
Organogenesis of Plant Tissues in Colchicine Allows Selecting in Field Trial Blueberry (Vaccinium spp. cv Duke) Clones with Commercial Potential
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Colchicine Treatments
2.3. Flow Cytometry
2.4. Acclimatization in Greenhouse Conditions
2.5. Field Trial for Selection of Elite Clone
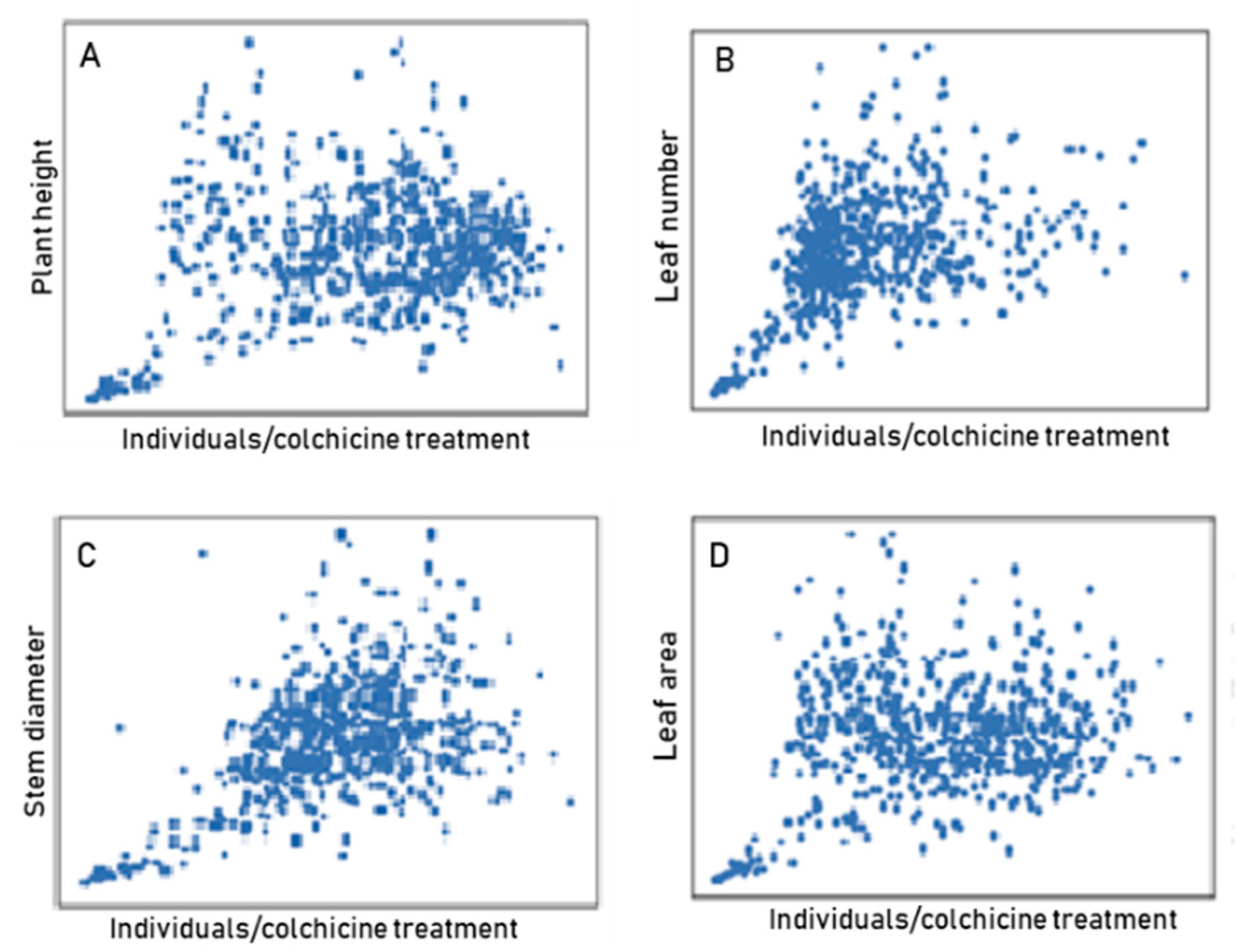
2.6. Characteristic Dispersion and Variability
2.7. Statistical Analysis
3. Results
3.1. Blueberries Production in Colchicine
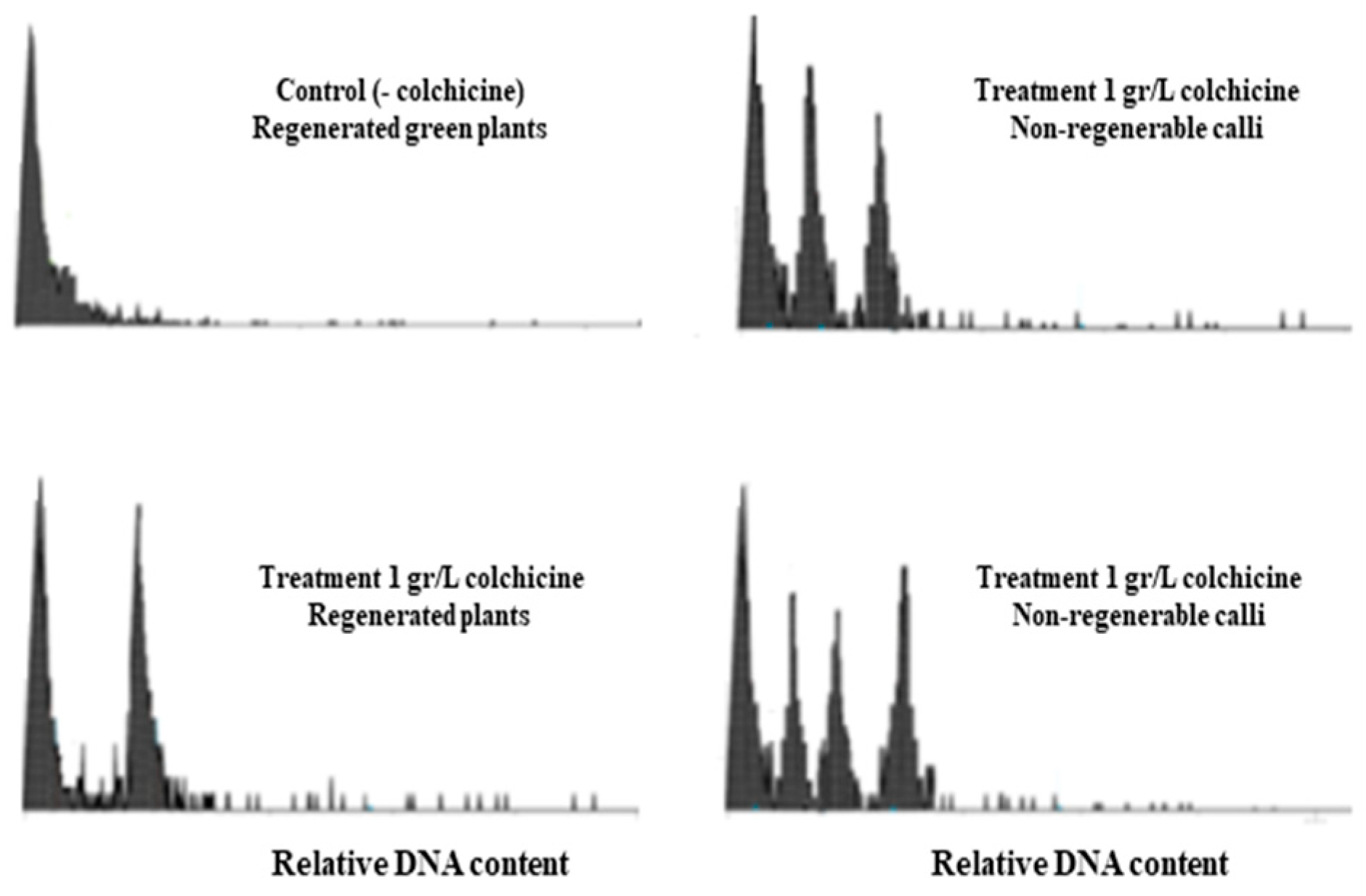
3.2. Flow Cytometry Analysis
