A Field Observation of Rotational Feeding by Neogobius melanostomus

United States Environmental Protection Agency, Office of Research and Development, National Health and Environmental Effects Research Laboratory, Mid-Continent Ecology Division, Duluth, MN 55804, USA
Fishes 2018, 3(1), 5; https://doi.org/10.3390/fishes3010005
Submission received: 19 December 2017
/
Revised: 16 January 2018
/
Accepted: 18 January 2018
/
Published: 22 January 2018
{kind=link}
Abstract
:Neogobius melanostomus, the round goby, was recorded by underwater video feeding on crushed dreissenid mussels at a depth of 12 m in Georgian Bay of Lake Huron, a Laurentian Great Lake. In the video, gobies used rotational or twist feeding to tear away particles from crushed mussels. At least 43 examples of this feeding maneuver occur in the video. Up to 120 gobies m−2 were visible at a time in the video. Mean standard length of gobies appearing in the video was 37 mm. Mean standard length of fish exhibiting twist feeding was larger, 48 mm. Mean size of intact mussels in visible clusters was about 10 × 20 mm, a size which exceeds the gape width of the largest gobies observed in the video. Neogobius melanostomus is known to use twisting to wrest small attached mussels from the substrates which can be crushed by their pharyngeal teeth. I surmise that the behavior observed in the video is an opportunistic manifestation of this inherent behavioral adaptation to overcome gap limitation and exploit a temporary windfall of food.
1. Introduction
Rotational feeding, also known as twist or torsional feeding, is a behavioral maneuver used by some aquatic vertebrates when food items are too large for the animal’s gape, are attached to a substrate, or are too tough to be nipped or sliced away by the feeding animal’s dentition [1]. In this maneuver, the feeding animal grasps the food item in its jaws and rotates or twists its body in relation to the prey item to remove a food particle of a size that can be ingested. The food item must be sufficiently massive or attached to a substrate, so that it has high inertia relative to the feeding animal. Otherwise, the food item will rotate with the twisting predator [2]. This behavior has been observed in crocodilians [3], caecilians [4], and a few groups of fishes including anguillids [5], muraenids [6], and representatives of a few other families [1]. Most fish species for which rotational feeding has been reported are anguilliform or have an attenuated body. One exception is Clinocottos globiceps, a marine cottid, which has been reported to use rotational feeding to tear away anemone tentacles [1].
During a benthic survey of Lake Huron in Canada, I obtained video evidence of this behavior by Neogobius melanostomus Pallas (round goby) feeding on large dreissenid mussels damaged during benthic videography. Neogobius melanostomus is a benthic predator native to Ponto-Caspian Eurasia and is invasive to the Laurentian Great Lakes. This species was first detected in the Great Lakes system in 1990 [7] and is now widespread and abundant in all the Great Lakes except Lake Superior, and in many Great Lakes tributaries [8].
Similar feeding behavior has been observed for N. melanostomus elsewhere in the Great Lakes. Divers observing the species in the St. Clair River, which connects Lakes Huron and Erie, reported that “a fish quickly grabbed the shell of the (zebra) mussel in its jaws and rolled over onto its back thrashing and twisting until the mouthful was wrenched free… and swallowed whole” [9]. In Lake Ontario, “gobies were seen to… consume attached dreissenid mussels, often by grabbing and twisting to loosen individual mussels” [10]. The video and analysis described herein provide additional details about this feeding behavior by N. melanostomus in the Great Lakes.
2. Methods
2.1. Study Area
The video imagery on which this report is based was collected on the morning of 11 August 2017 in north Georgian Bay, Lake Huron, Canada. Ship position was 3 km WSW of the Bustard Islands at 45.87851° N, 80.98847° W. Depth was 12.3 m.
2.2. Video Methods
Video imagery was collected using a SeaViewer 6000 SeaDrop color video camera (SeaViewer Cameras Inc., Tampa, FL, USA) with an external SeaViewer Model 10 LED light. The camera has a frame rate of 60 frames per second at 1080-pixel resolution. The camera and light were mounted “down-looking” within a stainless-steel frame with a square base of 60 × 60 cm and a height (bottom of legs to lifting ring) of 123 cm. The lens of the camera was mounted 62 cm above the substrate. Scale bars with 23 mm increments are visible in the image. A second independent camera and light of the same model were positioned on the outside of the frame for oblique observation. The down-looking and oblique fields of view did not overlap. Unless otherwise indicated, observations described herein are for imagery from the down-looking camera. The field of view at the substrate was 35 × 58 cm. The camera frame was lowered from the aft deck of the United States Environmental Protection Agency’s 28 m long Research Vessel Lake Explorer II using a deck winch, with the winch cable and video cables passing through a block suspended from a crane. Video cables were connected topside to SeaViewer deck consoles with real time display, GPS, clock, user text overlay, and a digital video recorder. Relevant information in the video image overlay includes local time (upper left), ship position (lower left), and date (upper right); other information can be ignored.
