- Vila, I.; Fuentes, L.; Contreras, M. Peces Límnicos de Chile. Bol. Mus. Nac. Hist. Nat. Chile 1999, 48, 61–75. (In Spanish) [Google Scholar]
- Shelton, J.M.; Weyl, O.L.F.; Impson, N.D.; Dallas, H.F.; Esler, K.J.; Paxton, B.R. Temperature mediates the impact of non-native rainbow trout on native freshwater fishes in South Africa’s Cape Fold Ecoregion. Biol. Invasions 2018, 1–18. [Google Scholar] [CrossRef]
- Penaluna, B.E.; Arismendi, I.; Soto, D. Evidence of Interactive Segregation between Introduced Trout and Native Fishes in Northern Patagonian Rivers, Chile. Trans. Am. Fish. Soc. 2009, 138, 839–845. [Google Scholar] [CrossRef]
- Bøhn, T.; Amundsen, P.A.; Sparrow, A. Competitive exclusion after invasion? Biol. Invasions 2008, 10, 359–368. [Google Scholar] [CrossRef]
- Soto, D.; Arismendi, I.; González, J.; Sanzana, J.; Jara, F.; Jara, C.; Guzman, E.; Lara, A. Southern Chile, trout and salmon country: Invasion patterns and threats for native species. Rev. Chil. Hist. Nat. 2006, 79, 97–117. [Google Scholar] [CrossRef]
- OECD. OECD Environmental Performance Reviews: Chile 2005; OECD: Paris, France, 2005. [Google Scholar]
- Schlaepfer, M.A.; Sax, D.F.; Olden, J.D. The Potential Conservation Value of Non-Native Species. Conserv. Biol. 2011, 25, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; Leach, J.H.; Carlton, J.T.; Secor, C.L. Exotic Species in the Great Lakes: A History of Biotic Crises and Anthropogenic Introductions. J. Great Lakes Res. 1993, 19, 1–54. [Google Scholar] [CrossRef]
- Prichard, C.G.; Bence, J.R. Estimating wounding of lake trout by sea lamprey in the upper Great Lakes: Allowing for changing size-specific patterns. J. Great Lakes Res. 2013, 39, 110–119. [Google Scholar] [CrossRef]
- Freeman, R.; Bowerman, W. Opening Rivers to Trojan Fish The Ecological Dilemma of Dam Removal in the Great Lakes. Conserv. Pract. 2002, 3, 35–39. [Google Scholar] [CrossRef]
- Dodd, H.R.; Hayes, D.B.; Baylis, J.R.; Carl, L.M.; Goldstein, J.D.; McLaughlin, R.L.; Noakes, D.L.G.; Porto, L.M.; Jones, M.L. Low-head Sea Lamprey Barrier Effects on Stream Habitat and Fish Communities in the Great Lakes Basin. J. Great Lakes Res. 2003, 29, 386–402. [Google Scholar] [CrossRef]
- Hauer, C.; Pulg, U.; Gabrielsen, S.E.; Barlaup, B.T. Application of step-backwater modelling for salmonid spawning habitat restoration in Western Norway. Ecohydrology 2015, 8, 1239–1261. [Google Scholar] [CrossRef]
- Person, E.; Bieri, M.; Peter, A.; Schleiss, A.J. Mitigation measures for fish habitat improvement in Alpine rivers affected by hydropower operations. Ecohydrology 2014, 7, 580–599. [Google Scholar] [CrossRef]
- Imholt, C.; Soulsby, C.; Malcolm, I.A.; Gibbins, C.N. Influence of contrasting riparian forest cover on stream temperature dynamics in salmonid spawning and nursery streams. Ecohydrology 2013, 6, 380–392. [Google Scholar] [CrossRef]
- Habit, E.M.; Dyer, B.S.; Vila, I. Estado de conocimiento de los peces dulceacuícolas de Chile. Gayana (Concepción) 2006, 70, 100–113. (In Spanish) [Google Scholar] [CrossRef]
- Orrego, R.; Marshall Adams, S.; Barra, R.; Chiang, G.; Gavilan, J.F. Patterns of fish community composition along a river affected by agricultural and urban disturbance in south-central Chile. Hydrobiologia 2009, 620, 35–46. [Google Scholar] [CrossRef]
- Stehr, A.; Aguayo, M.; Link, O.; Parra, O.; Romero, F.; Alcayaga, H. Modelling the hydrologic response of a mesoscale Andean watershed to changes in land use patterns for environmental planning. Hydrol. Earth Syst. Sci. 2010, 14, 1963–1977. [Google Scholar] [CrossRef] [Green Version]