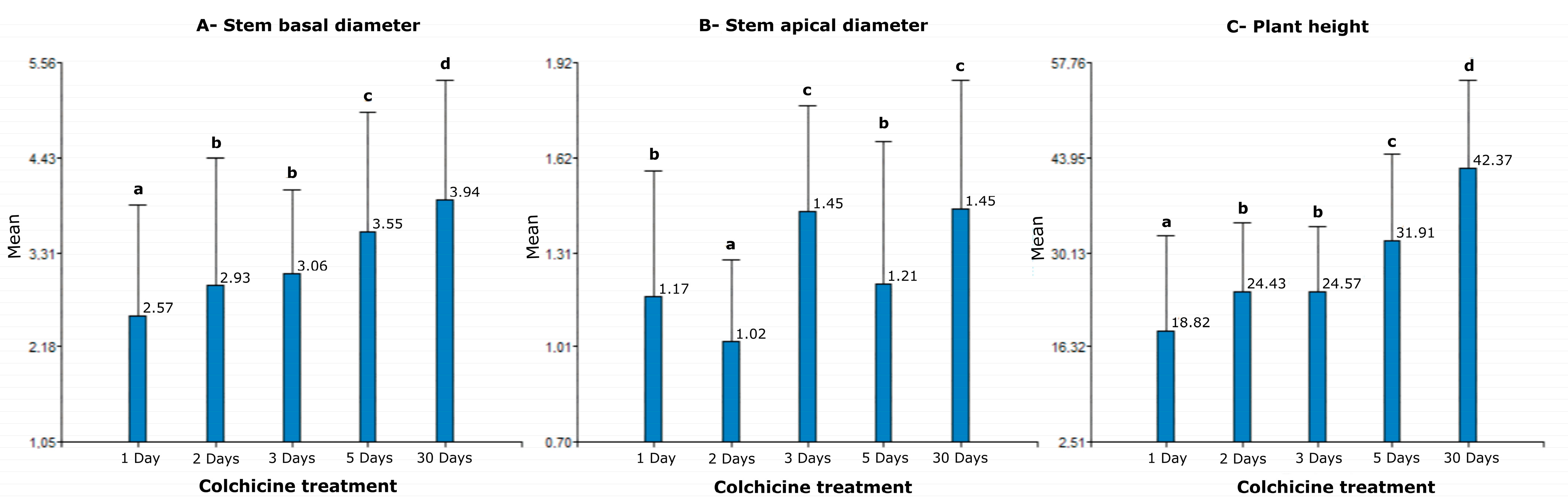
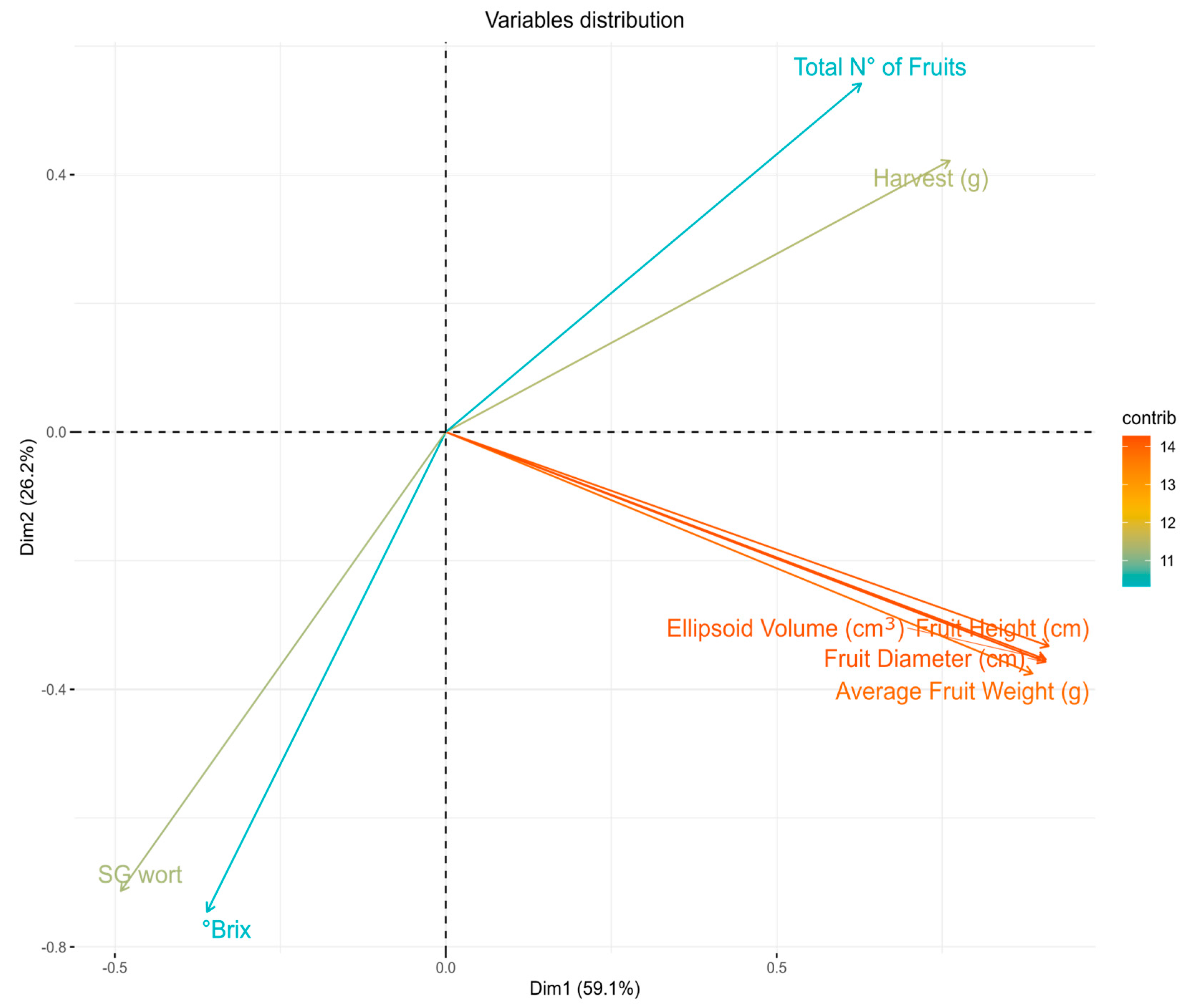
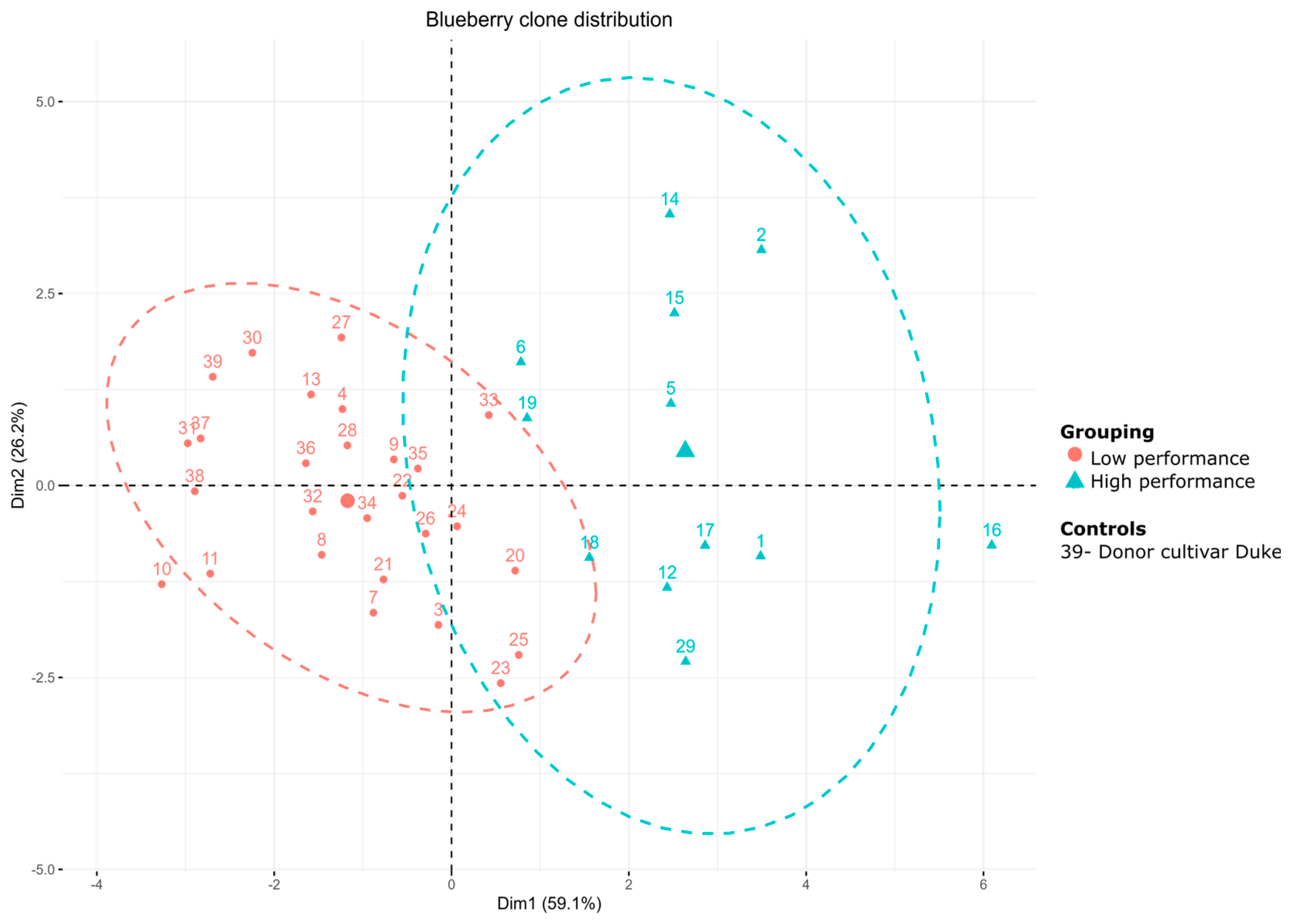
3.3. Evaluation of Agrobotanical Traits in Greenhouse Conditions
4. Discussion
- The donor Duke is a genotype introduced and adapted to Chile based on intense agronomic management practices. Therefore, the conditions of minimum tillage and zero agronomic management constitute an environmental pressure for the expression of their wild genetic potential, being the comparative basis for the selection of improved clones.
