
Changes in the Polyphenol Content of Red Raspberry Fruits during Ripening


1
Graduate School of Engineering, Osaka Electro-Communication University (OECU), 18-8 Hatsu-cho, Neyagawa-shi 572-8530, Osaka, Japan
2
Research Institute for Sustainable Humanosphere, Kyoto University, Uji 611-0011, Kyoto, Japan
*
Author to whom correspondence should be addressed.
Horticulturae 2021, 7(12), 569; https://doi.org/10.3390/horticulturae7120569
Submission received: 26 October 2021
/
Revised: 6 December 2021
/
Accepted: 7 December 2021
/
Published: 10 December 2021
(This article belongs to the Special Issue Bioactivity, Phytochemical and Nutritional Composition of Natural Products)
Abstract
:Berry fruits that contain large amounts of polyphenol compounds are expected to exhibit health and anti-aging effects due to the antioxidant activities of these components. Among the various polyphenols, flavan-3-ol derivatives are known to have a particularly high functionality. In this study, the maturity of red raspberry fruits is classified into eight stages based on the polyphenol content at each stage. Quantification of the various compounds and investigation of the DPPH and ABTS radical scavenging activities were carried out. The total polyphenol content, including that of the flavan-3-ol derivatives, was the highest in immature fruits, gradually decreasing during fruit maturation, during which the radical scavenging activity also decreased. Based on our quantitative results, it was considered that the decrease in the flavan-3-ol derivative content due to fruit ripening was largely related to the increase in the amount of anthocyanin derivatives. Considering that the decreased contents of these compounds were related to the expression levels of polyphenol biosynthetic enzymes, quantification was performed using the semi-quantitative polymerase chain reaction, but the only change observed was the increased expression of the enzyme that synthesizes anthocyanins during maturation. Therefore, it was suggested that it is necessary to inhibit anthocyanin synthesis to increase the contents of highly functional flavan-3-ol derivatives in the mature fruit.
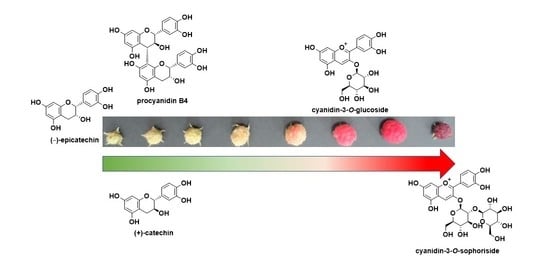
1. Introduction
There is currently great interest in the functional and bioactive compounds present in food sources since they are generally considered to be safe due to their frequent consumption in the daily diet. In particular, polyphenols, which are found in many healthy foods, as well as in fruits and vegetables, are thought to possess various health benefits [1,2].
Flavonoids, which is the generic term for flavones, anthocyanins, flavonols, flavan-3-ols (flavanols), and chalcones, are thought to be beneficial to human health because of their various biological activities. However, biological activity evaluations and elucidation of the mechanisms of action require large quantities of the corresponding compounds to obtain structure-activity relationship (SAR) profiles, and currently, it is difficult to obtain such quantities of polyphenol compounds from plants. To address this issue, we previously developed a stereoselective route to flavan-3-ol derivatives, oligomeric compounds [3,4,5,6,7,8], galloyled derivatives [9,10,11,12,13], and acylated derivatives [14,15,16], to permit SAR studies to be carried out, including those related to the anti-oxidative activities [5,6,7,8,9], radical scavenging activities [10,11], HeLa S3 proliferation-inhibitory activities [7,12,13,16], and anti-viral activities [17] of these compounds. It was found that fine structural differences, such as differences in stereochemistry and the presence or absence of modifications, are important in determining biological activity. In addition, these synthetic studies allowed us to prepare and store various compounds that are not commercially available to permit their use in further analytical and SAR studies. Indeed, our investigations demonstrated that flavan-3-ol derivatives possess a high functionality and that their oligomers exhibit various different biological activities. Therefore, our next challenge was to selectively and efficiently prepare the highly functional polyphenolic compounds present in plants.
For the purpose of the current study, we selected red raspberry (Rubus idaeus L.) as our experimental model plant because it is widely known for its edible fruits, in addition to the fact that it is rich in polyphenols and considered highly beneficial for human health [18,19]. Furthermore, red raspberries are common food products, and their extracts are known to exhibit anti-inflammatory [20], anti-metabolic syndrome [21], and antimicrobial properties [22]. Furthermore, it has been reported that increased raspberry consumption may improve age-related declines in balance and muscle strength [23]. There are also reports on the functionalities of compounds isolated from raspberries, such as the cytotoxicity of ellagitannin toward cancer cells [24] and the neuroprotective effect of phenylpropanoid compounds against cellular anti-oxygen stress [25].
The amount of polyphenol compounds present in raspberries varies depending on the cultivation environment [26,27] and the part of the plant being examined (e.g., leaves, stems, or fruits) [28]. Indeed, we previously reported that the composition of polyphenol compounds in the leaves of raspberries cultivated in a plant factory differed greatly depending on the cultivation environment, such as the irradiation wavelength of the light-emitting diode [29].
Recently, attention has been paid to changes in the contents of bioactive compounds during the maturation of various plants, and many studies have reported that the ripening stage is the point at which the majority of bioactive compounds are produced. For example, it has been confirmed that the anthocyanin content increases in the fruits of plums [30,31] and persimmons [32] during their maturation. Furthermore, in kiwifruits, it has been reported that the amounts of (+)-catechin (1) and (−)-epicatechin (2), which are also present in raspberries, increase during maturation [33]. Moreover, studies using grapes [34], redcurrants [35], cranberries [36], and strawberries [37] have reported changes in the amounts of such compounds present during maturation, wherein the expression levels of biosynthetic genes can vary. Recently, changes in the amounts of various bioactive compounds have been found during the maturation of Chinese raspberries [38]. More specifically, the color changes taking place and the amounts of carotenoids and polyphenol pigments were examined, and it was found that the polyphenol content tended to decrease with color change and maturation [38].
Thus, in the current study, the fruits of the Himbo-top® raspberry are classified into eight maturation stages, and the polyphenol content and DPPH and ABTS radical scavenging activities are examined at each stage. In addition, liquid chromatography-mass spectrometry is performed using our previously prepared standard compounds, and the flavan-3-ol monomer/oligomer, ellagitannin, and anthocyanin compounds present at each stage are quantified. Following this quantification, changes in the contents of these compounds during maturation are examined. Finally, the expression levels of flavan-3-ol biosynthesis-related enzymes are investigated, and the correlation between the total amount of anthocyanins and the anthocyanin biosynthetic enzymes is confirmed.
2. Materials and Methods
2.1. Reagents and Materials
HPLC grade methanol was used for extraction of the polyphenols, while molecular biology grade dimethyl sulfoxide (DMSO) was used to dissolve the crude extract. These solvents were obtained from FUJI FILM Wako Pure Chemical Co., Ltd. (Osaka, Japan). LC-MS grade acetonitrile and HPLC grade formic acid were obtained from Honeywell International Inc. (Trenton, NJ, USA). All commercially available chemicals employed for preparation of the crude extract solution were dissolved in DMSO and stored at −40 °C until required for use. Standard procyanidin B2 was obtained from Extrasynthese (Genay, France). The Folin-Ciocalteu reagent was obtained from Merck KGaA (Darmstadt, Germany). The DPPH (11-diphenyl-2-picrylhydrazyl) radical was obtained from Tokyo Chemical-Industry, Co., Ltd. (Tokyo, Japan). All other reagents and chemicals were special grade reagents, unless otherwise stated.
2.2. Raspberry Cultivation Method and Sample Preparation for Analysis
Himbo-Top® raspberry seedlings (Registration name of The Plant Variety Protection and Seed Act: RAFZAQU, Himbo Top®) used for the experiment were obtained from Tenkoen Co. (Yamagata, Japan). The raspberries were cultivated by open-field cultivation in Neyagawa City, Osaka Prefecture (latitude 34°45′37′′, longitude 135°37′37′′). The raspberry fruits were graded by their number of days of maturity. A sample (~10 g) of the raspberry fruit was crushed on ice with HPLC grade methanol (15 mL) in a mortar. Filtration was performed using Celite as a filter aid. The filtrate was then concentrated by evaporation under reduced pressure (RE601 Vacuum Controller, Tokyo, Japan) and dried in vacuo (GVD-05A, ULVAQ SINKU KIKO Co., Ltd., Miyazaki, Japan). The resulting residue was used to prepare a 100 mg/mL solution in DMSO. For HPLC analysis of the anthocyanins, samples of the fruits at each maturation stage were lyophilized for 24 h [39]. The dried fruits were powdered using a Speed mill MS-05 (Labonect, Osaka, Japan). To suppress heat, crushing was carried out three times for 20 s at 10 s intervals (60 s in total). The fruit samples (100 mg) were then extracted with 2% formic acid in methanol (1 mL) under vortex for 5 min, sonication for 5 min, and centrifugation for 10 min at 10,000× g. The pellets were re-extracted, and the supernatants were pooled before being reduced to dryness in vacuo. The obtained solid was used to prepare a 100 mg/mL solution in DMSO. The DMSO-solution extracts were stored at −40 °C until required for assaying and analysis.
2.3. Fruit Weight and Water Content
The ripening raspberry fruits were analyzed using a moisture meter MOC 63u (Shimadzu Corporation, Kyoto, Japan).
2.4. Total Polyphenol Analysis
The total phenolic content of each sample was determined using the modified Folin–Ciocalteu method [40,41]. More specifically, an aliquot (2 μL) of each crude extract (100 mg/mL in DMSO) was dissolved in Milli-Q water (200 μL) and mixed thoroughly with the Folin–Ciocalteu reagent (25 μL) and a 10% (w/v) sodium carbonate solution (25 μL). The mixture was allowed to stand for 30 min in the dark, and the absorbance was measured at 740 nm by a microplate reader (Multiskan FC, Thermo Fisher Scientific, CA, USA). The total phenolic content was calculated from a calibration curve, and the results are expressed as mg of gallic acid equivalents per 100 mg/mL DMSO solution of raspberry leaf extract.