- García, A.; Jorde, K.; Habit, E.; Caamaño, D.; Parra, O. Downstream environmental effects of dam operations: Changes in habitat quality for native fish species. River Res. Appl. 2011, 27, 312–327. [Google Scholar] [CrossRef]
- Wang, L.; Lyons, J.; Kanehl, P.; Bannerman, R. Impacts of urbanization on stream habitat and fish across multiple spatial scales. Environ. Manag. 2001, 28, 255–266. [Google Scholar] [CrossRef]
- Richter, A.; Kolmes, S.A. Maximum temperature limits for chinook, coho, and chum salmon, and steelhead trout in the Pacific Northwest. Rev. Fish. Sci. 2005, 13, 23–49. [Google Scholar] [CrossRef]
- Ellender, B.R.; Rivers-Moore, N.A.; Coppinger, C.R.; Bellingan, T.A.; Weyl, O.L.F. Towards using thermal stress thresholds to predict salmonid invasion potential. Biol. Invasions 2016, 18, 3513–3525. [Google Scholar] [CrossRef]
- Syrjänen, J. Ecology, Fisheries and Management of Wild Brown Trout Populations in Boreal Inland Waters; University of Jyväskylä: Jyväskylä, Finland, 2010. [Google Scholar]
- Baxter, C.V.; Frissell, C. a.; Hauer, F.R. Geomorphology, logging roads, and the distribution of bull trout spawning in a forested river basin: Implications for management and conservation. Trans. Am. Fish. Soc. 1999, 128, 854–867. [Google Scholar] [CrossRef]
- Steedman, R.J. Effects of experimental clearcut logging on water quality in three small boreal forest lake trout (Salvelinus namaycush) lakes. Can. J. Fish. Aquat. Sci. 2000, 57, 92–96. [Google Scholar] [CrossRef]
- Rosenberger, A.E.; Dunham, J.B.; Neuswanger, J.R.; Railsback, S.F. Legacy effects of wildfire on stream thermal regimes and rainbow trout ecology: an integrated analysis of observation and individual-based models. Freshw. Sci. 2015, 34, 1571–1584. [Google Scholar] [CrossRef]
- Lara, A.; Little, C.; Urrutia, R.; McPhee, J.; Álvarez-Garretón, C.; Oyarzún, C.; Soto, D.; Donoso, P.; Nahuelhual, L.; Pino, M.; et al. Assessment of ecosystem services as an opportunity for the conservation and management of native forests in Chile. For. Ecol. Manag. 2009, 258, 415–424. [Google Scholar] [CrossRef]
- Meador, M.R.; Goldstein, R.M. Assessing water quality at large geographic scales: Relations among land use, water physicochemistry, riparian condition, and fish community structure. Environ. Manag. 2003, 31, 504–517. [Google Scholar] [CrossRef] [PubMed]
- Barton, D.R.; Taylor, W.D.; Biette, R.M. Dimensions of riparian buffer strips required to maintain habitat in southern Ontario streams. North Am. J. Fish. Manag. 1985, 5, 364–378. [Google Scholar] [CrossRef]
- Burt, T.P.; Matchett, L.S.; Goulding, K.W.T.; Webster, C.P.; Haycock, N.E. Denitrification in riparian buffer zones: The role of floodplain hydrology. Hydrol. Process. 1999, 13, 1451–1463. [Google Scholar] [CrossRef]
- Hefting, M.M.; Clement, J.C.; Bienkowski, P.; Dowrick, D.; Guenat, C.; Butturini, A.; Topa, S.; Pinay, G.; Verhoeven, J.T.A. The role of vegetation and litter in the nitrogen dynamics of riparian buffer zones in Europe. Ecol. Eng. 2005, 24, 465–482. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The role of riparian vegetation in protecting and improving chemical water quality in streams. J. Am. Water Resour. Assoc. 2010, 46, 261–277. [Google Scholar] [CrossRef]
- Davies, P.; Nelson, M. Relationships between riparian buffer widths and the effects of logging on stream habitat, invertebrate community composition and fish abundance. Mar. Freshw. Res. 1994, 45, 1289–1305. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Smyth, C.; Boutin, S. Quantitative review of riparian buffer width guidelines from Canada and the United States. J. Environ. Manag. 2004, 70, 165–180. [Google Scholar] [CrossRef]