- The stressful selection pressures, which, in this case, correspond to the natural environmental conditions that occurred during the three-year period, allow for the selection of genotypes with greater adaptability and plasticity, suitable for the sustainable management of this crop.
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Lobos, G.A.; Hancock, J.F. Breeding Blueberries for a Changing Global Environment: A Review. Front. Plant Sci. 2015, 6, 782. [Google Scholar] [CrossRef]
- Ballington, J.R. Germplasm Resources Available to Meet Future Needs for Blueberry Cultivar Improvement. Fruit Var. J. 1990, 44, 54–62. [Google Scholar]
- Retamales, J.B.; Hancock, J.F. Blueberries; Crop Production Science in Horticulture; CABI: Cambridge, UK, 2012; ISBN 9781845938260. [Google Scholar]
- Luby, J.J. Breeding Cold-Hardy Fruit Crops in Minnesota. HortScience 1991, 26, 507–512. [Google Scholar] [CrossRef]
- Coagra. Arandanos Chilenos. Available online: https://coagra.cl/arandanos-chilenos/ (accessed on 29 August 2022).
- Arencibia, A.D.; Vergara, C.; Quiroz, K.; Carrasco, B.; Bravo, C.; García-Gonzales, R. An Approach for Micropropagation of Blueberry (Vaccinium corymbosum L.) Plants Mediated by Temporary Immersion Bioreactors (TIBs). Am. J. Plant Sci. 2013, 4, 1022–1028. [Google Scholar] [CrossRef]
- Ferrão, L.F.V.; Amadeu, R.R.; Benevenuto, J.; de Bem Oliveira, I.; Munoz, P.R. Genomic Selection in an Outcrossing Autotetraploid Fruit Crop: Lessons from Blueberry Breeding. Front. Plant Sci. 2021, 12, 676326. [Google Scholar] [CrossRef]
- Madani, H.; Escrich, A.; Hosseini, B.; Sanchez-Muñoz, R.; Khojasteh, A.; Palazon, J. Effect of Polyploidy Induction on Natural Metabolite Production in Medicinal Plants. Biomolecules 2021, 11, 899. [Google Scholar] [CrossRef]
- Tavan, M.; Mirjalili, M.H.; Karimzadeh, G. In Vitro Polyploidy Induction: Changes in Morphological, Anatomical and Phytochemical Characteristics of Thymus persicus (Lamiaceae). Plant Cell Tissue Organ Cult. (PCTOC) 2015, 122, 573–583. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on Colchicine Induced Chromosome Doubling for Enhancement of Quality Traits in Ornamental Plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef]
- Gallardo, R.K.; Zhang, Q.; Dossett, M.; Polashock, J.J.; Rodriguez-Saona, C.; Vorsa, N.; Edger, P.P.; Ashrafi, H.; Babiker, E.; Finn, C.E.; et al. Breeding Trait Priorities of the Blueberry Industry in the United States and Canada. HortScience 2018, 53, 1021–1028. [Google Scholar] [CrossRef]
- Ballington, J.R. The Role of Interspecific Hybridization in Blueberry Improvement. Acta Hortic. 2009, 810, 49–60. [Google Scholar] [CrossRef]
- Brevis, P.A.; Bassil, N.V.; Ballington, J.R.; Hancock, J.F. Impact of Wide Hybridization on Highbush Blueberry Breeding. J. Am. Soc. Hortic. Sci. 2008, 133, 427–437. [Google Scholar] [CrossRef]
- Han, X.; Yang, Y.; Han, X.; Ryner, J.T.; Ahmed, E.A.H.; Qi, Y.; Zhong, G.-Y.; Song, G.-Q. CRISPR Cas9- and Cas12a-Mediated GusA Editing in Transgenic Blueberry. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 148, 217–229. [Google Scholar] [CrossRef]
