
Bacterial and Fungal Endophytes of Grapevine Cultivars Growing in Primorsky Krai of Russia

 , , , , , and
, , , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Collection and Pre-Treatment
2.2. DNA Extraction and Illumina MiSeq Sequencing
2.3. Bioinformatics and Biostatistics
3. Results
3.1. Illumina Next-Generation Sequencing Results
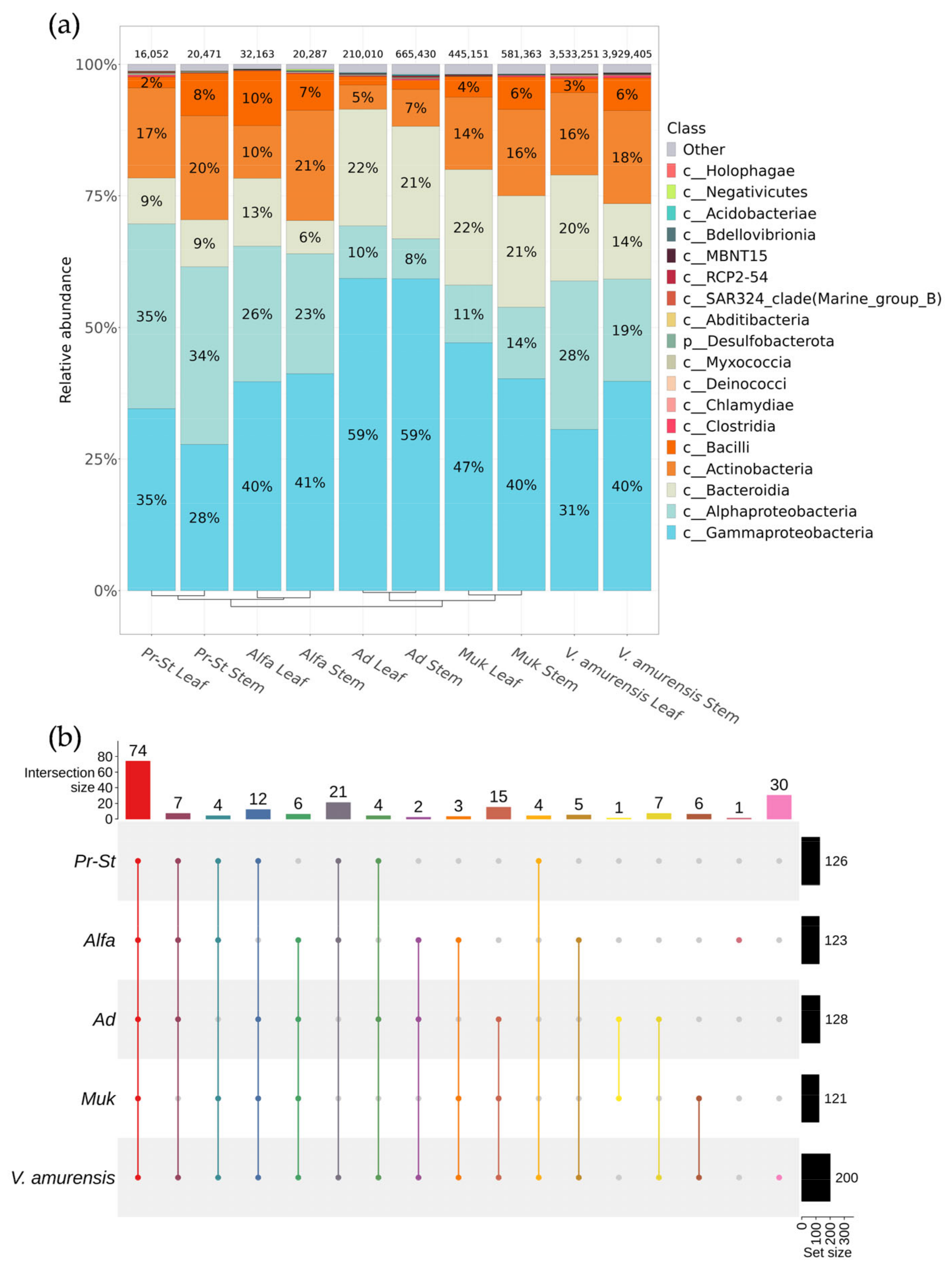
3.2. The Biodiversity of Bacterial Endophytes from Grapevine Cultivars in Primorsky Krai of Russia
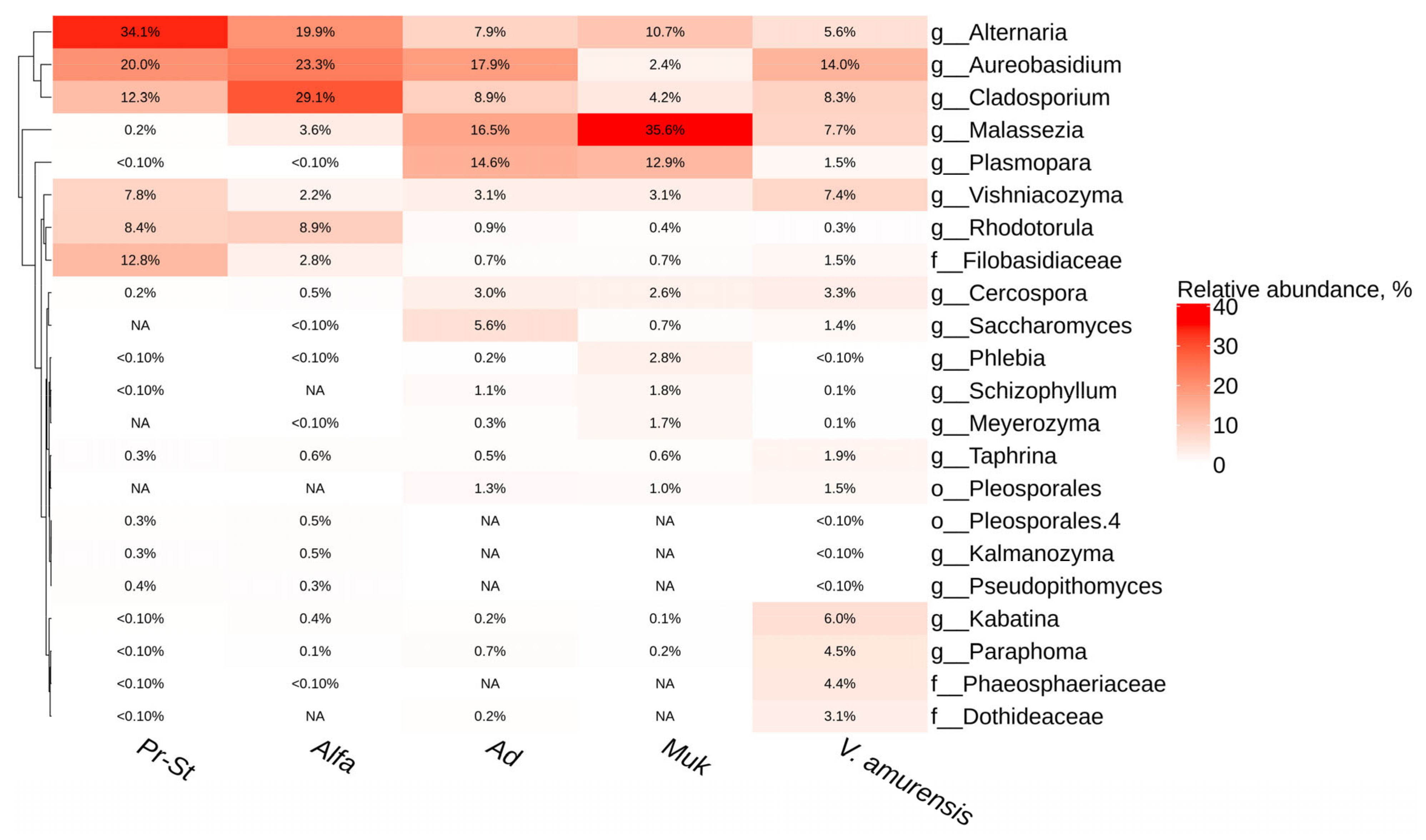
3.3. The Fungal and Fungi-like Endophytic Microorganisms from Grapevine Varietes in Primorsky Krai of Russia
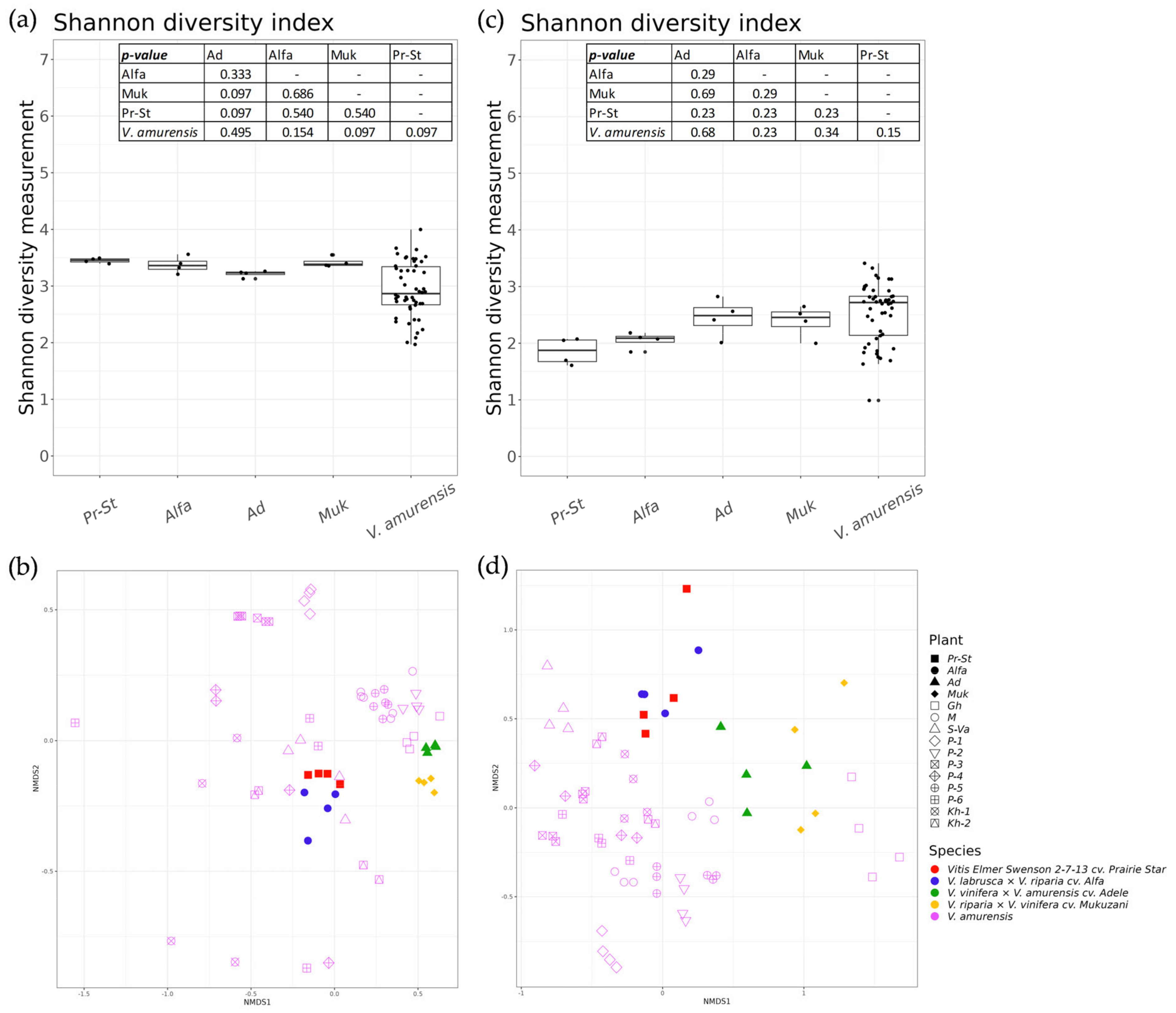
3.4. A Comparative Analysis of the Endophytic Microbial Communities from Cultivated and Wild Grapevine Varietes
3.5. Analysis of Associations of Endophytic Bacteria and Fungi in the Vitis Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Felber, A.C.; Orlandelli, R.C.; Rhoden, S.A.; Garcia, A.; Costa, A.T.; Azevedo, J.L.; Pamphile, J.A. Bioprospecting Foliar Endophytic Fungi of Vitis Labrusca Linnaeus, Bordô and Concord Cv. Ann. Microbiol. 2016, 66, 765–775. [Google Scholar] [CrossRef]
- Soejima, A.; Wen, J. Phylogenetic Analysis of the Grape Family (Vitaceae) Based on Three Chloroplast Markers. Am. J. Bot. 2006, 93, 278–287. [Google Scholar] [CrossRef]
- Compant, S.; Brader, G.; Muzammil, S.; Sessitsch, A.; Lebrihi, A.; Mathieu, F. Use of Beneficial Bacteria and Their Secondary Metabolites to Control Grapevine Pathogen Diseases. BioControl 2013, 58, 435–455. [Google Scholar] [CrossRef]
