Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials


G.K. Skryabin Institute of Biochemistry and Physiology of Microorganisms, Russian Academy of Sciences, pr-t Nauki, 5, 142290 Pushchino, Russia
*
Author to whom correspondence should be addressed.
Fermentation 2018, 4(2), 36; https://doi.org/10.3390/fermentation4020036
Submission received: 28 April 2018
/
Revised: 11 May 2018
/
Accepted: 15 May 2018
/
Published: 17 May 2018
(This article belongs to the Special Issue Yeast Biotechnology 2.0)
Abstract
:The world market of citric acid (CA) is one of the largest and fastest growing markets in the biotechnological industry. Microbiological processes for CA production have usually used the mycelial fungi Aspergillus niger as a producer and molasses as a carbon source. In this paper, we propose methods for CA production from renewable carbon substrates (rapeseed oil, glucose, glycerol, ethanol, glycerol-containing waste of biodiesel industry and glucose-containing aspen waste) by the mutant strain Yarrowia lipolytica NG40/UV5. It was revealed that Y. lipolytica grew and synthesized CA using all tested raw materials. The obtained results are sufficient for industrial use of most of the raw materials studied for CA production. Using rapeseed oil, ethanol and raw glycerol (which is an important feedstock of biodiesel production), a high CA production (100–140 g L−1) was achieved.
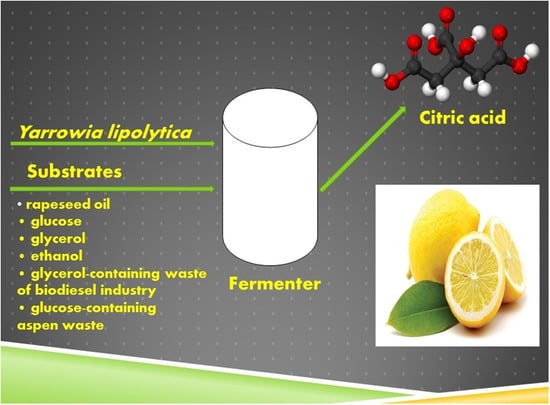
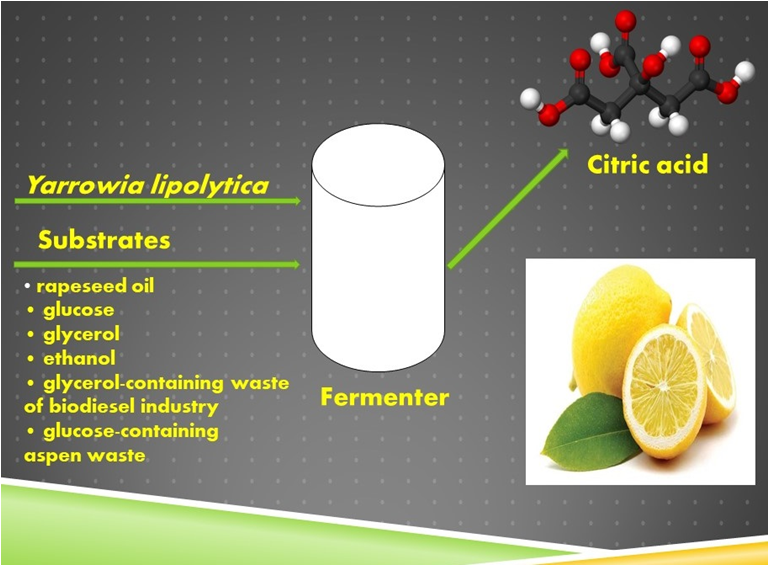
1. Introduction
Citric acid (CA) and its salts are widely used as an acidulate, flavoring agent and antioxidant in the production of beverages and confectionery, in infant formula, as well as in the chemical, pharmaceutical, electronic, defense, and other industries. The volume of citric acid globally exceeds two million tons per year and its production is annually increased by 5% [1].
Modern technologies of citric acid (CA) production are based on using various mutant strains of the mycelial fungi Aspergillus niger as a producer and molasses as a raw material. CA production by fungi is a complicated and environmentally unsafe process; as a result of its implementation, a large number of both liquid effluents containing mineral acids, ballast organic substances, cyanides, and solid wastes, primarily gypsum, are accumulated. Moreover, A. niger is an opportunistic pathogenic fungi and can cause allergic diseases and aspergillosis [2,3].
Over the past 40 years, the interest of researchers has focused on yeast as a producer of CA; the yeast Yarrowia lipolytica has been the most used CA producer [1,2,3]. Initially, this kind of yeast attracted the attention of researchers due to its ability to grow and synthesize CA in media with n alkanes—an available and cheap substrate [2]. However, due to changes in the world oil market, the use of this substrate has become economically unprofitable. In this regard, it is of interest to use other types of raw materials.