- Song, G.-Q.; Chen, Q. Overexpression of the MADS-Box Gene K-Domain Increases the Yield Potential of Blueberry. Plant Sci. 2018, 276, 22–31. [Google Scholar] [CrossRef]
- Walworth, A.; Song, G.-Q. The Cold-Regulated Genes of Blueberry and Their Response to Overexpression of VcDDF1 in Several Tissues. Int. J. Mol. Sci. 2018, 19, 1553. [Google Scholar] [CrossRef]
- Song, G.-Q.; Walworth, A.; Lin, T.; Chen, Q.; Han, X.; Irina Zaharia, L.; Zhong, G.-Y. VcFT-Induced Mobile Florigenic Signals in Transgenic and Transgrafted Blueberries. Hortic. Res. 2019, 6, 105. [Google Scholar] [CrossRef]
- Schaart, J.G.; van de Wiel, C.C.M.; Smulders, M.J.M. Genome Editing of Polyploid Crops: Prospects, Achievements and Bottlenecks. Transgenic Res. 2021, 30, 337–351. [Google Scholar] [CrossRef]
- Ren, Y.; Jing, Y.; Kang, X. In Vitro Induction of Tetraploid and Resulting Trait Variation in Populus alba × Populus glandulosa Clone 84 K. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 146, 285–296. [Google Scholar] [CrossRef]
- Liu, W.; Zheng, Y.; Song, S.; Huo, B.; Li, D.; Wang, J. In Vitro Induction of Allohexaploid and Resulting Phenotypic Variation in Populus. Plant Cell Tissue Organ Cult. (PCTOC) 2018, 134, 183–192. [Google Scholar] [CrossRef]
- Wang, Z.; Fan, G.; Dong, Y.; Zhai, X.; Deng, M.; Zhao, Z.; Liu, W.; Cao, Y. Implications of Polyploidy Events on the Phenotype, Microstructure, and Proteome of Paulownia australis. PLoS ONE 2017, 12, e0172633. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The Polyploidy and Its Key Role in Plant Breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- do Rêgo, M.M.; do Rêgo, E.R.; Otoni, W.C.; Bruckner, C.H. Epigenetic effects in induced in vitro tetraploids passion fruit (Passiflora edulis sims). In Proceedings of the VI International Symposium on In Vitro Culture and Horticultural Breeding, Brisbane, Australia, 24–28 August 2008; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2009; pp. 167–176. [Google Scholar]
- Kharel, P.; Creech, M.R.; Nguyen, C.D.; Vendrame, W.A.; Muñoz, P.R.; Huo, H. Effect of Explant Type, Culture Medium, and BAP Concentration on in vitro Shoot Development in Highbush Blueberry (Vaccinium corymbosum L.) Cultivars. Vitr. Cell. Dev. Biol. Plant 2022, 58, 1057–1065. [Google Scholar] [CrossRef]
- Sakhanokho, H.F.; Rinehart, T.A.; Stringer, S.J.; Islam-Faridi, M.N.; Pounders, C.T. Variation in nuclear DNA content and chromosome numbers in blueberry. Sci. Hortic. 2018, 233, 108–113. [Google Scholar] [CrossRef]
- Valenzuela, F.; D’Afonseca, V.; Hernández, R.; Gómez, A.; Arencibia, A.D. Validation of Reference Genes in a Population of Blueberry (Vaccinium corymbosum) Plants Regenerated in Colchicine. Plants 2022, 11, 2645. [Google Scholar] [CrossRef]
- Cappelletti, R.; Mezzetti, B. TDZ, 2iP and zeatin in Blueberry (Vaccinium corymbosum L. ’Duke’) in vitro Proliferation and Organogenesis. Acta Hortic. 2016, 1117, 321–324. [Google Scholar] [CrossRef]