2.3. Video Analysis
Windows 10.0 Movie Maker (Version 2012, Microsoft Corp., Redmond, WA, USA) was used to create a 0.25-speed version of the video. The videos were analyzed in playback using Windows 10.0 Media Player (Microsoft Corp., Redmond, WA, USA). Measurements of fish and mussels were made relative to scale bar increments (Video S1, Supplementary Materials). Measurements of dreissenid mussels and N. melanostomus depicted in the video were made to the nearest mm using a caliper held against the display monitor. Maximum shell length of mussels in clusters was difficult to determine from the video, so maximum shell width (mm) was measured. Shell length can be estimated from a length:width ratio of 2, based on selected measurements from the video and previous work [11]. Based on variation in the width of the scale bars in the video image there was about 1 mm of distortion between the center of the image and edge of the image on the vertical axis and about 1.7 mm of distortion on the horizontal axis. I did not attempt to correct for this error in screen measurements. The total length of the video is 133 s; the camera frame was stabilized at 43 s. I measured the standard length (mm) of each fish visible in the paused video at 10, 25, 40, 55 and 70 s from camera stabilization. I measured the standard length of N. melanostomus individuals that displayed rotational feeding whenever it occurred during the video.
3. Results
3.1. Habitat and Dreissenid Mussels
The geologic substrate at the video location was either bedrock or megaclast (particles > 4 m), with occasional cobble (64–256 mm; Video S1, Supplementary Material). Oblique video at the same location was consistent with this characterization of the substrate. A coating of periphyton was visible on the rock surfaces. Aquatic plants were not otherwise present.