2.5. Total Proanthocyanidin Analysis
The proanthocyanidin concentration of each sample was determined using the modified protein-precipitation method described by Harbertson et al. [42] and Cáceres-Mella et al. [43]. The protein solution for proanthocyanidin precipitation was prepared by dissolving BSA in a buffer containing 200 mM acetic acid and 170 mM NaCl adjusted to pH 4.9 using 1.0 M NaOH, to give a final protein concentration of 1.0 mg/mL. An aliquot (5 μL) of each raspberry leaf extract in DMSO was diluted with a buffer (125 μL) of 12% ethanol (v/v) containing 5 mg/mL potassium bitartrate and adjusted to pH 3.3 using 1.0 M HCl. An aliquot (250 μL) of the protein solution was then added to the diluted extract solution and incubated for 15 min at 4 °C. After centrifugation for 5 min at 15,000 rpm to pellet the proanthocyanidin protein precipitate, the pellet was washed with a buffer (50 μL) containing 200 mM acetic acid and 170 mM NaCl (pH 4.9). The wash solution was poured off, and an aliquot (70 μL) of a buffer containing 5% TAE (v/v) and 10% SDS (w/v) was added and then vortexed to dissolve the pellet completely. An aliquot (10 μL) of a ferric chloride reagent (10 mM FeCl3 in 0.01 N HCl) was immediately added to the mixture and incubated at room temperature (25 °C) for 10 min. After this time, the absorbance was read at 520 nm using a microplate reader (Multiskan FC, Thermo Fisher Scientific, San Jose, CA, USA). Total proanthocyanidin content was calculated from a calibration curve, and the results are expressed as mg of procyanidin B2 equivalent per 100 mg/mL DMSO solution of raspberry leaf extract.
2.6. Total Anthocyanin Analysis
Quantitative analysis of the total anthocyanins in the raspberry fruits was carried out using the method of Gao et al. [44]. The fruit at each ripening stage was extracted by adding methanol containing 5% formic acid (15 mL, HPLC grade). The total amount of anthocyanin was calculated from the values obtained by measuring the absorbance at 520 nm using a microplate reader. The results are expressed as mg of anthocyanin chloride.
2.7. DPPH Radical Scavenging Assay
The radical scavenging activity of each raspberry extract was determined by the DPPH assay as described earlier, with some modifications [45]. More specifically, a solution of DPPH in ethanol (30 μM, 1.0 mL) was added to an aliquot (1 μL) of each synthesized compound in DMSO and incubated at 30 °C for 30 min (n = 8). The scavenging activity was estimated from the OD (optical density) readings obtained at 515 nm using a microplate reader (Multiskan FC, Thermo Fisher Scientific, San Jose, CA, USA). Samples in which an aliquot (1 μL) of DMSO was added to ethanol (1.0 mL) were prepared as negative controls at the same time. The absorbance readings were converted into radical scavenging activities (as percentages) as follows: [(absorbance of the control − absorbance of the sample)/absorbance of the control] × 100; vitamin E (VE) was used as a standard.
2.8. ABTS Radical Scavenging Assay
The radical scavenging activity of each raspberry extract was determined by the ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) assay as described earlier, with some modifications [46]. More specifically, ABTS was dissolved in water to give a 7 mM concentration. ABTS radical cations were produced by reacting the ABTS stock solution with 2.45 mM potassium persulfate (final concentration) and allowing the mixture to stand in the dark at room temperature for 12–16 h before carrying out the assay. The ABTS radical stock solution was diluted with ethanol to an absorbance of 0.70 ± 0.02 at 740 nm. After adding an aliquot (1 mL) of the diluted ABTS radical solution to the raspberry leaf extract (1 μL) and mixing by vortexing, the reaction mixtures were incubated at 30 °C for 4 min. The scavenging activities (as percentages) were calculated from the absorbance readings at 740 nm as follows: [(absorbance of the control − absorbance of the sample)/absorbance of the control] × 100; (+)-Catechin was used as a standard.
2.9. Total Vitamin C Analysis
The amount of vitamin C was measured according to the method of Kishida et al. [47]. The mixture solution of raspberry fruit extract solution (1 μL, 10 mg/mL DMSO) and 5% metaphosphoric acid solution (1 mL) was prepared by the dropwise addition of an indophenol solution (0.15 mol/L) to oxidize vitamin C. Then, 2% thiourea-metaphosphoric acid solution (1 mL) was added to stop the oxidation reaction. Two percent (w/v) 2,4-dinitrophenylhydrazine-4.5 M sulfuric acid solution was added to the above reaction solution (250 μL) and heated at 50 °C for 90 min for derivatization. Quantitative analysis of vitamin C was performed by HPLC.
2.10. Total Glucose Analysis
The total glucose content was quantified according to the phenol-sulfuric acid method described by Dubois et al. [48]. More specifically, an aliquot (1 μL) of each crude extract (100 mg/mL in DMSO) was dissolved in a 5% phenol solution (200 μL) and mixed thoroughly with sulfuric acid (1 mL). The resulting mixture was then allowed to stand for 10 min, and its absorbance was measured at 740 nm.
2.11. LC-MS Analysis Conditions
LC-MS was performed using a Shimadzu LCMS-2020 system equipped with a DGU-20A3R degas unit, an LC-20A binary pump, an SIL-20AC autosampler, an SPD-M20A diode array detector, a CTO-20AC column oven, and a CBM-20A communications bus module connected to an LC work station (Shimadzu Corporation, Kyoto, Japan). A Wakopac® MS-5C18GT column (φ 150 mm × 2.0 mm, 5 μm, FUJIFILM Wako Pure Chemical Corporation, Osaka, Japan) was also employed. Mobile phase A consisted of 0.05% (v/v) formic acid, while mobile phase B was acetonitrile. Analyses of the raspberry extracts were carried out using a linear gradient from 0–35% B over 0–90 min; 35–100% B over 90–95 min; 100% B, over 95–100 min; and 0% B, over 100–110 min. The flow rate was set at 0.2 mL/min. The injection volume was 10 μL, and the temperature of the column oven was maintained at 40 °C. Anthocyanin analysis was carried out using 1% (v/v) formic acid as mobile phase A and acetonitrile as mobile phase B. Analyses of the raspberry extracts were achieved using a linear gradient from 5–25% B with a flow rate of 0.2 mL/min. A Shimadzu 2020 Quadrupole mass spectrometer (Shimadzu Corporation, Kyoto, Japan) equipped with a positive/negative ESI source was used as a detector. The mass spectrometer was operated in the negative selected-ion-monitoring (SIM) mode with a capillary voltage of 1.2 V for the identification of phenolic compounds, while the positive SIM mode was employed for the detection of betanin. The conditions for MS analysis were as follows: spray voltage = −3.5 V, dissolving line temperature = 250 °C, nebulizer gas flow = 1.5 L/min, heat block temperature = 200 °C, drying gas flow = 12.00 and 15.00 mL/min for the phenolics and betanin, respectively, and detector voltage = 1.2 V.
Identification of the flavan-3-ol derivatives was carried out by comparison with the retention times and mass spectra of standard samples. Commercially available catechins (Sigma-Aldrich Inc., St. Louis, MO, USA) and epicatechin (Sigma-Aldrich Inc., MO, USA) were used for identification. Procyanidin B4 was synthesized by our group [4,11]. The flavan-3-ol trimer was determined to be a trimer based on the similarity of its MS spectrum with those of procyanidins C1, C2, and C4, and other trimers [5]. However, the structure was not consistent with the procyanidin trimer synthesized and confirmed by our group. The quantitation of each compound amount was carried out from individual calibration curves.
2.12. Semi-Quantitative RT-PCR Analysis of Flavan-3-ol Derivative Biosynthetic-Enzyme-Related Gene Expression
The total RNA was extracted from raspberry leaves using an RNeasy Plant Mini Kit (Qiagen, Valencia, CA, USA) for the semi-quantitative reverse-transcription polymerase chain reaction (RT-PCR). The genes analyzed in this study were selected from those commonly used in raspberry analytical studies [49,50,51]. After treatment of the RNA samples with DNase using a DNAfree kit (Ambion; Applied Biosystems Japan, Tokyo, Japan), cDNAs were synthesized using SuperScript III reverse transcriptase (Invitrogen, Carlsbad, CA, USA), followed by incubation with RNase H (Invitrogen). RT-PCR was performed using GoTaq Green Master Mix (Promega, Madison, WI, USA) in a T100TM Thermal Cycler (Bio-Rad Laboratories, Hercules, CA, USA) with the gene-specific primer sets shown in Table 1 [6,34]. The reaction conditions were as follows: CHS [49]: after heating at 95 °C for 3 min, PCR was performed for 40 cycles (95 °C, 30 s; 50 °C, 30 s; 72 °C, 22 s); F3′H [50]: after heating at 95 °C for 5 min, PCR was performed for 40 cycles (95 °C, 30 s; 50 °C, 30 s; 72 °C, 45 s); DFR [50]: after heating at 95 °C for 5 min, PCR was performed for 40 cycles (95 °C, 30 s; 62 °C, 30 s; 72 °C, 45 s); FLS [50]: after heating at 95 °C for 5 min, PCR was performed for 40 cycles (95 °C, 30 s; 67 °C, 30 s; 72 °C, 45 s); UDP-glucose: after heating at 95 °C for 5 min, PCR was performed for 30 cycles (95 °C, 30 s; 47 °C, 30 s; 72 °C, 20 s); ANR [51]: after heating at 95 °C for 5 min, PCR was performed for 30 cycles (95 °C, 30 s; 49 °C, 30 s; 72 °C, 25 s); ANS [51]: after heating at 95 °C for 2 min, PCR was performed for 30 cycles (95 °C, 1 min; 55 °C, 20 s; 72 °C, 10 s); Histone: after heating at 95 °C for 3 min, PCR was performed for 29 cycles (95 °C, 30 s; 55 °C, 30 s; 72 °C, 25 s).
3. Results and Discussion
3.1. Raspberry Fruit Cultivation and Harvest Based on the Level of Maturity
Raspberries (Himbo Top®) were cultivated by open-field cultivation in Neyagawa City, Osaka Prefecture (latitude 34°45′37′′, longitude 135°37′37′′). The fruits were classified and analyzed according to the number of days elapsed from the day when the flower buds bloomed. As shown in Figure 1, it was confirmed that a red pigment, an anthocyanin, was produced as the fruit matured. Table 2 shows the fruit color, diameter, weight, and water content of the harvested fruit at each ripening stage. Generally, the fruit is considered suitable for consumption at maturity stage S7.