- Swift, J.F.; Hall, M.E.; Harris, Z.N.; Kwasniewski, M.T.; Miller, A.J. Grapevine Microbiota Reflect Diversity among Compartments and Complex Interactions within and among Root and Shoot Systems. Microorganisms 2021, 9, 92. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.I.; Elfar, K.; Eskalen, A. Evaluation of the Antifungal Activity of Endophytic and Rhizospheric Bacteria against Grapevine Trunk Pathogens. Microorganisms 2022, 10, 2035. [Google Scholar] [CrossRef]
- Jiao, S.; Zeng, F.; Huang, Y.; Zhang, L.; Mao, J.; Chen, B. Physiological, Biochemical and Molecular Responses Associated with Drought Tolerance in Grafted Grapevine. BMC Plant Biol. 2023, 23, 110. [Google Scholar] [CrossRef]
- Ren, C.; Fan, P.; Li, S.; Liang, Z. Advances in Understanding Cold Tolerance in Grapevine. Plant Physiol. 2023, 192, 1733–1746. [Google Scholar] [CrossRef]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A Critical Review of Plant Protection Tools for Reducing Pesticide Use on Grapevine and New Perspectives for the Implementation of IPM in Viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Carvalho, F.P. Pesticides, Environment, and Food Safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Knapp, D.G.; Lázár, A.; Molnár, A.; Vajna, B.; Karácsony, Z.; Váczy, K.Z.; Kovács, G.M. Above-Ground Parts of White Grapevine Vitis vinifera cv. Furmint Share Core Members of the Fungal Microbiome. Environ. Microbiol. Rep. 2021, 13, 509–520. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Kiselev, K.V. Interaction of Plants and Endophytic Microorganisms: Molecular Aspects, Biological Functions, Community Composition, and Practical Applications. Plants 2023, 12, 714. [Google Scholar] [CrossRef] [PubMed]