The choice of raw materials for developing CA biotechnology is determined by factors such as renewability, ability of the producer to assimilate the substrate with a high conversion rate, consumption value and cost price of the target product. To carry out fermentation processes, in addition to very expensive food raw materials, such as glucose [4,5,6,7] and plant oils [8,9,10], much cheaper substrates which are waste products of various industries, such as glycerol-containing waste of the biodiesel industry [9,10,11,12,13,14,15,16,17], glucose-containing wood hydrolysates [18,19], olive mill waste-water [20], and inulin [21] are used. Ethanol, a water-soluble individual compound which ensures the formation of a pure product and facilitates the isolation process, is also of great importance [22,23]. In the works of the above-mentioned researchers it was shown that all these substrates are promising for CA production and the use of glycerol-containing waste of biodiesel industry and glucose-containing wood hydrolyzates can increase the profitability of CA production process.
The aim of this work was a comparative study of CA production by the yeast Y. lipolytica on different types of renewable raw materials.
2. Materials and Methods
2.1. Microorganisms and Chemicals
2.2. Media and Cultivation Conditions
All experiments were done using the same equipment, nutrient medium and cultivation conditions. Strain Y. lipolytica NG40/UV5 was cultivated in a 10-L ANKUM-2M fermenter (Pushchino, Russia) with an initial volume of 5 L. The medium contained (g L−1): carbon source: (NH4)2SO4, 6; MgSO4·7H2O, 1.4; NaCl, 0.5; Ca(NO3)2, 0.8; KH2PO4, 2.0; K2HPO4, 0.2; Difco yeast extract (BD Diagnostic Systems, Sparks, MD, USA), 1.0; trace elements (mg L−1): FeSO4 × 7H2O, 14.9; MnSO4 × 4H2O, 0.2; ZnSO4 × 7H2O, 8.1; CuSO4 × 5H2O, 3.9. The fermentation conditions were maintained automatically at a constant level: Temperature 28−0.5 °C; pH 4.5.0−0.1; pO2 50% (of air saturation); agitation rate of 800 rpm. Pulsed addition of carbon source (by 2–20 g L−1) depending on the carbon source used was performed as the pO2 value changed by 10%. Cultivation was continued for 6 days.