3.2. Analysis of the Polyphenol Content at Each Maturation Stage
The structures of the main polyphenolic compounds present in the raspberries are shown in Figure 2. Compounds 1–4 are flavan-3-ol derivatives. Compounds 5 and 6 are quercetin derivatives, with 6 being a glucuronic acid conjugate. Compounds 7 and 8 are ellagitannin derivatives, where 7 is a dimer, and 8 is a trimer. Compounds 9–12 are anthocyanin derivatives. Although many other polyphenol compounds are also present, we focused on these 12 compounds due to their relatively large abundances in raspberries. Thus, at each stage of ripening, the fruits were crushed in methanol using a mortar. After a thorough extraction with methanol, the solvent was evaporated to dryness, and the resulting residue was dissolved in DMSO to obtain a 100 mg/mL solution.
Figure 3 shows the total polyphenol contents (A), the total proanthocyanidin contents (B), and the total anthocyanin contents (C) of the extracted solutions obtained at the various ripening stages (S1–S8). As indicated, the total polyphenol content and the total proanthocyanidin content were highest in the S1 stage, although both the total amount of polyphenols and the total amount of proanthocyanidins decreased between stages S1 and S2. In contrast, the total amount of anthocyanins began to increase beyond stage S6 and reached their highest value during stage S8; this change in the total anthocyanin content correlated with the observed change in color (Figure 1).
3.3. Radical Scavenging Activity at Each Maturation Stage
As mentioned above, raspberries have been reported to exhibit antioxidant properties, which have been attributed to the presence of various polyphenolic compounds in addition to vitamins such as vitamin C. Thus, the DPPH and ABTS radical scavenging activities, which are indexes of the antioxidant activity, were measured at each stage of maturity to confirm whether the polyphenol content (Figure 3) correlated with the antioxidant activity. As can be seen from Figure 4A,B, stage S1 showed the highest values for both the DPPH and ABTS radical scavenging activities, and these activities decreased as the fruit maturation process advanced. This reduction correlated with the total polyphenol content and the total proanthocyanidin content (Figure 3B), but not with the total anthocyanin content, thereby indicating that the anti-oxidative activity of raspberries was not significantly affected by the anthocyanin content, but was instead due to other polyphenol compounds. In addition, considering that the vitamin C and sugar contained in raspberry fruits may affect the DPPH radical scavenging activity, we measured the amount of vitamin C during each ripening stage (Figure 4B) in addition to the amount of glucose (Figure 4C). These compounds were present in their highest quantities in the fruits of stage S1, but the change in their levels with maturity was considered to have a low correlation with the change in the DPPH radical scavenging activity.
3.4. Identification and Quantification of the Polyphenol Compounds by HPLC Analysis
From the results presented in Figure 3 and Figure 4, it was suggested that during stage S1, the raspberry fruit contained the highest polyphenol content and exhibited the highest DPPH radical scavenging activity. Therefore, to confirm which components were responsible for these observations, HPLC analysis was performed. Identification of the compounds was carried out by estimating the molecular weight through a combination of LCMS and comparison with our standards, and the obtained results agreed well with those of previous studies [29,52,53]. Figure 5 shows the HPLC chromatograms obtained for the raspberry fruit of stage S1. The chromatogram presented in Figure 5A was obtained under conditions where non-cationized compounds, such as flavan-3-ols, are well separated, while that of Figure 5B was obtained under conditions where cationized anthocyanin derivatives can be separated. Thus, Table 3 shows the quantitative results of these analyses, wherein (+)-catechin (1), (−)-epicatechin (2), procyanidin B4 (3), flavan-3-ol trimer (4), quercetin (5), quercetin-3-O-glucuronide (6), lambertianin C (7), sanguiin H-6 (8), cyanidin-3-O-sophoroside (9), cyanidin-3-O-sambubiside (10), cyanidin-3-O-glucoside (11), and cyanidin-3-O-rutinoside (12) were identified and quantified using calibration curves based on standard samples. As indicated, during maturation, the contents of (+)-catechin (1), (−)-epicatechin (2), and flavan-3-ol trimer (4) gradually decreased to 87–89% of their original content from S1 to S8. In contrast, the contents of procyanidin B4 (3) and sanguiin H-6 (8) decreased significantly during maturation, arriving at levels 30 and 49% of their original values by stage S8. Moreover, anthocyanin derivatives, namely cyanidin-3-O-sophoroside (9), cyanidin-3-O-sambubiside (10), cyanidin-3-O-glucoside (11), and cyanidin-3-O-rutinoside (12), were detected during the middle stages of maturity (i.e., S3–S6), and their contents increased during fruit ripening. Of these four anthocyanin derivatives, compound 9 was present in the highest quantities following maturation. Based on the above quantitative results, it was therefore considered that the reduced content of the flavan-3-ol derivatives during maturation was largely related to the increased amount of anthocyanin derivatives.
3.5. Semi-Quantitative RT-PCR Analysis of the Expression Levels of the Flavan-3-ol Biosynthetic Enzymes
As described above, the total amount of polyphenols and total proanthocyanidins decreased as the fruit matured, and the radical scavenging activity decreased accordingly. In contrast, the amount of anthocyanins increased with fruit ripening, but no correlation was found between this increase and the radical scavenging activity. As shown in Table 3, the quantities of the flavan-3-ol derivatives, and in particular that of the procyanidin B4 dimer (3), showed a considerable decrease. Since our previous studies suggested that flavan-3-ol derivatives exhibit beneficial functionalities, we believe that a further reduction in their content is undesirable. Therefore, we considered that the decrease in the amount of flavan-3-ol derivatives might be related to the expression of biosynthetic enzymes in the fruits.
The expression levels of the flavan-3-ol biosynthetic enzymes were then measured to determine if they were responsible for the increased anthocyanin levels and decreased flavan-3-ol derivative levels. Figure 6 shows the putative biosynthetic pathway of the flavan-3-ol derivatives in raspberries [53], wherein they are believed to be biosynthesized from p-coumaroyl-CoA via dihydroquercetin. It is considered that procyanidin B4 (3) and the flavan-3-ol trimer (4) are produced by the reaction of leucocyanidin (as an electrophile derived from dihydroquercetin) with nucleophiles such as catechin (1) and epicatechin (4). In addition, since leucocyanidin is expected to biosynthesize cyanidin, it is considered that the biosynthesis of flavan-3-ol and the biosynthesis of cyanidin are mutually influenced. Therefore, the expression levels of flavan-3-ol derivative biosynthetic enzymes were measured according to the maturity of the fruit (Figure 7).
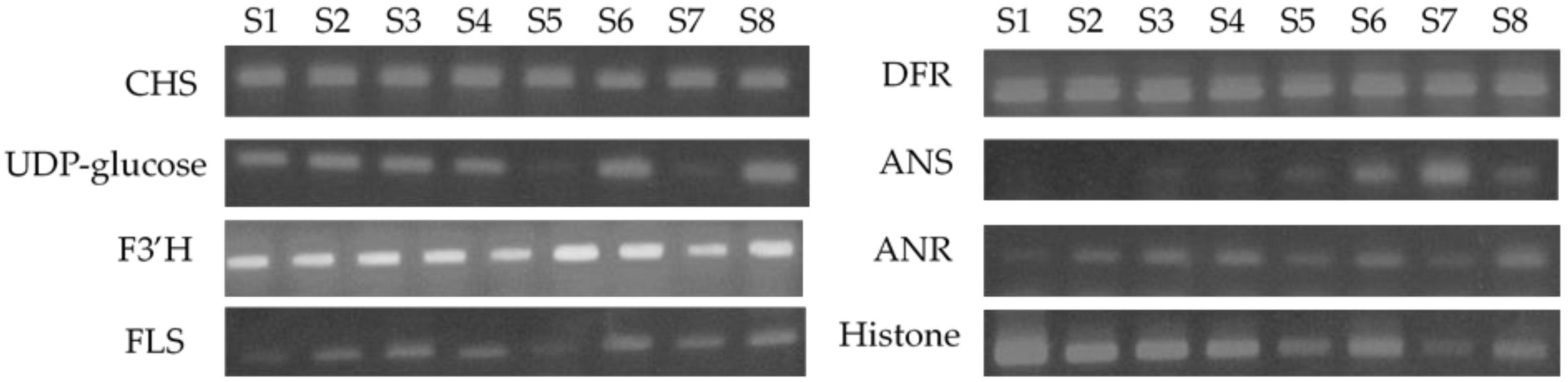
Figure 7 shows the semi-quantitative RT-PCR analysis of the expression levels of the flavan-3-ol derivative biosynthetic genes, namely, CHS, UDP-glucose, F3′H, FLS, DFR, ANS, and ANR (definitions provided in the caption of Figure 7). Histone was used as a control, and decreased expression levels of histone were found at fruit maturation stages S5 and S7. It should be noted that the reason for this phenomenon has not yet been confirmed. Among the various expression levels quantified, the one that changed most significantly was that of ANS, which synthesizes leucocyanidin from leucocyanidin. More specifically, the level of ANS increases significantly from stages S5 to S7, and this result correlates with the ripening stages in which the amount of anthocyanin derivatives increased. For the other enzymes examined, no significant differences were observed during maturation, and so our results suggest that the decrease in the amount of flavan-3-ol derivatives was caused by an increase in the expression of ANS, although the expression level of the external biosynthetic enzyme of the flavan-3-ol derivative did not change. Since the production of anthocyanins is promoted by this increase in the expression level of ANS, it was considered that the reduction in the flavan-3-ol content could not be prevented without lowering the levels of enzyme expression. Based on these results, we are currently investigating whether ANS expression can be reduced through variations in the cultivation conditions. However, we note that the color of the fruit indicates that the fruit is ripe and tasty, which increases the willingness of consumers to buy such products. In the future, we plan to study in detail the relationship between the expression of ANS and the biosynthesis of flavan-3-ol derivatives and search for conditions that do not reduce the amount of flavan-3-ol derivatives while expressing ANS.