Dreissenid mussel clusters (most likely Dreissena rostriformis bugensis Andrusov, quagga mussel [10]) were present along cracks and depressions in the rock surface. Clusters covered about 20% of the substrate between the scale bars in the video image. Mean (±95% confidence intervals) shell width of a sample of intact mussels visible in the video was 10.6 ± 0.6 mm (N = 25). There appeared to be very few small mussels (<5 mm width) in clusters visible in the video.
3.2. Round Goby Size and Behavior
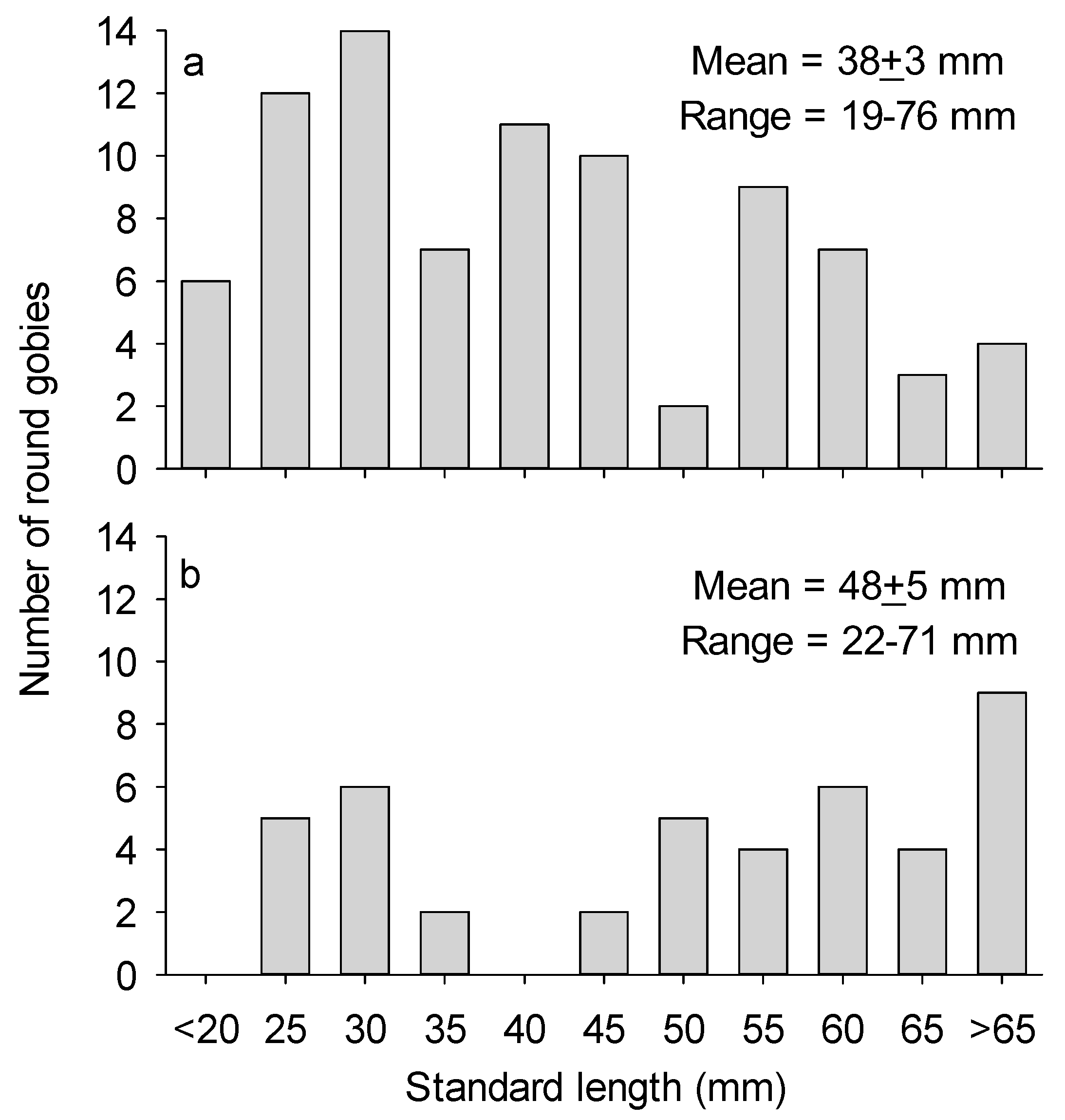
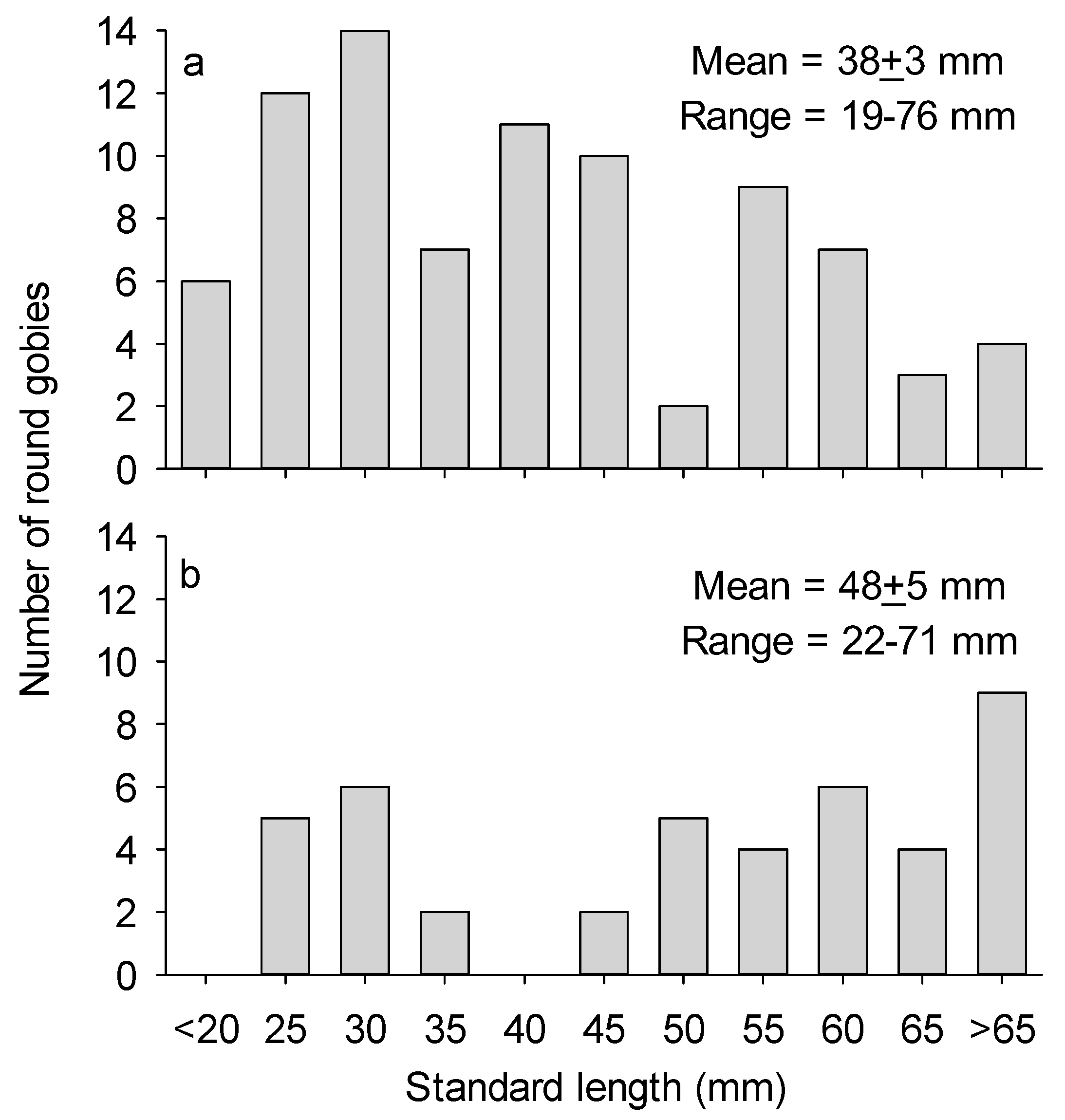
As the video frame was settling on the rock substrates several clusters of mussels were crushed by the legs of the frame, revealing light colored muscle and viscera (Video S1, Supplementary Materials). After the frame had stabilized at t = 11:10:58 (hours:minutes:seconds), the first N. melanostomus appeared in the video after 6 s at t = 11:11:04. Many more individuals swam in and out of view during the video. Figure 1a shows the size distribution of N. melanostomus appearing in the video pooled across the five measurements. Based on length at age relationships for Lake Huron [12], it is likely that four age classes were represented: young of the year and ages 1–3. Mean length of N. melanostomus in the video was 38 ± 3 mm (N = 85). Peak abundance of N. melanostomus observed during the video was about 120 m−2, but fish were likely attracted to the camera lights so this estimate of density is unreliable. No other fish species was observed in the video.
The first example of rotational feeding was observed at t = 11:11:14. In this maneuver, the fish typically grasped tissue still attached to the mussel’s shell and very rapidly twisted its body clockwise or counter-clockwise to detach a particle small enough to ingest. At least 43 examples of this behavior occur in the video. A good example occurs at t = 11:11:34 for a 55-mm round goby. During the twisting action, the fish’s body was typically rotated in a head-down orientation oblique or perpendicular to the substrate surface. From the 0.25-speed version of the video (Video S2, Supplementary Material), it appears that a feeding round goby initially flexes