- Hamaoka, K.; Aoki, Y.; Suzuki, S. Isolation and Characterization of Endophyte Bacillus velezensis KOF112 from Grapevine Shoot Xylem as Biological Control Agent for Fungal Diseases. Plants 2021, 10, 1815. [Google Scholar] [CrossRef] [PubMed]
- Mikheev, V.S.; Struchkova, I.V.; Ageyeva, M.N.; Brilkina, A.A.; Berezina, E.V. The Role of Phialocephala fortinii in Improving Plants’ Phosphorus Nutrition: New Puzzle Pieces. J. Fungi 2022, 8, 1225. [Google Scholar] [CrossRef]
- Ma, Y.; Jiao, J.; Fan, X.; Sun, H.; Zhang, Y.; Jiang, J.; Liu, C. Endophytic Bacterium Pseudomonas fluorescens RG11 May Transform Tryptophan to Melatonin and Promote Endogenous Melatonin Levels in the Roots of Four Grape Cultivars. Front. Plant Sci. 2017, 7, 2608. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant Growth-Promoting Bacteria That Confer Resistance to Water Stress in Tomatoes and Peppers. Plant Sci. 2004, 166, 525–530. [Google Scholar] [CrossRef]
- Theocharis, A.; Bordiec, S.; Fernandez, O.; Paquis, S.; Dhondt-Cordelier, S.; Baillieul, F.; Clément, C.; Barka, E.A. Burkholderia phytofirmans PsJN Primes Vitis Vinifera L. and Confers a Better Tolerance to Low Nonfreezing Temperatures. MPMI 2012, 25, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among Wine Grape Microbiome, Metabolome, and Fermentation Behavior Suggest Microbial Contribution to Regional Wine Characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef]