4. Conclusions
In this study, the maturity of red raspberry fruits was classified into eight stages based on the polyphenol content at each stage. At each stage, the total polyphenol amount, total proanthocyanidin amount, and flavan-3-ol derivative amounts were determined, and the DPPH radical scavenging activity was evaluated. Based on our results, it was found that the content of flavan-3-ol derivatives gradually decreased during maturation, and this was accompanied by a reduction in the radical scavenging activity. Considering that the decreased levels of these compounds were related to the expression levels of polyphenol biosynthetic enzymes, quantification was performed using the semi-quantitative polymerase chain reaction, although the only change observed was an increase in the expression level of the anthocyanin biosynthetic enzyme during maturation. It was, therefore, suggested that it is necessary to inhibit anthocyanin synthesis to increase the content of highly functional flavan-3-ol derivatives in raspberry fruit.
Author Contributions
Conceptualization, R.K. and A.S.; chemical analysis, R.K.; raspberry cultivation, R.K. and S.Y.; RT-PCR measurements, R.K. and T.K.; supervision, T.K. and A.S.; project administration, A.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by BRAIN: Program for Promotion of Basic and Applied Research for Innovations in Bio-oriented Industry (GR104, A.S.), the Technology Research Promotion Program for Agriculture, Forestry, Fisheries, and Food Industry (A.S.), the Tojuro Iijima Foundation for Food and Technology (A.S.), and the Kurata Memorial Hitachi Science and Technology Foundation (A.S.)
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We heartedly acknowledge researchers at the Central Research Laboratory, Daiwa House Industry Co., Ltd., for their most helpful advice on raspberry cultivation within the plant factory unit. Similarly, we wish to express our sincere gratitude to the researchers at the Toyama Prefectural Agricultural, Forestry and Fisheries Research Center, who instructed us on the basic principles of raspberry cultivation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Harborne, J.B. The Flavonoids: Advances in Research from 1986; Chapman and Hall: London, UK, 1993. [Google Scholar]
- Harborne, J.B.; Baxter, H. The Handbook of Natural Flavonoids; Wiley: New York, NY, USA, 1999. [Google Scholar]
- Saito, A.; Nakajima, N.; Tanaka, A.; Ubukata, M. Synthetic studies of proanthocyanidins. Part 2. Stereoselective gram-scale synthesis of procyanidin-B3. Tetrahedron 2002, 58, 7829–7837. [Google Scholar] [CrossRef]
- Saito, A.; Nakajima, N.; Tanaka, A.; Ubukata, M. Synthetic studies of proanthocyanidins. Part 4. The synthesis of procyanidin B1 and B4. TMSOTf-catalyzed cyclization of catechin and epicatechin condensation. Heterocycles 2003, 61, 287–298. [Google Scholar]
- Saito, A.; Doi, Y.; Tanaka, A.; Matsuura, N.; Ubukata, M.; Nakajima, N. Systematic synthesis of four epicatechin series procyanidin trimers and their inhibitory activity on the Maillard reaction and antioxidant activity. Bioorg. Med. Chem. 2004, 12, 4783–4790. [Google Scholar] [CrossRef]
- Saito, A.; Mizushina, Y.; Tanaka, A.; Nakajima, N. Versatile synthesis of epicatechin series procyanidin oligomers, and their antioxidant and DNA polymerase inhibitory activity. Tetrahedron 2009, 65, 7422–7428. [Google Scholar] [CrossRef]
- Hamada, Y.; Takano, S.; Ayano, Y.; Tokunaga, M.; Koashi, T.; Okamoto, S.; Doi, S.; Ishida, M.; Kawasaki, T.; Hamada, M.; et al. Structure–Activity relationship of oligomeric flavan-3-ols: Importance of upper-unit B-ring hydroxyl groups in the dimeric structure for strong activities. Molecules 2015, 20, 18870–18885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashino, Y.; Okamoto, T.; Mori, K.; Kawasaki, T.; Hamada, M.; Nakajima, N.; Saito, A. Regioselective synthesis of procyanidin B6, a 4-6-condensed (+)-catechin dimer, by intramolecular condensation. Molecules 2018, 23, 205. [Google Scholar] [CrossRef] [Green Version]
- Saito, A.; Emoto, M.; Tanaka, A.; Doi, Y.; Shoji, K.; Mizushina, Y.; Ikawa, H.; Yoshida, H.; Matsuura, N.; Nakajima, N. Stereoselective synthesis of procyanidin B3-3-O-gallate and 3,3′′-di-O-gallate, and their abilities as antioxidant and DNA polymerase inhibitor. Tetrahedron 2004, 60, 12043–12049. [Google Scholar] [CrossRef]
- Saito, A.; Mizushina, Y.; Ikawa, H.; Yoshida, H.; Doi, Y.; Tanaka, A.; Nakajima, N. Systematic synthesis of galloyl-substituted procyanidin B1 and B2, and their ability of DPPH radical scavenging activity and inhibitory activity of DNA polymerases. Bioorg. Med. Chem. 2005, 13, 2759–2771. [Google Scholar] [CrossRef] [PubMed]
- Sakuda, H.; Saito, A.; Mizushina, Y.; Yoshida, H.; Tanaka, A.; Nakajima, N. Synthesis of galloyl-substituted procyanidin B4 series, and their deep radical scavenging activity and DNA polymerase inhibitory activity. Heterocycles 2006, 67, 175–188. [Google Scholar]
- Mori, K.; Ayano, Y.; Hamada, Y.; Hojima, T.; Tanaka, R.; Higashino, Y.; Izuno, M.; Okamoto, T.; Kawasaki, T.; Hamada, M.; et al. Role of 2,3-cis structure of (–)-epicatechin-3,5-O-digallate in inhibition of HeLa S3 cell proliferation. Nat. Prod. Chem. Res. 2015, 3, 172. [Google Scholar]
- Hojima, T.; Komeda, S.; Higashino, Y.; Hamada, M.; Nakajima, N.; Kawasaki, T.; Saito, A. Role of 3,5-digalloyl and 3′,4′-dihydroxyl structure of (–)-epicatechin-3,5-digallate in inhibition of HeLa S3 cell proliferation. Nat. Prod. Chem. Res. 2017, 5, 250. [Google Scholar] [CrossRef]
- Matsubara, K.; Saito, A.; Tanaka, A.; Nakajima, N.; Akagi, R.; Mori, M.; Mizushina, Y. Epicatechin conjugated with fatty acid is a potent inhibitor of DNA polymerase and angiogenesis. Life Sci. 2007, 80, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, S.; Doi, S.; Harui, K.; Okamoto, T.; Okamoto, S.; Uenishi, J.; Kawasaki, T.; Nakajima, N.; Saito, A. Development of a new synthetic strategy for procyanidin dimer condensation using peracetylated electrophiles. Heterocycles 2014, 88, 1595–1602. [Google Scholar]
- Okamoto, S.; Ishihara, S.; Okamoto, T.; Doi, S.; Harui, K.; Higashino, Y.; Kawasaki, T.; Nakajima, N.; Saito, A. Inhibitory activity of synthesized acetylated procyanidin B1 analogues against HeLa S3 cells proliferation. Molecules 2014, 19, 1775–1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukuda, S.; Watashi, K.; Hojima, T.; Isogawa, M.; Iwamoto, M.; Omagari, K.; Suzuki, R.; Aizaki, H.; Kojima, S.; Sugiyama, M.; et al. A new class of hepatitis B and D virus entry inhibitors, proanthocyanidin and its analogs, that directly act on the viral large surface proteins. Hepatology 2017, 65, 1104–1116. [Google Scholar] [CrossRef] [Green Version]
- Hummer, K.E. Rubus pharmacology: Antiquity to the present. HortScience 2010, 45, 1587–1591. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.V.; Synder, D.M. Raspberries and human health: A review. J. Agric. Food Chem. 2010, 58, 3871–3883. [Google Scholar] [CrossRef] [PubMed]
- Szymanowska, U.; Baraniak, B.; Bogucka-Kocka, A. Antioxidant, anti-inflammatory, and postulated cytotoxic activity of phenolic and anthocyanidin-rich fractions from polana raspberry (Rubus ideaus L.) fruit and juice-in vitro study. Molecules 2018, 23, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, T.; Miranda-Garcia, O.; Sasaki, G.; Shay, N.F. Consumption of a single serving of red raspberries per day reduces metabolic syndrome parameters in high-fat fed mice. Food Funct. 2017, 8, 4081–4088. [Google Scholar] [CrossRef] [PubMed]
- Olle, M.; Ngouajio, M.; Siomos, A. Vegetable quality and productivity as influenced by growing medium: A review. Zemdirb. Agric. 2012, 99, 399–408. [Google Scholar]
- Shukitt-Hale, B.; Thangthaeng, N.; Kelly, M.E.; Smith, D.E.; Miller, M.G. Raspberry differentially improves age-related declines in psychomotor function dependent on baseline motor ability. Food Funct. 2017, 8, 4752–4759. [Google Scholar] [CrossRef]
- Nowak, A.; Sojka, M.; Klewicka, E.; Lipinska, L.; Klewicki, R.; Kolodziejczyk, K. Ellagitannins from Rubus ideaus L. exert geno- and cytotoxic effects against human colon adenocarcinoma cell line Caco-2. J. Agric. Food Chem. 2017, 65, 2947–2955. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yao, G.D.; Song, X.Y.; Wang, B.; Lin, B.; Wang, X.B.; Huang, X.X.; Song, S.J. Neuroprotective effects of 1,2-diarylpropane type phenylpropanoid enantiomers from red raspberry against H2O2-induced oxidative stress in human neuroblastoma SH-SY5Y cells. J. Agric. Food Chem. 2018, 66, 331–338. [Google Scholar] [CrossRef]