- Mazurek, M.; Siekierzyńska, A.; Piechowiak, T.; Spinardi, A.; Litwińczuk, W. Comprehensive Analysis of Highbush Blueberry Plants Propagated in vitro and Conventionally. Int. J. Mol. Sci. 2024, 25, 544. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, Z.; Zhang, C.; Shi, C.; Lyu, L.; Li, W.; Wu, W. Comparative Analysis of the Morphological, Physiological, Proteomic, and Metabolic Mechanisms of the “Biloxi” Blueberry Response to Shade Stress. Front. Plant Sci. 2022, 13, 877789. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, J.; Guo, Z.; Guan, Y.; Qu, G.; Liu, J.; Guo, Y.; Yan, X. Morphological Variation in Cynodon dactylon (L.) Pers., and Its Relationship with the Environment along a Longitudinal Gradient. Hereditas 2020, 157, 4. [Google Scholar] [CrossRef]
- Farinha, N.; Gonçalves, J.C.; Alves, M.A.; Delgado, T.; Anjos, O.; Debiasi, C. In vitro Establishment of Blueberries (Vaccinium corymbosum L.) Using Different Initial Explants. Acta Hortic. 2018, 1224, 145–156. [Google Scholar] [CrossRef]
- Debnath, S.C. A Two-Step Procedure for in Vitro Multiplication of Cloudberry (Rubus chamaemorus L.) Shoots Using Bioreactor. Plant Cell Tissue Organ Cult. (PCTOC) 2007, 88, 185–191. [Google Scholar] [CrossRef]
- Prado, M.J.; Rodriguez, E.; Rey, L.; González, M.V.; Santos, C.; Rey, M. Detection of Somaclonal Variants in Somatic Embryogenesis-Regenerated Plants of Vitis vinifera by Flow Cytometry and Microsatellite Markers. Plant Cell Tissue Organ Cult. (PCTOC) 2010, 103, 49–59. [Google Scholar] [CrossRef]
- LaBelle, C.A.; Massaro, A.; Cortés-Llanos, B.; Sims, C.E.; Allbritton, N.L. Image-Based Live Cell Sorting. Trends Biotechnol. 2021, 39, 613–623. [Google Scholar] [CrossRef]
- Kong, Q.; Yuan, J.; Gao, L.; Zhao, S.; Jiang, W.; Huang, Y.; Bie, Z. Identification of Suitable Reference Genes for Gene Expression Normalization in QRT-PCR Analysis in Watermelon. PLoS ONE 2014, 9, e90612. [Google Scholar] [CrossRef]
- Zhou, K.; Fleet, P.; Nevo, E.; Zhang, X.; Sun, G. Transcriptome Analysis Reveals Plant Response to Colchicine Treatment during on Chromosome Doubling. Sci. Rep. 2017, 7, 8503. [Google Scholar] [CrossRef]
- Redpath, L.E.; Aryal, R.; Lynch, N.; Spencer, J.A.; Hulse-Kemp, A.M.; Ballington, J.R.; Green, J.; Bassil, N.; Hummer, K.; Ranney, T.; et al. Nuclear DNA Contents and Ploidy Levels of North American Vaccinium Species and Interspecific Hybrids. Sci. Hortic. 2022, 297, 110955. [Google Scholar] [CrossRef]
- Marangelli, F.; Pavese, V.; Vaia, G.; Lupo, M.; Bashir, M.A.; Cristofori, V.; Silvestri, C. In vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis. Plants 2022, 11, 2349. [Google Scholar] [CrossRef]
- Tsuda, H.; Kunitake, H.; Yamasaki, M.; Komatsu, H.; Yoshioka, K. Production of Intersectional Hybrids between Colchicine-Induced Tetraploid Shashanbo (Vaccinium bracteatum) and Highbush Blueberry ‘Spartan’. J. Am. Soc. Hortic. Sci. 2013, 138, 317–324. [Google Scholar] [CrossRef]
- Miyashita, C.; Ishikawa, S.; Mii, M. In vitro Induction of the Amphiploid in Interspecific Hybrid of Blueberry (Vaccinium corymbosum × Vaccinium ashei) with Colchicine Treatment. Sci. Hortic. 2009, 122, 375–379. [Google Scholar] [CrossRef]