2.3. Assays
Biomass, concentration of CA, isocitric acid (ICA) and other organic acids were determined as described previously [7].
2.4. Calculations
Earlier, it was found that the mass yield of CA production (YCA), expressed in g of CA per g of carbon source, and the fermenter productivity, expressed in g (L·h)−1 were influenced by the medium dilution due to the addition of NaOH solution for maintaining a constant pH value [9,17]. In this regard, the total amount of CA in the culture broth at the end of the fermentation was used to calculate YCA and fermenter productivity. Formulas for calculation of YCA value and fermenter productivity were described earlier [9,17].
All the data presented are the mean values of three experiments and two measurements for each experiment; standard deviations were calculated (S.D. < 10%).
3. Results and Discussion
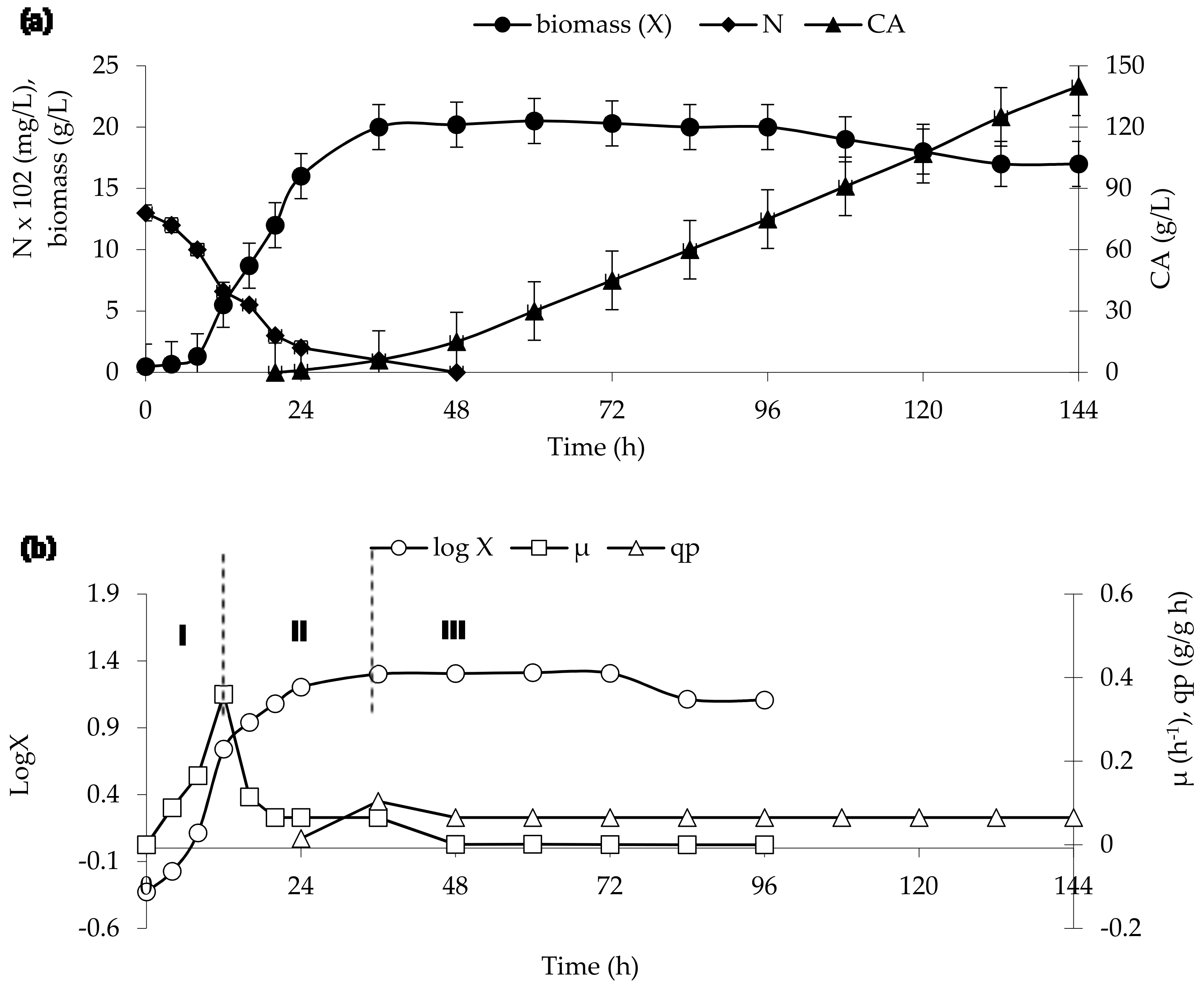
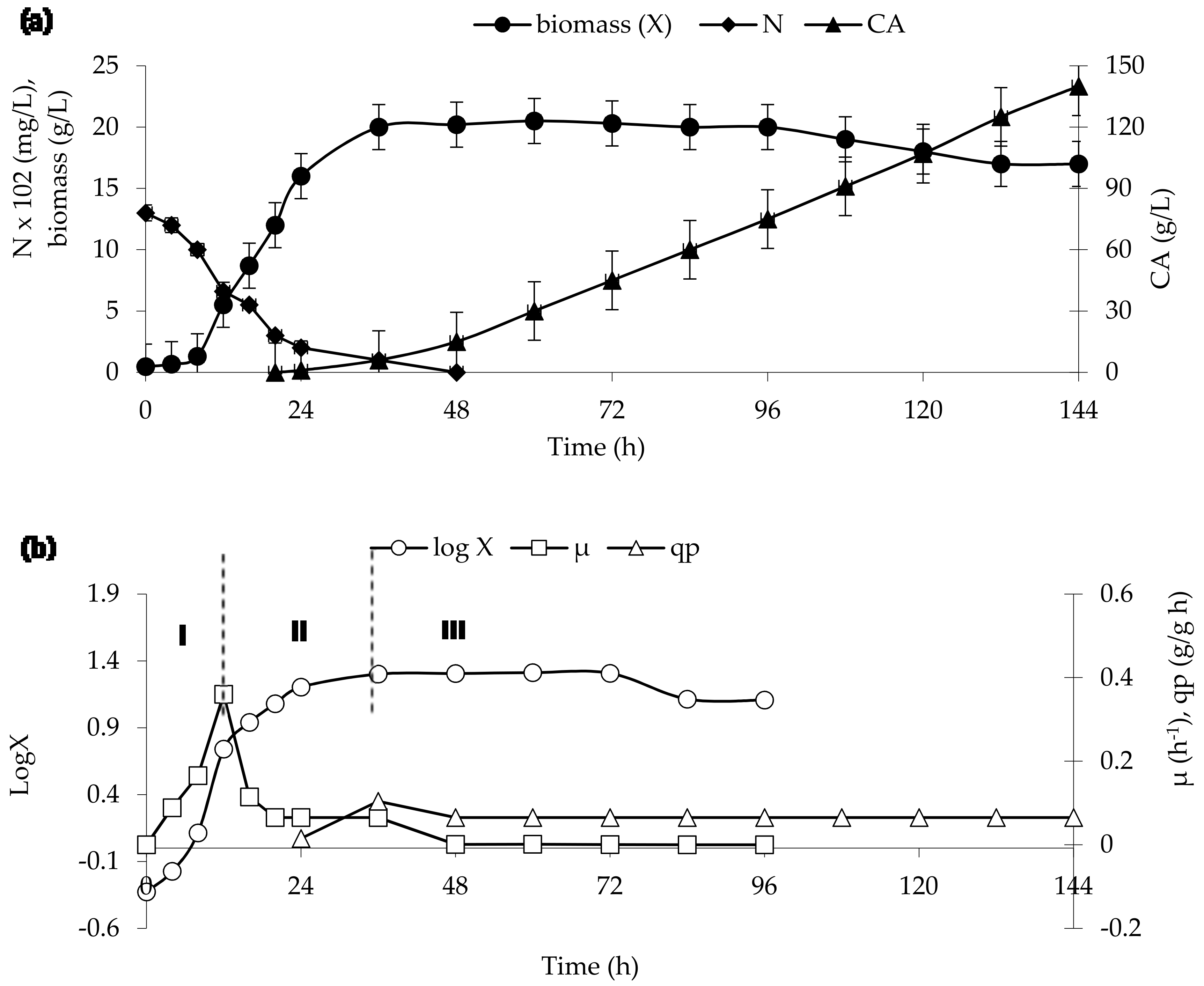
The dynamics of nitrogen consumption and the accumulation of biomass and CA by Y. lipolytica NG40/UV5 grown on rapeseed oil are shown in Figure 1a, while the logarithmic growth curve (µ) and the specific rate of biosynthesis of CA (qp) are shown in Figure 1b. As it can be seen in the latter figure, the growth curve had an exponential phase (phase I) lasting for 12 h, growth retardation phase (phase II) lasting from 12 to 36 h of cultivation, and stationary phase (phase III) lasting from 36 h to the end of cultivation. The retardation of growth coincided with the exhaustion of nitrogen from the medium. The specific growth rate attained a maximum (µmax = 0.360 h−1) in the exponential growth phase (12 h of cultivation). This value of µmax was more than two times higher than that of the other CA-producing strain Y. lipolytica (0.17–0.22 h−1) [4,8]. After 12 h of cultivation, µ gradually decreased to zero after 48 h of cultivation. The excretion of CA did not occur in the exponential growth phase but became active in the growth retardation and stationary phases. Within this cultivation period, the specific rate of CA production (qp) was between 0.065 and 0.104 g CA/g·h. At the end of cultivation (144 h), the strain produced 140 g L−1 CA and 5.3 g L−1 ICA (data not shown) with CA:ICA ratio of 26.4:1. The CA production yield YCA was 1.5 g g−1; the fermenter productivity was calculated to be 1.46 g (L·h)−1 with account for the dilution factor.
The data of the accumulation of biomass and CA by Y. lipolytica NG40/UV5 grown on other substrates compared to rapeseed oil are shown in Table 1.
As seen in Table 1, at the end of cultivation (144 h), Y. lipolytica NG40/UV5 produced 100.8 g L−1 CA and 4.9 g L−1 ICA with CA:ICA ratio of 20.6:1 in the medium containing glucose. The CA production yield YCA was 0.80 g g−1; the fermenter productivity was calculated to be 1.05 g (L·h)−1.
As seen in Table 1, Y. lipolytica NG40/UV5 only produced 31.2 g L−1 CA and 7.84 g L−1 ICA with CA:ICA ratio of 4:1 in the medium containing glucose-containing aspen waste. The CA production yield YCA was 0.50 g g−1; the fermenter productivity was calculated to be 0.325 g (L·h)−1.
As it can be seen from the data in Table 1, the mutant grows perfectly and synthesizes CA both in a medium with pure glycerol and in a medium with biodiesel-derived glycerol. Y. lipolytica NG40/UV5 produced 87 g L−1 CA with a ratio of CA to ICA of 6.7:1. The application of waste glycerol for Y. lipolytica NG40/UV5 cultivation increased CA production by 15% (up to 100 g L−1) compared to that obtained from pure glycerol; the CA to ICA ratio was 7.7:1. The fermenter productivity was high and reached 0.906 and 1.04 (g (L·h)−1) in the media with pure- and biodiesel-derived glycerol, respectively. The mass yield (YCA) reached 0.64 and 0.9 g/g in the media with pure- and biodiesel-derived glycerol, respectively.