its body from side to side to dislodge a particle of flesh. If that is not immediately successful, the fish violently twists the body at least 180°. The light-colored ventral surface of the fish became visible in most cases. It did not appear that fish achieved complete rotation. Most size classes of gobies were observed to use twist feeding, but the size distribution of fish using this maneuver was skewed toward larger fish (Figure 1b). Mean length of gobies exhibiting rotational feeding was 48 ± 5 mm (N = 43). I did not observe N. melanostomus feeding on intact mussels.
4. Discussion
Dreissenid mussels often form a significant portion of the diet of N. melanostomus in the Great Lakes and across their native range [8,13]. N. melanostomus is able to crush small mussels using molariform pharyngeal teeth and expel the shell fragments [11]. Laboratory feeding trials have shown that N. melanostomus of all sizes prefer smaller mussels [11]. In trials, dreissenid mussels >13 mm in length were not consumed by 100-mm long gobies [11,14], which have a gape width of about 12 mm. Similar size selectivity has also been documented in the laboratory for N. melanostomus fed other bivalve species [15]. Field studies of the species’ diet [16] confirm these laboratory findings. The mean length of mussels visible in the video was about 20 mm, and the largest N. melanostomus individual in the video was about 70 mm standard length. A round goby this size has a gape width of only about 8 mm [14]. Maximum length of dreissenid mussels consumed by N. melanostomus of any size seems to be about 14 mm [17]. I conclude that the round gobies I observed were limited by their gape from consuming mussels at this location until the mussels were crushed. In nature, round gobies may encounter mussels caused by physical disturbance of the substratum or during durophagy by other molluscivores (e.g., fish, birds, crayfish), but the degree to which this impacts the diet of round gobies in the Great Lakes or elsewhere is unknown.