- Novello, G.; Gamalero, E.; Bona, E.; Boatti, L.; Mignone, F.; Massa, N.; Cesaro, P.; Lingua, G.; Berta, G. The Rhizosphere Bacterial Microbiota of Vitis vinifera cv. Pinot Noir in an Integrated Pest Management Vineyard. Front. Microbiol. 2017, 8, 1528. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, D.; Squartini, A.; Crucitti, D.; Barizza, E.; Lo Schiavo, F.; Muresu, R.; Carimi, F.; Zottini, M. The Role of the Endophytic Microbiome in the Grapevine Response to Environmental Triggers. Front. Plant Sci. 2019, 10, 1256. [Google Scholar] [CrossRef] [PubMed]
- Aleynova, O.A.; Nityagovsky, N.N.; Dubrovina, A.S.; Kiselev, K.V. The Biodiversity of Grapevine Bacterial Endophytes of Vitis amurensis Rupr. Plants 2022, 11, 1128. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Nityagovsky, N.N.; Suprun, A.R.; Ananev, A.A.; Dubrovina, A.S.; Kiselev, K.V. The Diversity of Fungal Endophytes from Wild Grape Vitis amurensis Rupr. Plants 2022, 11, 2897. [Google Scholar] [CrossRef] [PubMed]
- Kiselev, K.V.; Nityagovsky, N.N.; Aleynova, O.A. A Method of DNA Extraction from Plants for Metagenomic Analysis Based on the Example of Grape Vitis amurensis Rupr. Appl. Biochem. Microbiol. 2023, 59, 361–367. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.H.; Kõljalg, U. UNITE QIIME Release for Eukaryotes; Version 29.11.2022; UNITE Community: London, UK, 2022. [Google Scholar]