- Wang, Y.; Liang, J.; Luan, G.; Zhang, S.; Zhuoma, Y.; Xie, J.; Zhou, W. Quantitative analysis of nine phenolic compounds and their antioxidant activities from thirty-seven varieties of raspberry grown in the Qinghai-Tibetan plateau region. Molecules 2019, 24, 3932. [Google Scholar] [CrossRef] [Green Version]
- Ponder, A.; Hallmann, E. The effects of organic and conventional farm management and harvest time on the polyphenol content in different raspberry cultivars. Food Chem. 2019, 301, 125295. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Liu, Y.; Qin, Y.; Wang, L.; Wu, Z. HPLC-ESI-qTOF-MS/MS characterization, antioxidant activities and inhibitory ability of digestive enzymes with molecular docking analysis of various parts of raspberry (Rubus ideaus L.). Antioxidants 2019, 8, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobori, R.; Hashimoto, S.; Koshimizu, H.; Yakami, S.; Hirai, M.; Noro, K.; Kawasaki, T.; Saito, A. Flavan-3-ols content in red raspberry leaves increases under blue LED-light irradiation. Metabolites 2019, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- LI, Q.; Chang, X.X.; Wang, H.; Brennan, C.S.; Guo, X.B. Phytochemicals accumulation in Sanhua Plum (Prunus salicina L.) during fruit development and their potential use as antioxidants. J. Agric. Food Chem. 2019, 67, 2459–2466. [Google Scholar] [CrossRef] [PubMed]
- Ndou, A.; Tinyani, P.P.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. An integrated approach for harvesting Natal plum (Carissa macrocarpa) for quality and functional compounds related to maturity stages. Food Chem. 2019, 293, 499–510. [Google Scholar] [CrossRef]
- Bordiga, M.; Travaglia, F.; Giuffrida, D.; Mangraviti, D.; Rigano, F.; Mondello, L.; Arlorio, M.; Coïsson, J.D. Characterization of peel and pulp proanthocyanidins and carotenoids during ripening in persimmon “Kaki Tipo” cv, cultivated in Italy. Food Res. Int. 2019, 120, 800–899. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Chen, D.; Fan, M.; Young, Q.S. UPLC-QqQ-MS/MS-based phenolic quantification and antioxidant activity assessment for thinned young kiwifruits. Food Chem. 2019, 281, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Bombai, G.; Pasini, F.; Verardo, V.; Sevindik, O.; Di Foggia, M.; Tessarin, P.; Bregoli, A.M.; Taddei, P.; Cabone, M.F.; Rombola, A.D. Monitoring of compositional changes during berry ripening in grape seed extracts of cv. sagiovese (Vitis vinifera L.). J. Sci. Food Agric. 2017, 97, 3058–3064. [Google Scholar] [CrossRef]
- Zorenc, Z.; Veberic, R.; Koron, D.; Miosic, S.; Hutabarat, O.S.; Halbwirth, H.; Mikulic-Petkovsek, M. Polyphenol metabolism in different colored cultivars of red currant (Ribes rubrum L.) through fruit ripening. Planta 2017, 246, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Johnson-Cicalese, J.; Singh, A.P.; Vorsa, N. Characterization and quantification of flavonoids and organic acid over fruit development in American cranberry (Vaccinium macrocarpon) cultivars using HPLC and APCI-MS/MS. Plant Sci. 2017, 262, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Nagpala, E.G.; Guidarelli, M.; Gasperotti, M.; Masuero, D.; Bertolini, P.; Vrhovsek, U.; Baraldi, E. Polyphenols variation in fruits of the susceptible strawberry cultivar alba during-ripening and upon fungal pathogen interaction and possible involvement in unripe fruit tolerance. J. Agric. Food Chem. 2016, 64, 1869–1878. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, J.; Chen, Z.; Jiang, J.; Jacson, A. Characterization of carotenoids and phenolics during fruit ripening of Chinese raspberry (Rubus chingii Hu). RSC Adv. 2021, 11, 10804–10813. [Google Scholar] [CrossRef]
- Boeges, G.; Degeneve, A.; Mullen, W.; Crozier, A. Identification of Flavonoid and Phenolic Antioxidants in Black Currants, Blueberries, Raspberry, Red Currants, and Cranberries. J. Agric. Food Chem. 2010, 58, 3901–3909. [Google Scholar]
- Folin, O.; Denis, W. A Colorimetric method for the determination of phenols (and derivatives) in urine. J. Biol. Chem. 1915, 22, 305–308. [Google Scholar] [CrossRef]
- Juilcunen-Tiittoo, R. Phenolic constituents in the leaves of northern willows: Methods for the analysis of certain phenolics. J. Agric. Food Chem. 1985, 33, 213–217. [Google Scholar] [CrossRef]
- Harbertrson, J.F.; Kennedy, J.A.; Adams, D.O. Tannin in skins and seed of cabernet sauvignon, syrah, and pinot noir berries during ripening. Am. J. Enol. Vitic. 2002, 52, 54–59. [Google Scholar]
- Cáceres-Mella, Á.; Peña-Neira, Á.; Narváez-Bastias, J.; Jara-Campos, C.; López-Solís, R.; Canals, J.M. Comparison of analysis methods for measuring proanthocyanidins in wines and their relationship with perceived astringency. Int. J. Food Sci. Technol. 2013, 48, 2588–2594. [Google Scholar] [CrossRef]
- Gao, L.; Mazz, G. Quantitation and distribution of simple and acylated anthocyanins and other phenolics in blueberries. J. Food Sci. 1994, 59, 1057–1059. [Google Scholar] [CrossRef]
- Nanjo, F.; Goto, K.; Seto, R.; Suzuki, M.; Sakai, M.; Hara, Y. Scavenging effects of tea catechins and their derivatives on 1,1-diphenyl-2-picrylhydrazyl radical. Free Radic. Biol. Med. 1996, 21, 895–902. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Rad. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Kishida, E.; Nishimoto, Y.; Kojo, S. Specific determination of ascorbic acid with chemical derivatization and high-performance liquid chromatography. Anal. Chem. 1992, 64, 1505–1507. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substance. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Zheng, D.; Schröder, G.; Schröder, J.; Hrazdina, G. Molecular and biochemical characterization of three aromatic polyketide synthase genes from Rubus idaeus. Plant Mol. Biol. 2001, 46, 1–15. [Google Scholar] [CrossRef] [PubMed]
- McCallum, S.; Woodhead, M.; Hackett, C.A.; Kassim, A.; Paterson, A.; Graham, J. Genetic and environmental, effects influencing fruit color and QTL analysis in raspberry. Theor. Appl. Genet. 2010, 121, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Rafique, M.Z.; Carvalho, E.; Stracke, R.; Palmieri, L.; Harrera, L.; Feller, A.; Malnoy, M.; Martens, S. Martens. Nonsense mutation inside anthocyanidin synthase gene controls pigmentation in yellow raspberry (Rubus idaeus L.). Front. Plant Sci. 2016, 7, 1892. [Google Scholar] [CrossRef] [PubMed]
- Beekwilder, J.; Jonker, H.; Meesters, P.; Hall, R.D.; Van der Meer, I.M.; Ric de Vos, C.H. Antioxidants in Raspberry: On line analysis links antioxidant activity to diversity of individual metabolites. J. Agric. Food Chem. 2005, 53, 3313–3320. [Google Scholar] [CrossRef] [PubMed]
- Krauze-Baranowska, M.; Glod, D.; Kula, M.; Majdam, M.; Halasa, R.; Matkowski, A.; Kozlowska, W.; Kawiak, A. Chemical composition and biological activity of Rubus ideaus shoots—A traditional herbal remedy of Eastern Europe. BMC Complement. Altern. Med. 2014, 14, 480. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Maturation stages S1–S8 of the raspberry fruit. The stages were classified according to the number of days elapsed from the day when the flower buds bloomed (day 1); S1: 6–9 d; S2: 10–14 d; S3: 15–18 d; S4: 19–23 d; S5: 24–28 d; S6: 29–30 d; S7: 31–36 d; and S8: 37–44 d.
Figure 1.
Maturation stages S1–S8 of the raspberry fruit. The stages were classified according to the number of days elapsed from the day when the flower buds bloomed (day 1); S1: 6–9 d; S2: 10–14 d; S3: 15–18 d; S4: 19–23 d; S5: 24–28 d; S6: 29–30 d; S7: 31–36 d; and S8: 37–44 d.
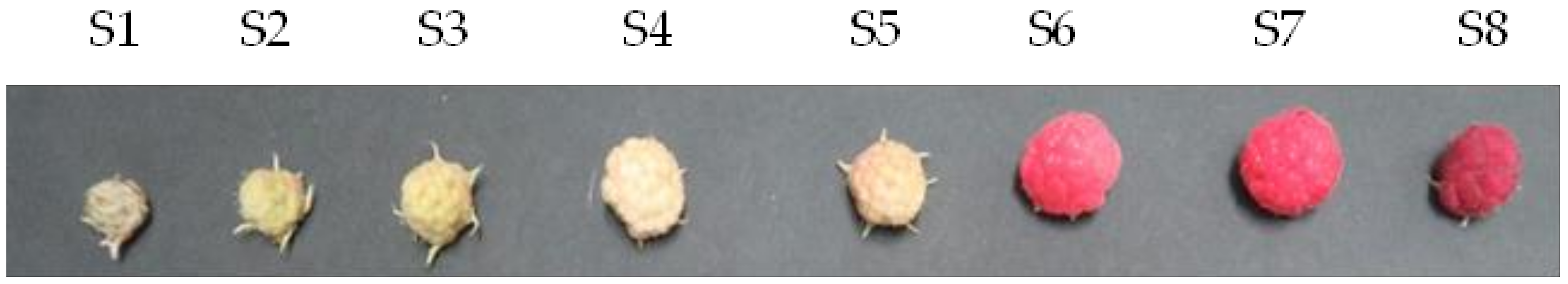
Figure 2.
Chemical structures of the raspberry fruit polyphenol compounds (1–12). 1: (+)-Catechin; 2 (−)-epicatechin; 3: procyanidin B4; 4: flavan-3-ol trimer; 5: quercetin; 6: quercetin-3-O-glucuronide; 7: lambertianin C; 8: sanguiin H-6; 9: cyanidin-3-O-sophoroside; 10: cyanidin-3-O-sambubiside; 11: cyanidin-3-O-glucoside; and 12: cyanidin-3-O-rutinoside.
Figure 2.
Chemical structures of the raspberry fruit polyphenol compounds (1–12). 1: (+)-Catechin; 2 (−)-epicatechin; 3: procyanidin B4; 4: flavan-3-ol trimer; 5: quercetin; 6: quercetin-3-O-glucuronide; 7: lambertianin C; 8: sanguiin H-6; 9: cyanidin-3-O-sophoroside; 10: cyanidin-3-O-sambubiside; 11: cyanidin-3-O-glucoside; and 12: cyanidin-3-O-rutinoside.