As seen in Table 1, Y. lipolytica NG40/UV5 produced 106.7 g L−1 CA and 15 g L−1 ICA with CA:ICA ratio of 7.1:1 in the medium containing ethanol. The CA production yield YCA was 0.87 g g−1; the fermenter productivity was calculated to be 1.32 g/L·h.
Comparative data on the most efficient processes of CA production by yeasts Y. lipolytica from various carbohydrate-containing substrates are given in Table 2. As seen in this table, in the experiments with wild and mutant strains, Y. lipolytica produced CA in industrially sufficient amounts. For instance, in the mutant strain Saccharomycopsis lipolytica NTG9 grown on rapeseed oil, the CA concentration reached 152.3 g L−1 with the yield (YCA) of 1.5 g g−1 [25]. Aurich et al. [26] obtained a CA concentration of 198 g L−1 with the yield (YCA) of 1.16 g g−1, which was achieved after a 300 h fed-batch cultivation of the wild strain Y. lipolytica H181. The wild strains Y. lipolytica H222 and Y. lipolytica W29, grown on glucose, produced 41 g L−1 [4] and 49 g L−1 of CA, respectively [27]. Recently, we found that the wild strain Y. lipolytica VKM Y 2373, cultivated in a medium with glucose under cell growth limitation using nitrogen, phosphorus and sulfur, produced CA at a level of 80–85 g L−1 with a yield of 0.70–0.75 g g−1 [7]. The overexpression of gene PYR encoding pyruvate carboxylase in Y. lipolytica, resulted in the production of CA at a level of 95–111.1 g L−1 with the yield (YCA) of 0.75–0.93 g g−1 [5,6]. Strain Y. lipolytica ACA-DS 50109 cultivated on glucose and olive mill wastewaters produced CA (28.9 g L−1) with the product yield (YCA) of 0.53 g g−1 [28]. Later, the authors of the last article improved the process of CA production up to 52.0 g L−1 with the product yield (YCA) of 0.64 g g−1 using strain Y. lipolytica ACA-YC 5033, which was also able to remove harmful phenolic compounds from olive mill wastewaters [29]. The glycerol-grown yeast Y. lipolytica NRRL YB-423 produced 21.6 g L−1 of CA with mass yield of 0.54 g g−1 [12]; strain Y. lipolytica ACA-DC 50109 synthesized 62.5 g L−1 of CA with mass yield of 0.56 g g−1 from raw glycerol [20], while the recombinant strain Y. lipolytica NCYC3825 was able to produce 58 g L−1 of CA [30]. Earlier, we indicated that the other mutant Y. lipolytica NG40/UV7 synthesized, CA (122.2 g L−1) with the yield of 0.95 g g−1. The high CA production (up to 140 g L−1) has also been reported for acetate-negative mutant Y. lipolytica Wratislavia AWG7, grown on crude glycerol [13] and recombinant strain Y. lipolytica H222-S4 (p67ICL1), harboring the invertase encoding ScSUC2 gene of Saccharomyces cerevisiae under inducible XPR2 promoter control and multiple ICL1 copies, cultivated on sucrose [31].
It should be noted that plant raw materials, such as wood, straw, and agricultural products processing waste, are inexpensive, accessible, renewable, and environmentally friendly substrates for microbiological synthesis of practically valuable compounds. However, the effective conversion of these substrates into easily assimilable carbohydrates (glucose, xylose, and higher glucose-containing polymers) is a difficult task. The traditional technologies of hydrolysis of plant raw materials with the use of strong acids and alkalis are associated with the formation of by-products that inhibit the growth of microorganisms and biosynthesis of target substances. Therefore, recombinant producers, effectively assimilating plant raw materials, were developed. For instance, Ledesmo-Amaro et al. (2016) engineered Y. lipolytica able to metabolize xylose to produce CA and lipids. The overexpression of xylose reductase, xylitol dehydrogenase and xylulokinase resulted in a production of 80 g L−1 of CA from xylose by mutant strain Y. lipolytica [19].
Rakicka et al. (2016) reported the production from inulin by engineered strain Y. lipolytica Wratislavia K1. The overexpression of the INU1 gene from Kluyveromyces marxianus coding inulinase resulted in the effective hydrolysis of inulin by mutant and the production of a high amount of CA (105.2 g L−1 from 200 g L−1 inulin) [21].