- Bisanz, J. qiime2R: Importing QIIME2 Artifacts and Associated Data into R Sessions. 2018. Available online: https://github.com/jbisanz/qiime2R (accessed on 18 October 2023).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- De Vries, A.; Ripley, B.D. ggdendro: Create Dendrograms and Tree Diagrams Using “ggplot2”. 2023. Available online: https://andrie.github.io/ggdendro/ (accessed on 18 October 2023).
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes. 2022. Available online: https://cran.r-project.org/web/packages/RColorBrewer/ (accessed on 18 October 2023).
- Gu, Z. Circlize: Circular Visualization. 2022. Available online: https://cran.r-project.org/web/packages/circlize/ (accessed on 18 October 2023).
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Pedersen, T.L. Tidygraph: A Tidy API for Graph Manipulation. 2023. Available online: https://tidygraph.data-imaginist.com/ (accessed on 18 October 2023).
- Pedersen, T.L. ggraph: An Implementation of Grammar of Graphics for Graphs and Networks. 2022. Available online: https://ggraph.data-imaginist.com/ (accessed on 18 October 2023).
- Peschel, S.; Müller, C.L.; von Mutius, E.; Boulesteix, A.-L.; Depner, M. NetCoMi: Network Construction and Comparison for Microbiome Data in R. Brief. Bioinform. 2021, 22, bbaa290. [Google Scholar] [CrossRef]
- Kurtz, Z.D.; Müller, C.L.; Miraldi, E.R.; Littman, D.R.; Blaser, M.J.; Bonneau, R.A. Sparse and Compositionally Robust Inference of Microbial Ecological Networks. PLoS Comput. Biol. 2015, 11, e1004226. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 18 October 2023).