Neogobius melanostomus has been observed in the Great Lakes using torsion to wrench small dreissenid mussels from the substrate, which are swallowed whole [9,10,18]. By exerting torque force with body and head rotations, N. melanostomus can break byssal threads that resist oral jaw bite force [19]. Though they would be prevented by gape limitation from exploiting intact attached mussels at the video location, round gobies immediately and repeatedly exhibited behavior analogous to that used by other gape-limited predators to scavenge food from high inertia food sources such as fish carcass [5]. My unique video observations support the hypothesis that rotational or “twist” feeding used by round gobies to forage on mussels “is apparently a modification of rotational feeding in which a predator spins to tear chunks from prey too large to swallow whole [20]”.
I think it most likely that the rotational feeding behavior I documented for N. melanostomus feeding on the tissue of crushed mussels is a manifestation of an inherent behavioral adaptation for feeding on attached mussels [20], including in their native range [15]. It should be noted however, that round gobies have been observed feeding on fish carcasses at least once [21], so the origin of rotational feeding in the species (small attached mussels or other high inertia food sources) is uncertain. The behavioral propensity of N. melanostomus to aggressively twist their bodies (even when held in the hand) is absent in other benthic fish native to the Laurentian Great Lakes (e.g., Cottidae, Etheostomatinae) [20]. Rotational feeding allows N. melanostomus to both consume attached dreissenid mussels and to overcome gape limitation when opportunistically feeding on other food sources. I surmise this flexibility in feeding behavior is among the factors accounting for the success of N. melanostomus as an invader.
Supplementary Materials
Video S1: Full speed video of feeding round gobies; Video S2: 1/4 speed video of feeding round gobies. Links: Video S1 https://zenodo.org/record/1157438#.WmhXvKjiaUl; Video S2 https://zenodo.org/record/1157438#.WmhXvKjiaUl.
Acknowledgments
I thank the scientists and ship’s crew of the US Environmental Protection Agency’s Research Vessel Lake Explorer II. Comments by Mark Vinson, Barb Sheedy, Molly Wick, Carolyn Foley, and Matt Pawlowski improved the paper. The views expressed in this paper are the author’s and do not necessarily reflect the views or policies of the U.S. Government. Mention of trade names does not constitute endorsement or recommendation for use.
Conflicts of Interest
The author declares no conflict of interest.
References
- Helfman, G.S. Mode selection and mode switching in foraging animals. Adv. Stud. Behav. 1990, 19, 249–298. [Google Scholar]
- Taylor, M.A. How tetrapods feed in water: A functional analysis by paradigm. Zool. J. Linn. Soc. 1987, 91, 171–195. [Google Scholar] [CrossRef]
- Fish, F.E.; Bostic, S.A.; Nicastro, A.J.; Beneski, J.T. Death roll of the alligator: Mechanics of twist feeding in water. J. Exp. Biol. 2007, 210, 2811–2818. [Google Scholar] [CrossRef] [PubMed]
- Measey, G.J.; Herrel, A. Rotational feeding in caecilians: Putting a spin on the evolution of cranial design. Biol. Lett. 2006, 2, 485–487. [Google Scholar] [CrossRef] [PubMed]
- Helfman, G.S.; Clark, J.B. Rotational feeding: Overcoming gape-limited foraging in anguillid eels. Copeia 1986, 3, 679–685. [Google Scholar] [CrossRef]
- Miller, T.J. Feeding behavior of Echidna nebulosa, Enchelycore pardalis, and Gymnomuraena zebra (Teleostei: Muraenidae). Copeia 1989, 3, 662–672. [Google Scholar] [CrossRef]
- Jude, D.J.; Reider, R.H.; Smith, G.R. Establishment of Gobiidae in the Great Lakes basin. Can. J. Fish. Aquat. Sci. 1992, 49, 416–421. [Google Scholar] [CrossRef]
- Kornis, M.S.; Mercado-Silva, N.; Vander Zanden, M.J. Twenty years of invasion: A review of round goby Neogobius melanostomus biology, spread and ecological implications. J. Fish Biol. 2012, 80, 235–285. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Lashbrook, G. Two New Fish Aliens in the Great Lakes; Michigan Natural Resources: Lansing, MI, USA, 1993; pp. 17–19.