4. Conclusions
The results of the experiments indicated that the mutant Y. lipolytica NG40/UV5 was able to grow and synthesize CA on media containing all types of the investigated renewable raw materials. However, it should be noted that the accumulation of the by-product of fermentation—ICA—was even very high in four of the six substrates investigated (glucose-containing aspen waste, glycerol, glycerol waste of biodiesel industry and ethanol). The best results were obtained using rapeseed oil (140 g L−1 CA; CA:ICA ratio of 26.2:1; mass yield of CA production (YCA) of 1.5 g g−1, and fermenter productivity of 1.46 g (L·h)−1). However, economic considerations may lead to the fact that using a cheaper and less pure substrate (glycerol waste of biodiesel industry; 100 g L−1 CA; CA:ICA ratio of 7.7:1; mass yield of CA production (YCA) of 0.9 g g−1, and fermenter productivity of 1.04 g (L·h)−1) would be preferable on a production scale.
Author Contributions
I.G.M. conceived and designed the experiments, wrote the paper; S.V.K. and J.N.L. performed the experiments.
Funding
The reported study was funded by Russian Foundation for Basic Research (RFBR) according to the research project № 16-08-00702.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cavallo, E.; Charreau, H.; Cerrutti, P.; Foresti, M.L. Yarrowia lipolytica: A model yeast for citric acid production. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Anastassiadis, S.; Morgunov, I.G.; Kamzolova, S.V.; Finogenova, T.V. Citric acid production patent review. Recent Pat. Biotechnol. 2008, 2, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Finogenova, T.V.; Morgunov, I.G.; Kamzolova, S.V.; Chernyavskaya, O.G. Organic acid production by the yeast Yarrowia lipolytica: A review of prospects. Appl. Biochem. Microbiol. 2005, 41, 418–425. [Google Scholar] [CrossRef]
- Moeller, L.; Strehlitz, B.; Aurich, A.; Zehnsdorf, A.; Bley, T. Optimization of citric acid production from glucose by Yarrowia lipolytica. Eng. Life Sci. 2007, 7, 504–511. [Google Scholar] [CrossRef]
- Tan, M.J.; Chen, X.; Wang, Y.K.; Liu, G.L.; Chi, Z.M. Enhanced citric acid production by a yeast Yarrowia lipolytica over-expressing a pyruvate carboxylase gene. Bioprocess Biosyst. Eng. 2016, 39, 1289–1296. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.Y.; Lu, Y.; Chi, Z.; Liu, G.L.; Zhao, S.F.; Jiang, H.; Chi, Z.M. Cloning and characterization of a pyruvate carboxylase from Penicillium rubens and overexpression of the gene in the yeast Yarrowia lipolytica for enhanced citric acid production. Mar. Biotechnol. 2016, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Morgunov, I.G. Metabolic peculiarities of the citric acid overproduction from glucose in yeasts Yarrowia lipolytica. Bioresour. Technol. 2017, 243, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Morgunov, I.G.; Aurich, A.; Perevoznikova, O.A.; Shishkanova, N.V.; Stottmeister, U.; Finogenova, T.V. Lipase secretion and citric acid production in Yarrowia lipolytica yeast grown on animal and vegetable fat. Food Technol. Biotechnol. 2005, 43, 113–122. [Google Scholar]
- Kamzolova, S.V.; Lunina, J.N.; Morgunov, I.G. Biochemistry of citric acid production from rapeseed oil by Yarrowia lipolytica yeast. J. Am. Oil Chem. Soc. 2011, 88, 1965–1976. [Google Scholar] [CrossRef]
- Aurich, A.; Specht, R.; Müller, R.A.; Stottmeister, U.; Yovkova, V.; Otto, C.; Holz, M.; Barth, G.; Heretsch, P.; Thomas, F.A.; et al. Microbiologically Produced Carboxylic Acids Used as Building Blocks in Organic Synthesis. In Reprogramming Microbial Metabolic Pathways; Wang, X., Chen, J., Quinn, P., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 391–424. Available online: https://link.springer.com/chapter/10.1007%2F978-94-007-5055-5_19 (accessed on 28 September 2012).