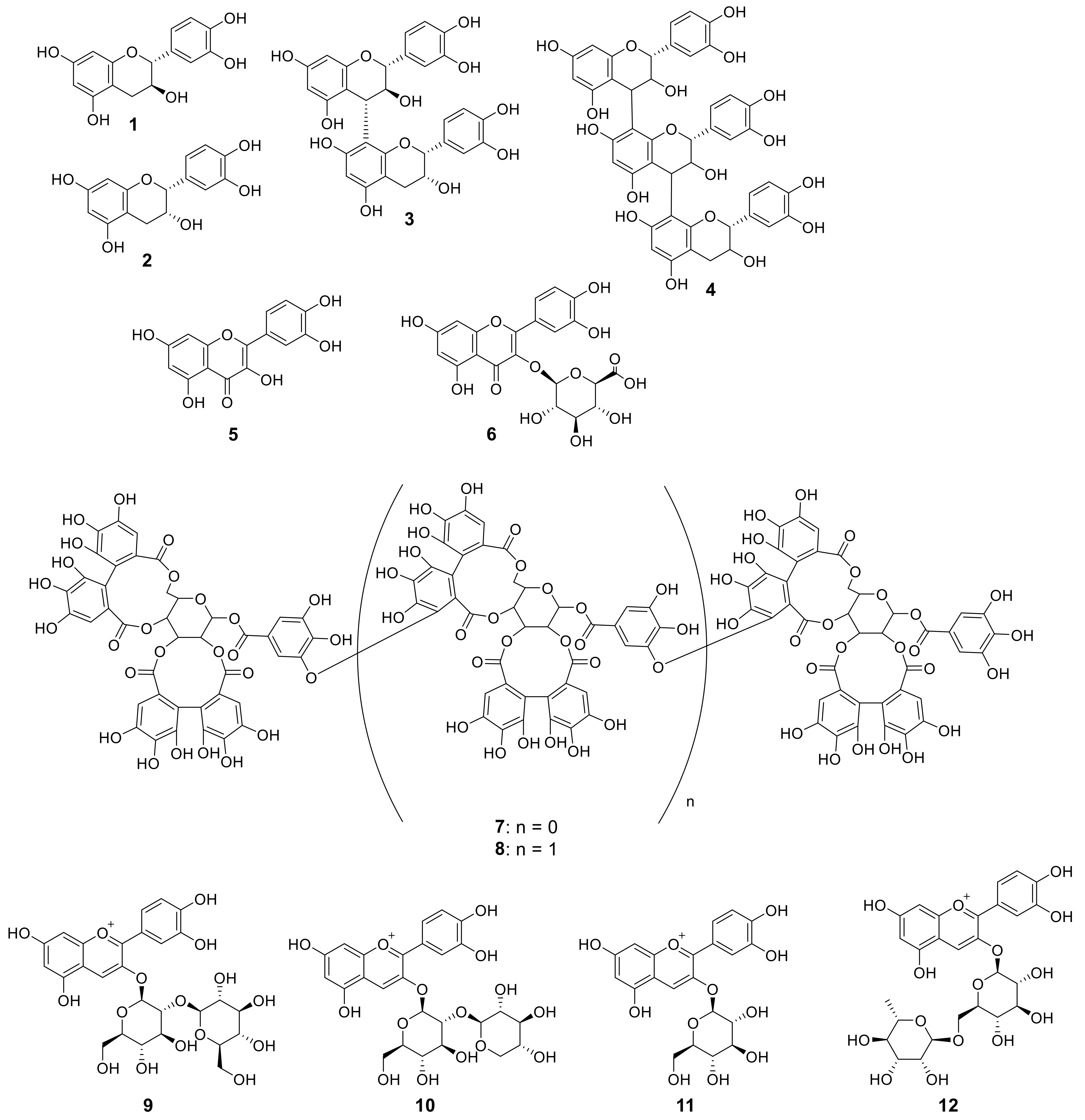
Figure 3.
Quantification of the amounts of polyphenol compounds during ripening stages S1–S8. (A) The total polyphenol content was determined by the Folin-Ciocalteu method. Data represent mg of gallic acid equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. (B) The total proanthocyanidin content was determined by a modified protein precipitation method. Data represent mg of procyanidin B2 equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. (C) The total anthocyanin content was measured using fruits extracted with methanol containing 5% formic acid. Data represent mg of cyanidin chloride equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. Error bars represent the standard deviation (SD) of the mean (n = 8).
Figure 3.
Quantification of the amounts of polyphenol compounds during ripening stages S1–S8. (A) The total polyphenol content was determined by the Folin-Ciocalteu method. Data represent mg of gallic acid equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. (B) The total proanthocyanidin content was determined by a modified protein precipitation method. Data represent mg of procyanidin B2 equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. (C) The total anthocyanin content was measured using fruits extracted with methanol containing 5% formic acid. Data represent mg of cyanidin chloride equivalents per 100 mg/mL DMSO solution of the raspberry fruit extract. Error bars represent the standard deviation (SD) of the mean (n = 8).
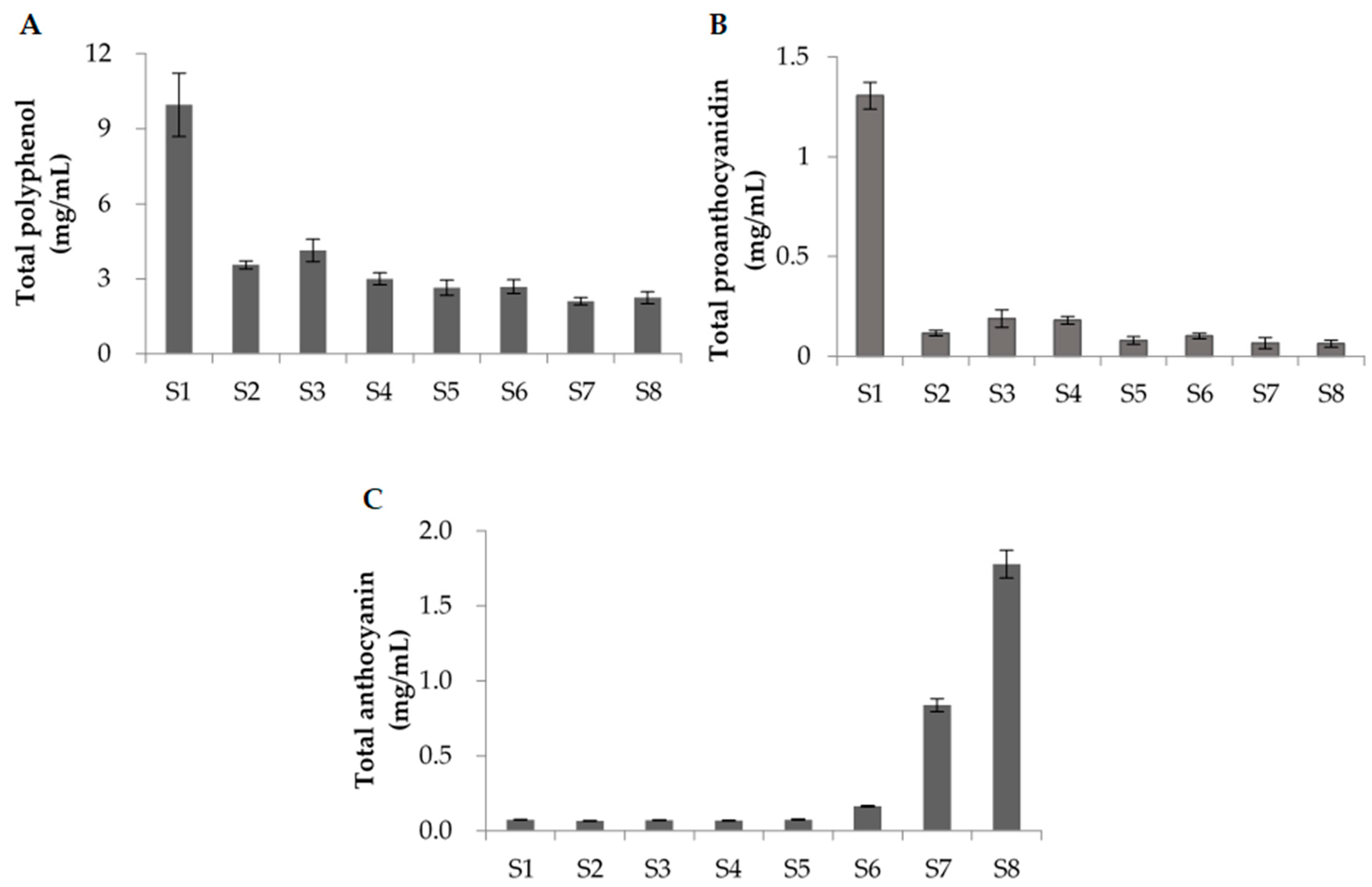
Figure 4.
DPPH radical scavenging activity and ABTS radical scavenging activity at each ripening stage. (A) DPPH radical scavenging activity. Vitamin E was used as a control. Error bars represent the SD of the mean (n = 8). (B) ABTS radical scavenging activity. (+)-Catechin was used as a control. Error bars represent the SD of the mean (n = 8). (C) The vitamin C content was quantified using the 2,4-dinitrophenylhydrazine method. Data represent 1 mg of ascorbic acid equivalents per 100 mg/mL of the raspberry fruit extract DMSO solution. Error bars represent the SD of the mean (n = 4). (D) The total glucose content was determined using the phenol-sulfuric acid method. Data represent 1 mg of glucose equivalents per 100 mg/mL of the raspberry fruit extract DMSO solution. Error bars represent the SD of the mean (n = 4).
Figure 4.
DPPH radical scavenging activity and ABTS radical scavenging activity at each ripening stage. (A) DPPH radical scavenging activity. Vitamin E was used as a control. Error bars represent the SD of the mean (n = 8). (B) ABTS radical scavenging activity. (+)-Catechin was used as a control. Error bars represent the SD of the mean (n = 8). (C) The vitamin C content was quantified using the 2,4-dinitrophenylhydrazine method. Data represent 1 mg of ascorbic acid equivalents per 100 mg/mL of the raspberry fruit extract DMSO solution. Error bars represent the SD of the mean (n = 4). (D) The total glucose content was determined using the phenol-sulfuric acid method. Data represent 1 mg of glucose equivalents per 100 mg/mL of the raspberry fruit extract DMSO solution. Error bars represent the SD of the mean (n = 4).