- Romero, F.I.; Cozano, M.A.; Gangas, R.A.; Naulin, P.I. Zonas ribereñas: Protección, restauración y contexto legal en Chile. Bosque (Valdivia) 2014, 35, 1–2. (In Spanish) [Google Scholar] [CrossRef]
- Habit, E.; Piedra, P.; Ruzzante, D.E.; Walde, S.J.; Belk, M.C.; Cussac, V.E.; Gonzalez, J.; Colin, N. Changes in the distribution of native fishes in response to introduced species and other anthropogenic effects. Glob. Ecol. Biogeogr. 2010, 19, 697–710. [Google Scholar] [CrossRef]
- Lacy, S.N.; Meza, F.J.; Marquet, P.A. Can environmental impact assessments alone conserve freshwater fish biota? Review of the Chilean experience. Environ. Impact Assess. Rev. 2017, 63, 87–94. [Google Scholar] [CrossRef]
- Arismendi, I.; Penaluna, B.; Soto, D. Body condition indices as a rapid assessment of the abundance of introduced salmonids in oligotrophic lakes of southern Chile. Lake Reserv. Manag. 2011, 27, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Milner, N.J.; Elliott, J.M.; Armstrong, J.D.; Gardiner, R.; Welton, J.S.; Ladle, M. The natural control of salmon and trout populations in streams. Fish. Res. 2003, 62, 111–125. [Google Scholar] [CrossRef]
- Valdes-Pineda, R.; Pizarro, R.; Garcia-Chevesich, P.; Valdes, J.B.; Olivares, C.; Vera, M.; Balocchi, F.; Perez, F.; Vallejos, C.; Fuentes, R.; et al. Water governance in Chile: Availability, management and climate change. J. Hydrol. 2014, 519, 2538–2567. [Google Scholar] [CrossRef]
- O’Briain, R.; Shephard, S.; Coghlan, B. River reaches with impaired riparian tree cover and channel morphology have reduced thermal resilience. Ecohydrology 2017, 10, 1–9. [Google Scholar] [CrossRef]
- Arismendi, I.; Gonzáles, J.; Soto, D.; Penaluna, B.E. Piscivory and diet overlap between two non-native fishes in southern Chilean streams. Austral Ecol. 2012, 37, 346–354. [Google Scholar] [CrossRef]
- Montgomery, D.R.; Buffington, J.M. Channel-reach morphology in mountain drainage basins. Bull. Geol. Soc. Am. 1997, 109, 596–611. [Google Scholar] [CrossRef]
- Hitt, N.P.; Angermeier, P.L. Evidence for fish dispersal from spatial analysis of stream network topology. J. N. Am. Benthol. Soc. 2008, 27, 304–320. [Google Scholar] [CrossRef] [Green Version]
- Mao, L.; Ugalde, F.; Iroume, A.; Lacy, S.N. The Effects of Replacing Native Forest on the Quantity and Impacts of In-Channel Pieces of Large Wood in Chilean Streams. River Res. Appl. 2017, 33, 73–88. (In Spanish) [Google Scholar] [CrossRef]
- Arratia F., G. Preferencias de habitat de peces siluriformes de aguas continentales de Chile (Fam. Diplomystidae y Trichomycteridae). Stud. Neotrop. Fauna Environ. 1983, 18, 217–237. [Google Scholar] [CrossRef]
- Roni, P.; Beechie, T.; Pess, G.; Hanson, K. Wood placement in river restoration: fact, fiction, and future direction. Can. J. Fish. Aquat. Sci. 2015, 72, 466–478. [Google Scholar] [CrossRef] [Green Version]
- Pardo, R.; Vila, I.; Capella, J.J. Competitive interaction between introduced rainbow trout and native silverside in a Chilean stream. Environ. Biol. Fishes 2009. [Google Scholar] [CrossRef]
- Jowet, I.G.; Richardson, J.; Boubée, J.A.T. Effects of riparian manipulation on stream communities in small streams: Two case studies. N. Z. J. Mar. Freshw. Res. 2009, 43, 763–774. [Google Scholar] [CrossRef] [Green Version]
- Tschaplinski, P.J.; Pike, R.G. Carnation Creek watershed experiment—long-term responses of coho salmon populations to historic forest practices. Ecohydrology 2017, 10. [Google Scholar] [CrossRef]
- QGIS Development Team QGIS Geographic Information System; Open Source Geospatial Foundation Project; American Scientific Publishers: Stevenson Ranch, CA, USA, 2015.
- Lamothe, K.A.; Allen, R.M.; Winningham, K.; Dennis, C.; Johnson, R.L. Stocking for a trophy bass fishery: searching for size differences among largemouth bass and hybrids in southern Arkansas reservoirs. Lake Reserv. Manag. 2016, 32, 194–207. [Google Scholar] [CrossRef]
- Seber, G.A.F.; Cren, E.D. Le Estimating Population Parameters from Catches Large Relative to the Population. J. Anim. Ecol. 1967, 36, 631. [Google Scholar] [CrossRef]
- R Core Team R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; ISBN 3-900051-07-0. [Google Scholar]
Figure 1.
Box plots of Fulton’s k for Oncorhynchus mykiss sampled across nine headwater sites from Central Chile.
Figure 1.
Box plots of Fulton’s k for Oncorhynchus mykiss sampled across nine headwater sites from Central Chile.
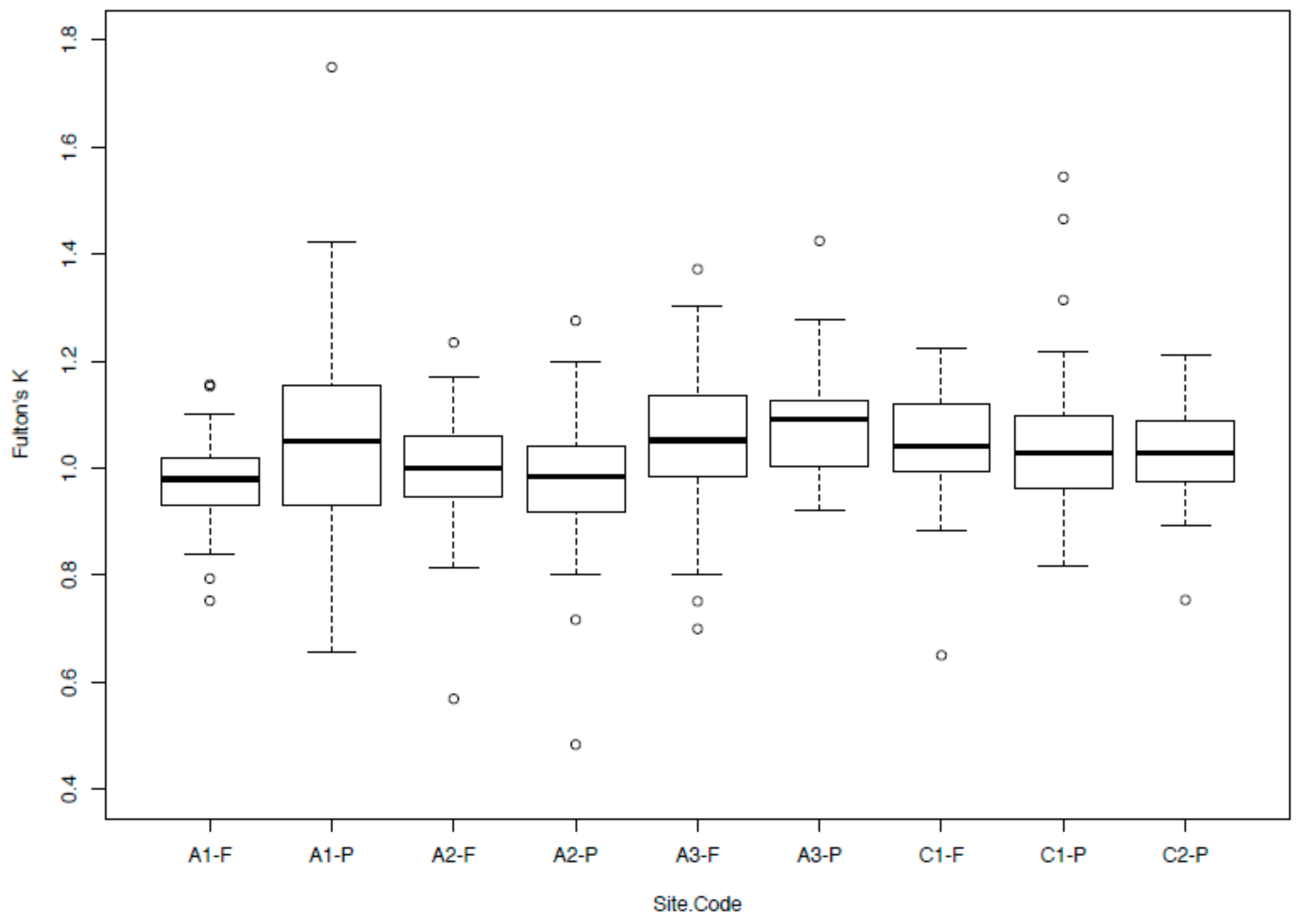
Figure 2.
Scatter plot of catch per unit effort (CPUE) (fish/500 m2) and mean O. mykiss fish condition (k) across nine headwater sites from Central Chile (R2 = 0.521).