- Rymowicz, W.; Rywińska, A.; Żarowska, B.; Juszczyk, P. Citric acid production from raw glycerol by acetate mutants of Yarrowia lipolytica. Chem. Pap. 2006, 60, 391–395. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzym. Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Rywińska, A.; Rymowicz, W.; Źarowska, B.; Wojtatowicz, M. Biosynthesis of citric acid from glycerol by acetate mutants of Yarrowia lipolytica in fed-batch fermentation. Food Technol. Biotechnol. 2009, 47, 1–6. [Google Scholar]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic activities of biotechnological interest in Yarrowia lipolytica grown on glycerol in repeated batch cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. The citric acid production from raw glycerol by Yarrowia lipolytica yeast and its regulation. Appl. Microbiol. Biotechnol. 2013, 97, 7387–7397. [Google Scholar] [CrossRef] [PubMed]
- Rywinska, A.; Juszczyk, P.; Wojtatowicz, M.; Robak, M.; Lazar, Z.; Tomaszewska, L.; Rymowicz, W. Glycerol as a promising substrate for Yarrowia lipolytica biotechnological applications. Biomass Bioenergy 2013, 48, 148–166. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V. Physiologo-biochemical characteristics of citrate-producing yeast Yarrowia lipolytica grown on glycerol-containing waste of biodiesel industry. Appl. Microbiol. Biotechnol. 2015, 99, 6443–6450. [Google Scholar] [CrossRef] [PubMed]
- Wojtatowicz, M.; Rymowicz, W.; Kautola, H. Comparison of different strains of the yeast Yarrowia lipolytica for citric acid production from glucose hydrol. Appl. Biochem. Biotechnol. 1991, 31, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Amaro, R.; Lazar, Z.; Rakicka, M.; Guo, Z.; Fouchard, F.; Coq, A.C.; Nicaud, J.M. Metabolic engineering of Yarrowia lipolytica to produce chemicals and fuels from xylose. Metab. Eng. 2016, 38, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Fakas, S.; Fick, M.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Biotechnological valorisation of raw glycerol discharged after biodiesel (fatty acid methyl-esters) manufacturing process: Production of 1,3-propanediol, citric acid and single oil. Biomass Bioengergy 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Rakicka, M.; Lazar, Z.; Rywinska, A.; Rymowicz, W. Efficient utilization of inulin and glycerol as fermentation substrates in erythritol and citric acid production using expressing inulinase. Chem. Pap. 2016, 70, 1452–1459. [Google Scholar] [CrossRef]
- Stephanopoulos, G. Challenges in engineering microbes for biofuels production. Science 2007, 315, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Weusthuis, R.A.; Aarts, J.M.M.J.G.; Sanders, J.P.M. From biofuel to bioproduct: Is bioethanol a suitable fermentation feedstock for synthesis of bulk chemicals? Biofuels Bioprod. Biorefin. 2011, 5, 486–494. [Google Scholar] [CrossRef]
- Finogenova, T.V.; Puntus, I.F.; Kamzolova, S.V.; Lunina, I.N.; Monastyrskaia, S.E.; Morgunov, I.G.; Boronin, A.M. Obtaining of the mutant Yarrowia lipolytica strains producing citric acid from glucose. Prikl. Biokhimiia Mikrobiol. 2008, 44, 219–224. [Google Scholar]
- Good, D.W.; Droniuk, R.; Lawford, R.G.; Fein, J.E. Isolation and characterization of a Saccharomycopsis lipolytica mutant showing increased production of citric acid from canola oil. Can. J. Microbiol. 1985, 31, 436–440. [Google Scholar] [CrossRef]
- Aurich, A.; Förster, A.; Mauesberger, S.; Barth, G.; Stottmeister, U. Citric acid production from renewable resources by Yarrowia lipolytica. Biotechnol. Adv. 2003, 21, 454–455. [Google Scholar]
- Papanikolaou, S.; Chatzifragkou, A.; Fakas, S.; Galiotou-Panayotou, M.; Komaitis, M.; Nicaud, J.M.; Aggelis, G. Biosynthesis of lipids and organic acids by Yarrowia lipolytica strains cultivated on glucose. Eur. J. Lipid Sci. Technol. 2009, 111, 1221–1232. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Fakas, S.; Komaitis, M.; Aggelis, G. Citric acid production by Yarrowia lipolytica cultivated on olive-mill wastewater-based media. Bioresour. Technol. 2008, 99, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Sarris, D.; Stoforos, N.G.; Mallouchos, A.; Kookos, I.K.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Production of added-value metabolites by Yarrowia lipolytica growing in olive mill wastewater-based media under aseptic and non-aseptic conditions. Eng. Life Sci. 2017, 17, 695–709. [Google Scholar] [CrossRef]
- Celińska, E.; Grajek, W. A novel multigene expression construct for modification of glycerol metabolism in Yarrowia lipolytica. Microb. Cell Fact. 2013, 12, 102. [Google Scholar] [CrossRef] [PubMed]
- Förster, A.; Aurich, A.; Mauersberger, S.; Barth, G. Citric acid production from sucrose using a recombinant strain of the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2007, 75, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Time courses of nitrogen consumption, biomass accumulation, and citric acid production in Y. lipolytica grown on rapeseed oil (a) and calculated parameters of the process (b): I—the exponential cell growth; II—the cell growth retardation; III—the stationary phase.