- Gu, Z.; Eils, R.; Schlesner, M. Complex Heatmaps Reveal Patterns and Correlations in Multidimensional Genomic Data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Nityagovsky, N.N.; Ananev, A.A.; Suprun, A.R.; Ogneva, Z.V.; Dneprovskaya, A.A.; Beresh, A.A.; Tyunin, A.P.; Dubrovina, A.S.; Kiselev, K.V. The Endophytic Microbiome of Wild Grapevines Vitis amurensis Rupr. and Vitis coignetiae Pulliat Growing in the Russian Far East. Plants 2023, 12, 2952. [Google Scholar] [CrossRef]
- Hamaoka, K.; Aoki, Y.; Takahashi, S.; Enoki, S.; Yamamoto, K.; Tanaka, K.; Suzuki, S. Diversity of Endophytic Bacterial Microbiota in Grapevine Shoot Xylems Varies Depending on Wine Grape-Growing Region, Cultivar, and Shoot Growth Stage. Sci. Rep. 2022, 12, 15772. [Google Scholar] [CrossRef]
- Nanetti, E.; Palladino, G.; Scicchitano, D.; Trapella, G.; Cinti, N.; Fabbrini, M.; Cozzi, A.; Accetta, G.; Tassini, C.; Iannaccone, L.; et al. Composition and Biodiversity of Soil and Root-Associated Microbiome in Vitis vinifera Cultivar Lambrusco Distinguish the Microbial Terroir of the Lambrusco DOC Protected Designation of Origin Area on a Local Scale. Front. Microbiol. 2023, 14, 1108036. [Google Scholar] [CrossRef]
- Nigris, S.; Baldan, E.; Tondello, A.; Zanella, F.; Vitulo, N.; Favaro, G.; Guidolin, V.; Bordin, N.; Telatin, A.; Barizza, E.; et al. Biocontrol Traits of Bacillus licheniformis GL174, a Culturable Endophyte of Vitis vinifera cv. Glera. BMC Microbiol. 2018, 18, 133. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, W.; Li, C.; Wang, H.; Wang, H.; Ling, Y.; Yan, G.; Chang, Y. Effects of Antibiotics on Corncob Supported Solid-Phase Denitrification: Denitrification and Antibiotics Removal Performance, Mechanism, and Antibiotic Resistance Genes. J. Environ. Sci. 2023, 130, 24–36. [Google Scholar] [CrossRef]
- Wang, Y.; Li, D.; Liu, M.; Xia, C.; Fan, Q.; Li, X.; Lan, Z.; Shi, G.; Dong, W.; Li, Z.; et al. Preparation of Active Chitooligosaccharides with a Novel Chitosanase AqCoA and Their Application in Fungal Disease Protection. J. Agric. Food Chem. 2021, 69, 3351–3361. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Jung, J.-H.; Kim, M.-K.; Choe, H.N.; Seong, C.N.; Lim, S. Spirosoma taeanense sp. nov., a Radiation Resistant Bacterium Isolated from a Coastal Sand Dune. Antonie Van Leeuwenhoek 2021, 114, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Maeng, S.; Damdintogtokh, T.; Zhang, J.; Kim, M.-K.; Srinivasan, S.; Kim, M.K. Spirosoma profusum sp. nov., and Spirosoma validum sp. nov., Radiation-Resistant Bacteria Isolated from Soil in South Korea. Antonie Van Leeuwenhoek 2021, 114, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, X.-Y.; Su, X.-J.; Hu, Q.; Jiang, J.-D. Spirosoma sordidisoli sp. nov., a Propanil-Degrading Bacterium Isolated from a Herbicide-Contaminated Soil. Antonie Van Leeuwenhoek 2019, 112, 1523–1532. [Google Scholar] [CrossRef]
- Wassermann, B.; Korsten, L.; Berg, G. Plant Health and Sound Vibration: Analyzing Implications of the Microbiome in Grape Wine Leaves. Pathogens 2021, 10, 63. [Google Scholar] [CrossRef]