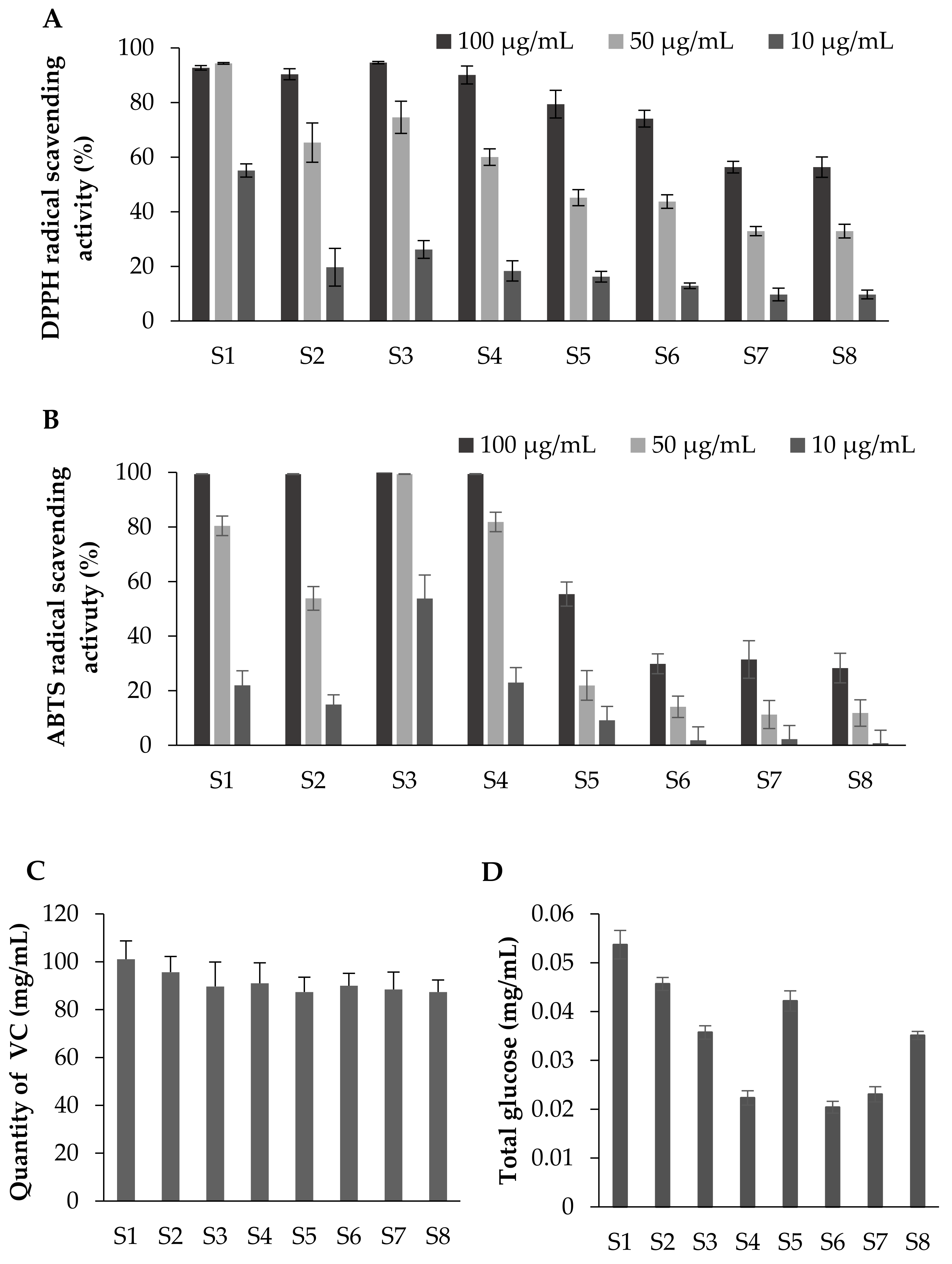
Figure 5.
HPLC chromatogram of the raspberry fruit extract of S1. Peak number 1: (+)-Catechin (RT: 19.66 min, MS: 289 m/z); 2 (−)-Epicatechin (RT: 23.41 min, MS: 289 m/z); 3: Procyanidin B4 (RT: 20.78 min, MS: 577 m/z); 4: Flavan-3-ol trimer (RT: 22.57 min, MS: 865 m/z); 5: Quercetin (RT: 31.44 min, MS: 301 m/z); 6: Quercetin-3-O-glucuronide (RT: 32.21 min, MS: 447 m/z); 7: Lambertianin C (RT: 27.79 min); 8: Sanguiin H-6 (RT: 28.63 min, MS: 934 m/z); 9: Cyanidin-3-O-sophoroside; 10: Cyanidin-3-O-sambubiside; 11: Cyanidin-3-O-glucoside; 12: Cyanidin-3-O-rutinoside. The structure of each compound is shown in Figure 1. (A) General conditions for polyphenol analysis. (B) Conditions for anthocyanin analysis.
Figure 5.
HPLC chromatogram of the raspberry fruit extract of S1. Peak number 1: (+)-Catechin (RT: 19.66 min, MS: 289 m/z); 2 (−)-Epicatechin (RT: 23.41 min, MS: 289 m/z); 3: Procyanidin B4 (RT: 20.78 min, MS: 577 m/z); 4: Flavan-3-ol trimer (RT: 22.57 min, MS: 865 m/z); 5: Quercetin (RT: 31.44 min, MS: 301 m/z); 6: Quercetin-3-O-glucuronide (RT: 32.21 min, MS: 447 m/z); 7: Lambertianin C (RT: 27.79 min); 8: Sanguiin H-6 (RT: 28.63 min, MS: 934 m/z); 9: Cyanidin-3-O-sophoroside; 10: Cyanidin-3-O-sambubiside; 11: Cyanidin-3-O-glucoside; 12: Cyanidin-3-O-rutinoside. The structure of each compound is shown in Figure 1. (A) General conditions for polyphenol analysis. (B) Conditions for anthocyanin analysis.
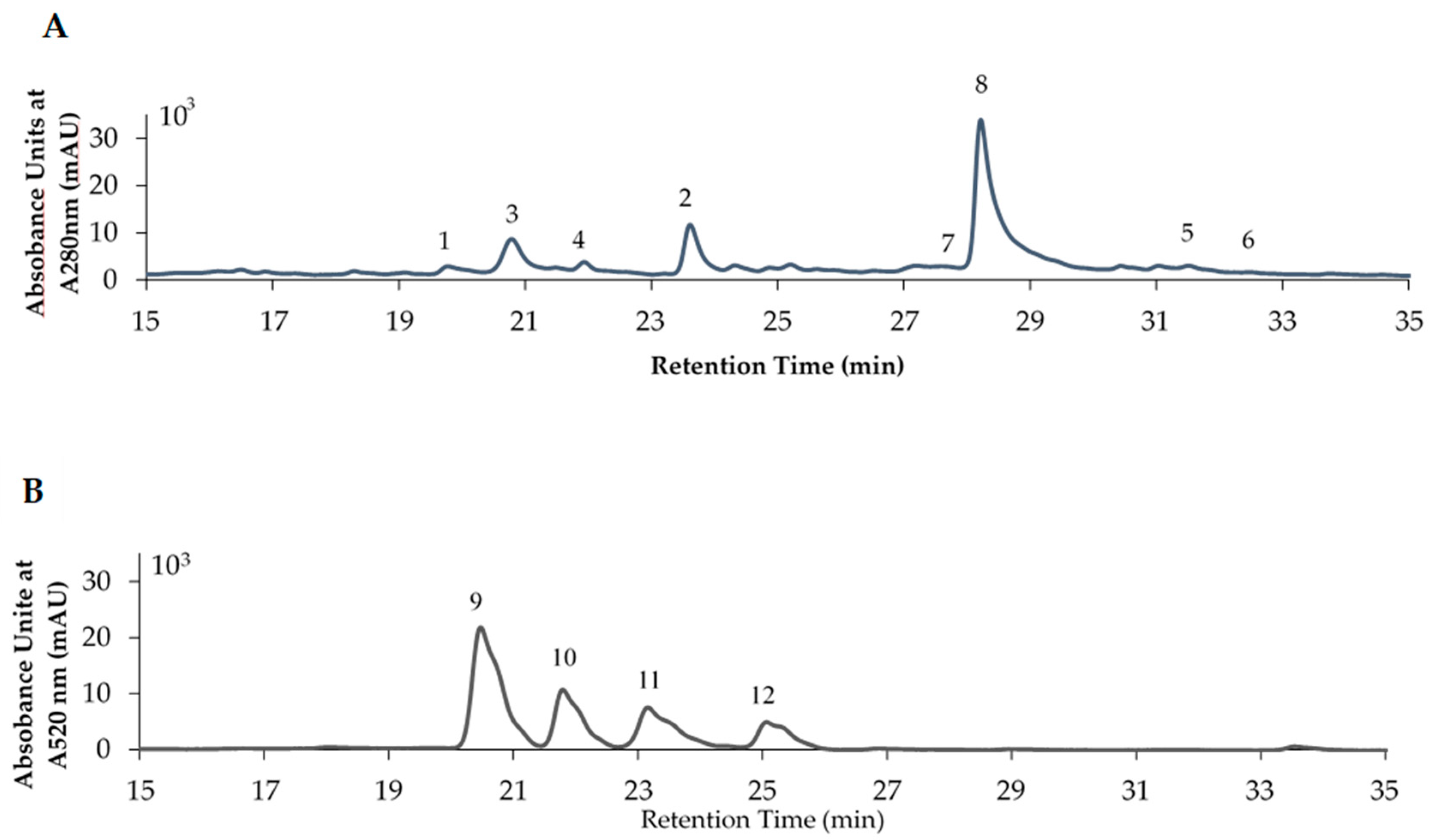
Figure 6.
Partial schematic representation of flavonoid biosynthesis. CHS: chalcone synthase; UDP-glucose: uridine diphosphate glucose; F3′H: flavonoid 3′-hydroxylase; FLS: flavanol synthase; DFR: dihydroflavanol 4-reductase; LAR: leucoanthocyanidin reductase; ANS: anthocyanin synthase; and ANR: anthocyanidin reductase.
Figure 6.
Partial schematic representation of flavonoid biosynthesis. CHS: chalcone synthase; UDP-glucose: uridine diphosphate glucose; F3′H: flavonoid 3′-hydroxylase; FLS: flavanol synthase; DFR: dihydroflavanol 4-reductase; LAR: leucoanthocyanidin reductase; ANS: anthocyanin synthase; and ANR: anthocyanidin reductase.