- Debnath, S.C.; Ghosh, A. Phenotypic Variation and Epigenetic Insight into Tissue Culture Berry Crops. Front. Plant Sci. 2022, 13, 1042726. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explants | Colchicine | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 mg/L (Control) | 0.5 mg/L | 1 mg/L | 2 mg/L | |||||
| VS | NRC | VS | NRC | VS | NRC | VS | NRC | |
| Leaf discs * (upper third) | 47 (47%) | 0 | 21 (21%) | 9 (9%) | 11 (11%) | 16 (16%) | 0 | 0 |
| Nodal segments ** (three buds) | 246 (82%) | 0 | 94 (31%) | 25 (8%) | 36 (12%) | 42 (14%) | 0 | 24 (8%) |
| Treatments | Number of Explants * | Regenerated Shoots (Clone)/% | Percent | Multiplication in Colchicine | Rate | In Vitro Rooted Plants | Percent | Plants Adapted in Greenhouse | Percent ** | Clones Transplanted to Field | Percent **** |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 day | 1000 | 405/13.5 | 13.5 | 2041 | 1:5 | 1884 | 92.3 | 1149 | 60.9 | 15 | 3.7 |
| 2 days | 1000 | 484/16.1 | 16.1 | 3621 | 1:7 | 3427 | 94.6 | 1988 | 56.8 | 18 | 3.7 |
| 3 days | 1000 | 262/8.7 | 8.7 | 1560 | 1:6 | 1492 | 95.6 | 821 | 55 | 21 | 8 |
| 5 days | 1000 | 421/14 | 14 | 3455 | 1:8 | 3112 | 90.1 | 1891 | 60.8 | 49 | 11.6 |
| 30 days | 1000 | 385/12.8 | 12.8 | 2310 | 1:6 | 1708 | 73.9 | 938 | 54.8 | 54 | 14 |
| Total | 5000 | 1957/39.14 | 12 987 | 11 623 | 6 787 | 157 | |||||
| Control (-colchicine) | 1000 | 3426/114.2 *** | 114.2 *** | N.C | N.C | 3912 | 114.1 *** | 3708 | 94.7 | - | - |
| Clone | Weight (g) * | Δ * | Diameter (cm) * | Δ * | Height (cm) * | Δ * | Ellipsoid Volume (cm3) * | Δ * |
|---|---|---|---|---|---|---|---|---|
| 1–436 | 3.11 b | 1.68 | 1.91 a | 0.5 | 1.35 b | 0.29 | 20.63 b | 11.8 |
| 5–437 | 2.35 c | 0.92 | 1.76 c | 0.35 | 1.26 c | 0.2 | 16.35 c | 7.52 |
| 12–102 | 2.92 c | 1.49 | 1.90 b | 0.49 | 1.34 c | 0.28 | 20.26 c | 11.43 |
| 17–421 | 2.84 c | 1.41 | 1.82 c | 0.41 | 1.33 c | 0.27 | 18.45 c | 9.62 |
| 18–420 | 2.67 c | 1.24 | 1.72 c | 0.31 | 1.31 c | 0.25 | 16.23 c | 7.40 |
| 29–440 | 3.12 a | 1.69 | 1.90 b | 0.49 | 1.38 a | 0.32 | 20.87 a | 12.04 |
| (39) Donor ** | 1.43 d | - | 1.41 d | - | 1.06 d | - | 8.83 d | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández, R.; López, A.; Valenzuela, B.; D’Afonseca, V.; Gomez, A.; Arencibia, A.D. Organogenesis of Plant Tissues in Colchicine Allows Selecting in Field Trial Blueberry (Vaccinium spp. cv Duke) Clones with Commercial Potential. Horticulturae 2024, 10, 283. https://doi.org/10.3390/horticulturae10030283
Hernández R, López A, Valenzuela B, D’Afonseca V, Gomez A, Arencibia AD. Organogenesis of Plant Tissues in Colchicine Allows Selecting in Field Trial Blueberry (Vaccinium spp. cv Duke) Clones with Commercial Potential. Horticulturae. 2024; 10(3):283. https://doi.org/10.3390/horticulturae10030283
Chicago/Turabian StyleHernández, Ricardo, Alan López, Bárbara Valenzuela, Vivian D’Afonseca, Aleydis Gomez, and Ariel D. Arencibia. 2024. "Organogenesis of Plant Tissues in Colchicine Allows Selecting in Field Trial Blueberry (Vaccinium spp. cv Duke) Clones with Commercial Potential" Horticulturae 10, no. 3: 283. https://doi.org/10.3390/horticulturae10030283