Figure 1.
Time courses of nitrogen consumption, biomass accumulation, and citric acid production in Y. lipolytica grown on rapeseed oil (a) and calculated parameters of the process (b): I—the exponential cell growth; II—the cell growth retardation; III—the stationary phase.

{kind=link}
{kind=link}
Table 1.
Citric acid production by Y. lipolytica on various carbon sources.
| Substrates | Biomass (g L−1) | CA (g L−1) | ICA (g L−1) | CA:ICA | Productivity (g (L·h)−1) | YCA (g g−1) |
|---|---|---|---|---|---|---|
| Rapeseed oil | 17.0 ± 1.1 | 140.0 ± 5.0 | 5.3 ± 0.8 | 26.2:1 | 1.46 | 1.50 |
| Glucose | 18.7 ± 1.3 | 100.8 ± 9.2 | 4.9 ± 0.9 | 20.6:1 | 1.05 | 0.80 |
| Glucose-containing aspen waste | 5.6 ± 0.8 | 31.2 ± 2.1 | 7.84 ± 0.9 | 4:1 | 0.325 | 0.50 |
| Glycerol | 16.8 ± 1.1 | 87 ± 6.4 | 13 ± 1.1 | 6.7:1 | 0.906 | 0.64 |
| Glycerol waste of biodiesel industry | 20.0 ± 1.8 | 100 ± 3.4 | 15 ± 1.2 | 7.7:1 | 1.04 | 0.90 |
| Ethanol | 15.3 ± 1.4 | 106.7 ± 2.7 | 15 ± 1.4 | 7.1:1 | 1.32 | 0.87 |
Table 2.
Comparative data of the processes of CA production from various substrates using Y. lipolytica strains.
Table 2.
Comparative data of the processes of CA production from various substrates using Y. lipolytica strains.
| Strain | Substrate | Characteristics of Strain | CA (g L−1) | YCA (g g−1) | References |
|---|---|---|---|---|---|
| S. lipolytica NTG9 | canola oil | mutant/nitrosoguanidine | 152.3 | 1.50 | [25] |
| Y. lipolytica H181 | sunflower oil | wild type | 198.0 | 1.16 | [26] |
| Y. lipolytica H222 | glucose | wild type | 41.0 | 0.55 | [4] |
| Y. lipolytica W29l | wild type | 49.0 | 0.85 | [27] | |
| Y. lipolytica VKM Y 2373 | wild type | 80–85 | 0.70–0.75 | [7] | |
| Y. lipolytica PG86 | PYC gene expression | 95.0 | 0.75 | [5] | |
| Y. lipolytica PR32 | PYC gene expression | 111.1 | 0.93 | [6] | |
| Y. lipolytica ACA-DS 50109 | glucose + olive mill wastewaters | wild type | 28.9 | 0.53 | [28] |
| Y. lipolytica ACA-YC 5033 | wild type | 52.0 | 0.64 | [29] | |
| Y. lipolytica NRRL YB-423 | glycerol | wild type | 21.6 | 0.54 | [12] |
| Y. lipolytica ACA-DC 50109 | raw glycerol | wild type | 62.5 | 0.56 | [20] |
| Y. lipolytica NCYC 3825 | raw glycerol | multigene expression | 58.8 | 0.17 | [30] |
| Y. lipolytica NG40/UV7 | raw glycerol | mutant/nitrosoguanidine/UV | 122.2 | 0.95 | [17] |
| Y. lipolytica Wratislavia AWG7 | raw glycerol | mutant/acetate− | 139.0 | 0.70 | [13] |
| Y.lipolytica H222-S4 (p67ICL1) | sucrose | ScSUC2/ICL1 | 127–140 | 0.75–0.82 | [31] |
| Y. lipolytica XYL+ | xylose | XYL gene expression | 80.0 | 0.53 | [19] |
| Y. lipolytica Wratislavia K1 | inulin | INU1 gene expression | 105.2 | 0.53 | [21] |
| Y. lipolytica NG40/UV5 | rapeseed oil | mutant/nitrosoguanidine/UV | 140.0 | 1.5 | Present study |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials. Fermentation 2018, 4, 36. https://doi.org/10.3390/fermentation4020036
AMA Style
Morgunov IG, Kamzolova SV, Lunina JN. Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials. Fermentation. 2018; 4(2):36. https://doi.org/10.3390/fermentation4020036
Chicago/Turabian StyleMorgunov, Igor G., Svetlana V. Kamzolova, and Julia N. Lunina. 2018. "Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials" Fermentation 4, no. 2: 36. https://doi.org/10.3390/fermentation4020036
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.