- Wilson, K.A.; Howell, E.T.; Jackson, D.A. Replacement of zebra mussels by quagga mussels in the Canadian nearshore of Lake Ontario: The importance of substrate, round goby abundance, and upwelling frequency. J. Great Lakes Res. 2006, 32, 11–28. [Google Scholar] [CrossRef]
- Ghedotti, M.J.; Smihula, J.C.; Smith, G.R. Zebra mussel predation by round gobies in the laboratory. J. Great Lakes Res. 1995, 21, 665–669. [Google Scholar] [CrossRef]
- French, J.R., III; Black, M.G. Maximum length and age of round gobies (Apollonia melanostomus) in Lake Huron. J. Freshw. Ecol. 2009, 24, 173–175. [Google Scholar] [CrossRef]
- Charlebois, P.M.; Marsden, J.E.; Goettel, R.G.; Wolfe, R.K.; Jude, D.J. The Round Goby, Neogobius melanostomus (Pallas): A Review of European and North American Literature; Illinois Natural History Survey Special Publication: Champaign, IL, USA, 1997. [Google Scholar]
- Ray, W.J.; Corkum, L.D. Predation of zebra mussels by round gobies, Neogobius melanostomus. Environ. Biol. Fishes 1997, 50, 267–273. [Google Scholar] [CrossRef]
- Schrandt, M.N.; Stone, L.C.; Klimek, B.; Mäkelin, S.; Heck, K.L.; Mattila, J.; Herlevi, H. A laboratory study of potential effects of the invasive round goby on nearshore fauna of the Baltic Sea. Aquat. Invasions 2016, 11, 327–335. [Google Scholar] [CrossRef]
- Andraso, G.M.; Ganger, M.T.; Adamczyk, J. Size-selective predation by round gobies (Neogobius melanostomus) on dreissenid mussels in the field. J. Great Lakes Res. 2011, 37, 298–304. [Google Scholar] [CrossRef]
- Kipp, R.; Hébert, I.; Lacharité, M.; Ricciardi, A. Impacts of predation by the Eurasian round goby (Neogobius melanostomus) on mollusks in the upper St. Lawrence River. J. Great Lakes Res. 2012, 38, 78–89. [Google Scholar] [CrossRef]
- Djuricich, P.; Janssen, J. Impact of round goby predation on zebra mussel size distribution at Calumet Harbor, Lake Michigan. J. Great Lakes Res. 2001, 27, 312–318. [Google Scholar] [CrossRef]
- Morano, J.L. Ecomorphology of Round Goby, Neogobius melanostomus, Predation on Zebra Mussels, Dreissena polymorpha. Masters’s Thesis, Loyola University, Chicago, IL, USA, 2007. [Google Scholar]
- Houghton, C.J.; Janssen, J. Variation in predator-prey interactions between round gobies and dreissenid mussels. In Quagga and Zebra Mussels Biology, Impacts, and Control, 2nd ed.; Nalepa, T., Schlosser, D., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 359–367. [Google Scholar]
- Polačik, M.; Jurajda, P.; Blažek, R.; Janáč, M. Carcass feeding as a cryptic foraging mode in round goby, Neogobius melanostomus. J. Fish Biol. 2015, 87, 194–199. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plot (a): length-frequency distribution of N. melanostomus appearing in the video. Plot (b): length-frequency distribution of N. melanostomus exhibiting rotational feeding in the video; some fish were observed exhibiting this behavior more than once.
Figure 1.
Plot (a): length-frequency distribution of N. melanostomus appearing in the video. Plot (b): length-frequency distribution of N. melanostomus exhibiting rotational feeding in the video; some fish were observed exhibiting this behavior more than once.
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Angradi, T.R. A Field Observation of Rotational Feeding by Neogobius melanostomus. Fishes 2018, 3, 5. https://doi.org/10.3390/fishes3010005
AMA Style
Angradi TR. A Field Observation of Rotational Feeding by Neogobius melanostomus. Fishes. 2018; 3(1):5. https://doi.org/10.3390/fishes3010005
Chicago/Turabian StyleAngradi, Ted R. 2018. "A Field Observation of Rotational Feeding by Neogobius melanostomus" Fishes 3, no. 1: 5. https://doi.org/10.3390/fishes3010005