- Zhu, L.; Li, T.; Xu, X.; Shi, X.; Wang, B. Succession of Fungal Communities at Different Developmental Stages of Cabernet Sauvignon Grapes From an Organic Vineyard in Xinjiang. Front. Microbiol. 2021, 12, 718261. [Google Scholar] [CrossRef] [PubMed]
- Olmo, H.P. Vinifera rotundifolia Hybrids as Wine Grapes. Am. J. Enol. Vitic. 1971, 22, 87–91. [Google Scholar] [CrossRef]
- Staudt, G.; Kassemeyer, H. Evaluation of Downy Mildew Resistance in Various Accessions of Wild Vitis Species. Vitis 1995, 34, 225–228. [Google Scholar]
- Díez-Navajas, A.M.; Wiedemann-Merdinoglu, S.; Greif, C.; Merdinoglu, D. Nonhost Versus Host Resistance to the Grapevine Downy Mildew, Plasmopara viticola, Studied at the Tissue Level. Phytopathology 2008, 98, 776–780. [Google Scholar] [CrossRef] [PubMed]
- Massi, F.; Torriani, S.F.F.; Borghi, L.; Toffolatti, S.L. Fungicide Resistance Evolution and Detection in Plant Pathogens: Plasmopara viticola as a Case Study. Microorganisms 2021, 9, 119. [Google Scholar] [CrossRef]
- Toffolatti, S.L.; Serrati, L.; Sierotzki, H.; Gisi, U.; Vercesi, A. Assessment of QoI Resistance in Plasmopara viticola Oospores. Pest Manag. Sci. 2007, 63, 194–201. [Google Scholar] [CrossRef]
- Huang, X.; Wang, X.; Kong, F.; van der Lee, T.; Wang, Z.; Zhang, H. Detection and Characterization of Carboxylic Acid Amide-Resistant Plasmopara viticola in China Using a TaqMan-MGB Real-Time PCR. Plant Dis. 2020, 104, 2338–2345. [Google Scholar] [CrossRef]
- Tournas, V.H.; Katsoudas, E. Mould and Yeast Flora in Fresh Berries, Grapes and Citrus Fruits. Int. J. Food Microbiol. 2005, 105, 11–17. [Google Scholar] [CrossRef]
- Ding, Y.; Wei, R.; Wang, L.; Wang, W.; Wang, H.; Li, H. Exploring the Ecological Characteristics of Natural Microbial Communities along the Continuum from Grape Berries to Winemaking. Food Res. Int. 2023, 167, 112718. [Google Scholar] [CrossRef]
- Shi, J.; Huang, D.; Du, Y.; Zhu, S.; Hussain, Z.; Haider, M.S.; Anwar, R. Effects of Exogenous Nitric Oxide Treatment on Grape Berries Against Botrytis cinerea and Alternaria alternata Related Enzymes and Metabolites. Plant Dis. 2023, 107, 1510–1521. [Google Scholar] [CrossRef]
- Yu, X.; Shentu, X.; Dong, S.; Hao, P.; Bian, Y.; Ma, Z. Application of Metabolites of Alternaria alternata 31 in Preventing and Treating Rhizoctonia solani, Fusarium oxysporium, and Botrytis cinerea. CN Patent 102,204,570 A, 5 October 2011. [Google Scholar]
- Ortega, H.E.; Torres-Mendoza, D.; Cubilla-Rios, L. Patents on Endophytic Fungi for Agriculture and Bio- and Phytoremediation Applications. Microorganisms 2020, 8, 1237. [Google Scholar] [CrossRef] [PubMed]
- Grube, M.; Schmid, F.; Berg, G. Black Fungi and Associated Bacterial Communities in the Phyllosphere of Grapevine. Fungal Biol. 2011, 115, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Lu, Y.; Chi, Z.; Liu, G.-L.; Jiang, H.; Hu, Z.; Chi, Z.-M. Genome Editing of Different Strains of Aureobasidium melanogenum Using an Efficient Cre/Loxp Site-Specific Recombination System. Fungal Biol. 2019, 123, 723–731. [Google Scholar] [CrossRef]