Figure 2.
Scatter plot of catch per unit effort (CPUE) (fish/500 m2) and mean O. mykiss fish condition (k) across nine headwater sites from Central Chile (R2 = 0.521).

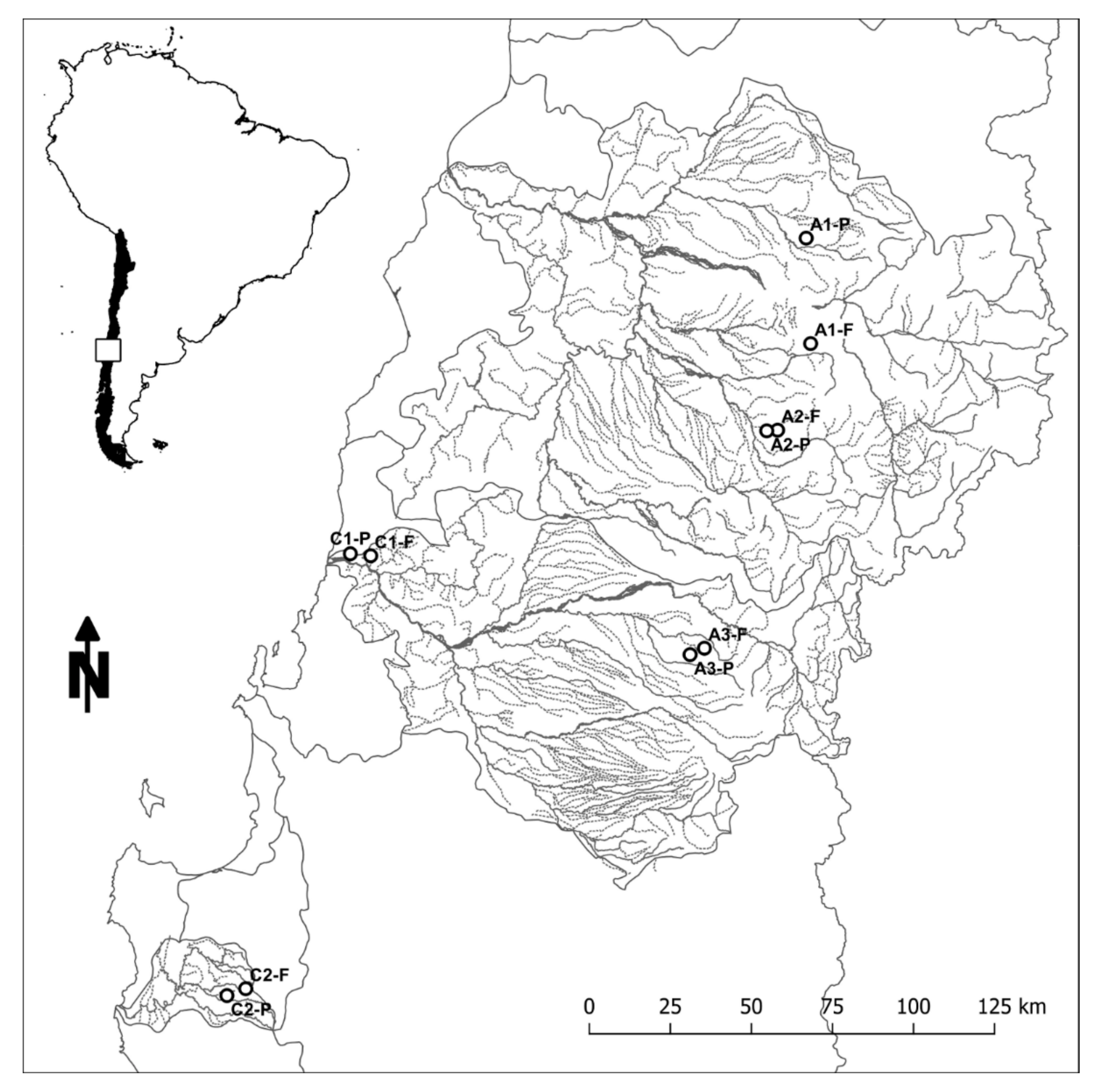
Figure 3.
Paired sampling catchments in the uplands of Central Chile. Two pairs (C1 and C2) are located on the Coastal Range, and three pairs (A1, A2, and A3) are located on the Andean Range. Pair designation is based on dominant land use (F: native forest, P: pine plantation). Study area is indicated by the box in the map insert of South America.
Figure 3.
Paired sampling catchments in the uplands of Central Chile. Two pairs (C1 and C2) are located on the Coastal Range, and three pairs (A1, A2, and A3) are located on the Andean Range. Pair designation is based on dominant land use (F: native forest, P: pine plantation). Study area is indicated by the box in the map insert of South America.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Log-transformed length-weight regression relationships (F: statistic and R2: correlation coefficient), standardised abundance (catch per unit effort (CPUE)), and Fulton’s condition factor (k) for Oncorhynchus mykiss and Nematogenys inermis, organized by sampling sites, paired catchments, land use types, mountain ranges and mountain range × land use (bold: p < 0.0001), where m is slope and b is intercept. CPUE = number of fish/500 m2.
Table 1.
Log-transformed length-weight regression relationships (F: statistic and R2: correlation coefficient), standardised abundance (catch per unit effort (CPUE)), and Fulton’s condition factor (k) for Oncorhynchus mykiss and Nematogenys inermis, organized by sampling sites, paired catchments, land use types, mountain ranges and mountain range × land use (bold: p < 0.0001), where m is slope and b is intercept. CPUE = number of fish/500 m2.