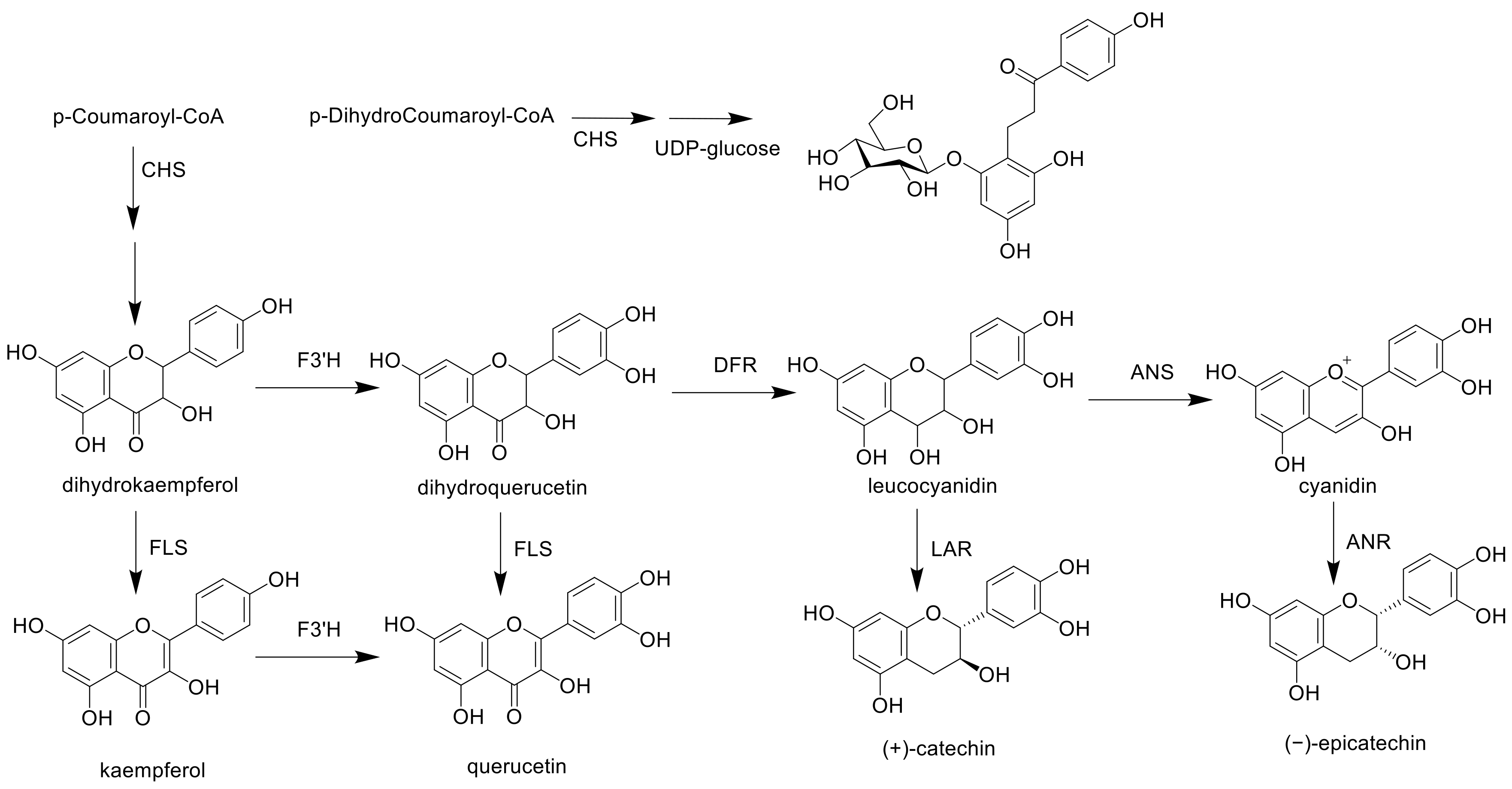
Figure 7.
Expression levels of genes encoding flavan-3-ol biosynthetic enzymes throughout the various maturation stages of raspberry fruit. The total RNA was extracted from fresh fruits and was used for sqRT-PCR analysis.
Figure 7.
Expression levels of genes encoding flavan-3-ol biosynthetic enzymes throughout the various maturation stages of raspberry fruit. The total RNA was extracted from fresh fruits and was used for sqRT-PCR analysis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The sequence of the primers used for the RT-PCR experiments.
| Gene | Accession No. of Sequence Used for Primer Design | Primer Sequence |
|---|---|---|
| CHS | AF292367 | F: AAC CCT TGT TTC TTC GTA CCA TTA |
| R: GAT GGG TAG CTA GTA CTT ACA CAT | ||
| UDP-glucose | AWT04749 | F: CCATGTTTTCTTGGTTTCCTT |
| R: ATGAAAGGGTTGTTAATGAGG | ||
| F3′H | GT029980 | F: TGA TGA AGC TTT ATA AGC ATG TGA GC |
| R: GGG TCC ACT CTC TTG GTG AA | ||
| FLS | GT029981 | F: AGG TGA ACAGGT GGA GTT GG |
| R: TGA AGA CCA TCA TCG AAT GC | ||
| DFR | GT029979 | F: ATG CGA AAC AAC TTG CAT TT |
| R: GCT ACG ATT CAC GAC ATT GC | ||
| ANS | KX950789.1 | F: ATC GTA ATG CAC ATA GGC GAC ACC |
| R: CCT TGG GCG GCT CAG AGA AAA | ||
| ANR | AMP19723 | F: ATC TCA AAC AAG ACT GCT TGT G |
| R: GAG AGT ATT GAC AGT CAC TGC AG | ||
| Histone | AF301365.1 | F: CAA GGA AGC AAT TGG CTA CCA AGG |
| R: AGT TGG ATA TCC TTG GGC ATA ATA |
Table 2.
Fruit diameter, weight, and water content at each ripening stage.
| S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | |
|---|---|---|---|---|---|---|---|---|
| Fruit Color | green | green | green | white | white | red | red | red |
| Fruit Size (cm) | 0.80 ± 0.05 | 1.05 ± 0.03 | 1.15 ± 0.05 | 1.37 ± 0.05 | 1.37 ± 0.03 | 1.48 ± 0.05 | 1.53 ± 0.06 | 1.19 ± 0.11 |
| Fruit Weight (g) | 0.19 ± 0.03 | 0.47 ± 0.01 | 0.68 ± 0.04 | 1.05 ± 0.16 | 1.11 ± 0.27 | 1.39 ± 0.03 | 1.45 ± 0.06 | 0.68 ± 0.22 |
| Water Content (%) | 70.6 ± 5.35 | 77.2 ± 3.00 | 80.8 ± 0.49 | 84.4 ± 1.53 | 83.7 ± 2.34 | 85.4 ± 0.30 | 85.2 ± 0.44 | 78.2 ± 3.70 |
Table 3.
Polyphenol contents (mg/100 mg of DW) in raspberry fruit extracts at various stages of maturation.
Table 3.
Polyphenol contents (mg/100 mg of DW) in raspberry fruit extracts at various stages of maturation.
| Mg/ML | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 |
|---|---|---|---|---|---|---|---|---|
| (+)-catechin (1) | 2.29 ± 0.005 | 2.29 ± 0.004 | 2.29 ± 0.006 | 2.18 ± 0.009 | 2.04 ± 0.0006 | 2.02 ± 0.005 | 2.00 ± 0.008 | 2.00 ± 0.003 |
| (−)-epicatechin (2) | 3.12 ± 0.01 | 3.33 ± 0.009 | 3.07 ± 0.03 | 2.70 ± 0.02 | 2.74 ± 0.02 | 2.70 ± 0.006 | 2.71 ± 0.007 | 2.70 ± 0.008 |
| procyanidin B4 (3) | 0.23 ± 0.002 | 0.22 ± 0.0001 | 0.21 ± 0.001 | 0.11 ± 0.002 | 0.09 ± 0.001 | 0.08 ± 0.001 | 0.07 ± 0.002 | 0.07 ± 0.001 |
| flavan-3-ol trimer (4) | 0.35 ± 0.0006 | 0.35 ± 0.0001 | 0.33 ± 0.0001 | 0.31 ± 0.0005 | 0.31 ± 0.0003 | 0.31 ± 0.0005 | 0.31 ± 0.0001 | 0.31 ± 0.0003 |
| sauguiin H-6 (8) | 1.98 ± 0.007 | 1.77 ± 0.04 | 2.05 ± 0.03 | 0.85 ± 0.12 | 0.70 ± 0.009 | 0.76 ± 0.021 | 0.72 ± 0.009 | 0.97 ± 0.01 |
| cyanidin-3-O-sophoroside (9) | - | - | - | - | 0.69 ± 0.001 | 0.68 ± 0.005 | 1.96 ± 0.002 | 1.59 ± 0.01 |
| cyanidin-3-O-sambubiside (10) | - | - | - | - | 0.27 ± 0.002 | 0.24 ± 0.05 | 0.65 ± 0.007 | 0.67 ± 0.009 |
| cyanidin-3-O-glucoside (11) | - | - | 0.26 ± 0.005 | 0.21 ± 0.003 | 0.49 ± 0.003 | 0.50 ± 0.009 | 0.88 ± 0.02 | 0.83 ± 0.02 |
| cyanidin-3-O-rutinoside (12) | - | - | - | - | - | 0.21 ± 0.001 | 0.35 ± 0.008 | 0.42 ± 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kobori, R.; Yakami, S.; Kawasaki, T.; Saito, A. Changes in the Polyphenol Content of Red Raspberry Fruits during Ripening. Horticulturae 2021, 7, 569. https://doi.org/10.3390/horticulturae7120569
AMA Style
Kobori R, Yakami S, Kawasaki T, Saito A. Changes in the Polyphenol Content of Red Raspberry Fruits during Ripening. Horticulturae. 2021; 7(12):569. https://doi.org/10.3390/horticulturae7120569
Chicago/Turabian StyleKobori, Ryo, Syuichi Yakami, Takashi Kawasaki, and Akiko Saito. 2021. "Changes in the Polyphenol Content of Red Raspberry Fruits during Ripening" Horticulturae 7, no. 12: 569. https://doi.org/10.3390/horticulturae7120569
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.