- Longhi, S.J.; Martín, M.C.; Merín, M.G.; Morata De Ambrosini, V.I. Yeast Multi-Enzymatic Systems for Improving Colour Extraction, Technological Parameters and Antioxidant Activity of Wine. Food Technol. Biotechnol. 2022, 60, 556–570. [Google Scholar] [CrossRef]
- Martins, G.; Vallance, J.; Mercier, A.; Albertin, W.; Stamatopoulos, P.; Rey, P.; Lonvaud, A.; Masneuf-Pomarède, I. Influence of the Farming System on the Epiphytic Yeasts and Yeast-like Fungi Colonizing Grape Berries during the Ripening Process. Int. J. Food Microbiol. 2014, 177, 21–28. [Google Scholar] [CrossRef]
- Yu, J.; Wang, X.-W.; Liu, S.-L.; Shen, S.; Zhou, L.-W. Taxonomy and Phylogeny of Resinicium sensu lato from Asia-Pacific Revealing a New Genus and Five New Species (Hymenochaetales, Basidiomycota). IMA Fungus 2021, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Agrios, G. Plant Pathology, 4th ed.; Academic Press: London, UK, 1997; Volume 144, p. 146. [Google Scholar]
- Al-Sadi, A.M. Bipolaris Sorokiniana-Induced Black Point, Common Root Rot, and Spot Blotch Diseases of Wheat: A Review. Front. Cell. Infect. Microbiol. 2021, 11, 584899. [Google Scholar] [CrossRef]
- Purnomo, A.S.; Kamei, I.; Kondo, R. Degradation of 1,1,1-Trichloro-2,2-Bis (4-Chlorophenyl) Ethane (DDT) by Brown-Rot Fungi. J. Biosci. Bioeng. 2008, 105, 614–621. [Google Scholar] [CrossRef]
- Soliman, S.S.M.; Greenwood, J.S.; Bombarely, A.; Mueller, L.A.; Tsao, R.; Mosser, D.D.; Raizada, M.N. An Endophyte Constructs Fungicide-Containing Extracellular Barriers for Its Host Plant. Curr. Biol. 2015, 25, 2570–2576. [Google Scholar] [CrossRef]
- Agler, M.T.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.-T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Palaniyandi, S.A.; Yang, S.H.; Zhang, L.; Suh, J.-W. Effects of Actinobacteria on Plant Disease Suppression and Growth Promotion. Appl. Microbiol. Biotechnol. 2013, 97, 9621–9636. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleynova, O.A.; Nityagovsky, N.N.; Ananev, A.A.; Suprun, A.R.; Ogneva, Z.V.; Dneprovskaya, A.A.; Beresh, A.A.; Dubrovina, A.S.; Chebukin, P.A.; Kiselev, K.V. Bacterial and Fungal Endophytes of Grapevine Cultivars Growing in Primorsky Krai of Russia. Horticulturae 2023, 9, 1257. https://doi.org/10.3390/horticulturae9121257
Aleynova OA, Nityagovsky NN, Ananev AA, Suprun AR, Ogneva ZV, Dneprovskaya AA, Beresh AA, Dubrovina AS, Chebukin PA, Kiselev KV. Bacterial and Fungal Endophytes of Grapevine Cultivars Growing in Primorsky Krai of Russia. Horticulturae. 2023; 9(12):1257. https://doi.org/10.3390/horticulturae9121257
Chicago/Turabian StyleAleynova, Olga A., Nikolay N. Nityagovsky, Alexey A. Ananev, Andrey R. Suprun, Zlata V. Ogneva, Alina A. Dneprovskaya, Alina A. Beresh, Alexandra S. Dubrovina, Pavel A. Chebukin, and Konstantin V. Kiselev. 2023. "Bacterial and Fungal Endophytes of Grapevine Cultivars Growing in Primorsky Krai of Russia" Horticulturae 9, no. 12: 1257. https://doi.org/10.3390/horticulturae9121257