| Species | Grouping | Location | N | F-value | R2 | m | b | CPUE | k |
|---|---|---|---|---|---|---|---|---|---|
| O. mykiss | Site | A1-F | 47 | 2170.67 | 0.98 | 3.056 | −2.08 | 90.7 | 0.97 |
| A1-P | 10 | 747.13 | 0.99 | 2.501 | −1.366 | 26.8 | 1.08 | ||
| A2-F | 34 | 969.78 | 0.97 | 2.736 | −1.722 | 88.3 | 0.99 | ||
| A2-P | 41 | 532.35 | 0.93 | 2.746 | −1.737 | 204.2 | 0.97 | ||
| A3-F | 48 | 420.18 | 0.9 | 2.927 | −1.912 | 33.8 | 1.06 | ||
| A3-P | 24 | 677.33 | 0.97 | 3.039 | −2.009 | 13.6 | 1.10 | ||
| C1-F | 41 | 2668.69 | 0.99 | 2.915 | −1.885 | 43.7 | 1.04 | ||
| C1-P | 42 | 1055.01 | 0.96 | 2.969 | −1.957 | 93.0 | 1.06 | ||
| C2-P | 28 | 718.54 | 0.97 | 2.865 | −1.84 | 26.6 | 1.02 | ||
| Land Use | Forest | 170 | 6264.85 | 0.97 | 2.904 | −1.891 | 64.1 | 1.02 | |
| Plantation | 146 | 3591.47 | 0.96 | 2.837 | −1.81 | 73.9 | 1.02 | ||
| Range | Andes | 205 | 5330.64 | 0.96 | 2.834 | −1.813 | 76.2 | 1.02 | |
| Coast | 111 | 4666.36 | 0.98 | 2.941 | −1.923 | 54.4 | 1.00 | ||
| N. inermis | Sampling Site | A1-P | 3 | - | - | - | - | 12.6 | 0.83 |
| C1-P | 17 | 1275.98 | 0.99 | 3.459 | −2.682 | 21.8 | 0.76 | ||
| Combined | 20 | 1307.6 | 0.99 | 3.378 | −2.581 | 17.2 | 0.77 |
Table 2.
Physical characteristics of the sampled catchment sites.
| Catchment Code | A1-F | A1-P | A2-F | A2-P | A3-F | A3-P | C1-F | C1-P | C2-F | C2-P |
|---|---|---|---|---|---|---|---|---|---|---|
| Location | Andes | Andes | Andes | Andes | Andes | Andes | Coast | Coast | Coast | Coast |
| Catchment area (km2) | 11.86 | 8.68 | 2.35 | 3.53 | 10.21 | 11.22 | 4.92 | 4.83 | 1.43 | 2.54 |
| Max elevation (m) | 2087 | 913 | 1153 | 1078 | 1551 | 1168 | 571 | 558 | 982 | 730 |
| Min elevation (m) | 660 | 452 | 520 | 530 | 565 | 576 | 78 | 75 | 560 | 224 |
| Average slope | 0.086 | 0.016 | 0.047 | 0.046 | 0.043 | 0.015 | 0.08 | 0.029 | 0.168 | 0.073 |
| Native forest (%) | 100 | 32 | 100 | 34 | 98 | 29 | 62 | 25 | 99 | 40 |
| Pine plantation (%) | 0 | 68 | 0 | 66 | 2 | 71 | 38 | 75 | 1 | 60 |
| Water temperature (°C) | 16.5 | 15.6 | 15.4 | 17.1 | 14.6 | 12.8 | 14.1 | 14.1 | 13.2 | 14.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lacy, S.; Ugalde, F.; Mao, L. Invasive Rainbow Trout (Oncorhynchus mykiss) Are Not Affected by Different Land Uses in a Multi-Use, Mediterranean Climate Landscape. Fishes 2018, 3, 37. https://doi.org/10.3390/fishes3040037
AMA Style
Lacy S, Ugalde F, Mao L. Invasive Rainbow Trout (Oncorhynchus mykiss) Are Not Affected by Different Land Uses in a Multi-Use, Mediterranean Climate Landscape. Fishes. 2018; 3(4):37. https://doi.org/10.3390/fishes3040037
Chicago/Turabian StyleLacy, Shaw, Fernando Ugalde, and Luca Mao. 2018. "Invasive Rainbow Trout (Oncorhynchus mykiss) Are Not Affected by Different Land Uses in a Multi-Use, Mediterranean Climate Landscape" Fishes 3, no. 4: 37. https://doi.org/10.3390/fishes3040037