Principal Component Analysis for Clustering Probiotic-Fortified Beverage Matrices Efficient in Elimination of Shigella sp.

Abstract
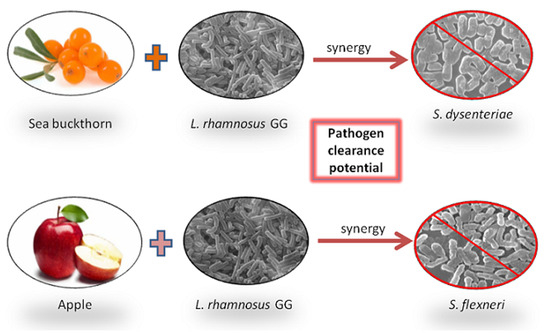
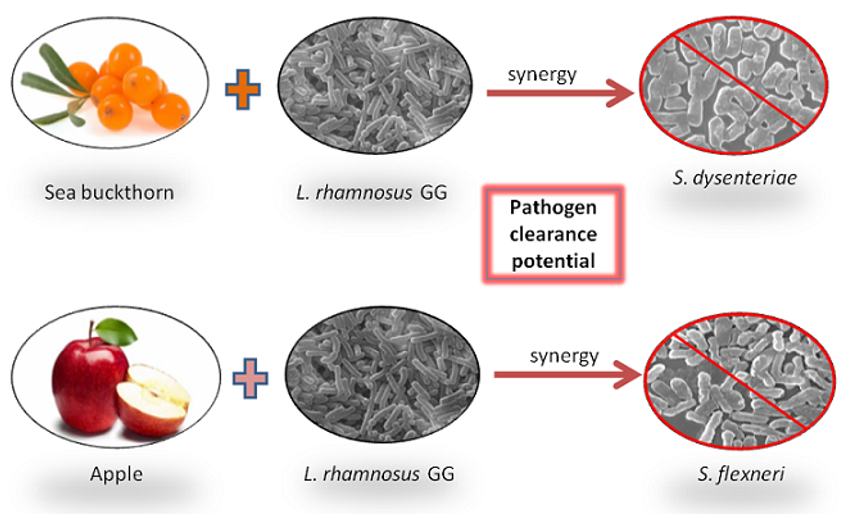
:
1. Introduction
2. Materials and Methods
2.1. Raw Materials and Bacterial Strains
2.2. Development of Juice Matrices
2.3. Evaluation of Anti-Shigella Potential of the Probiotic Beverages during Shelf Storage
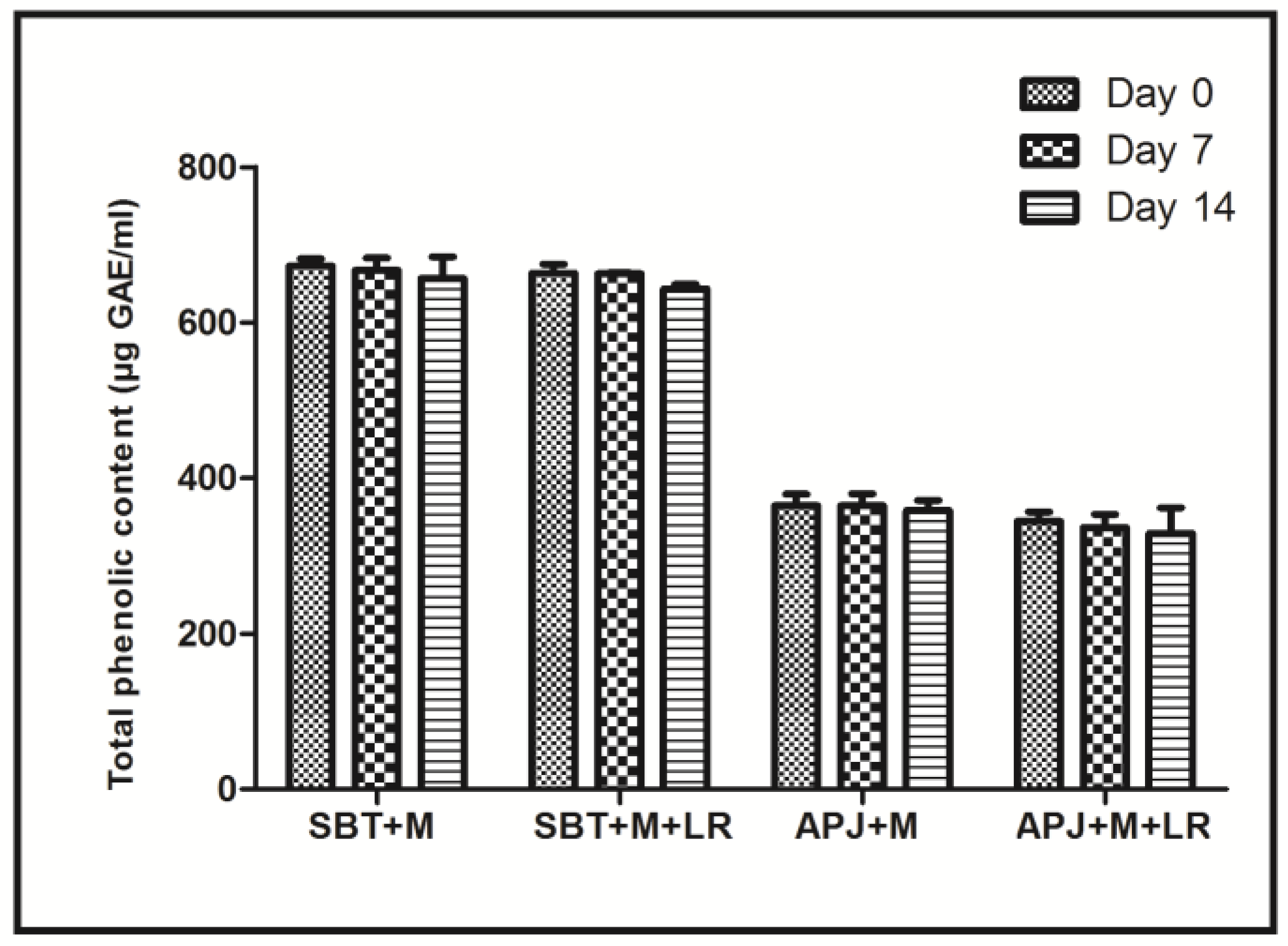
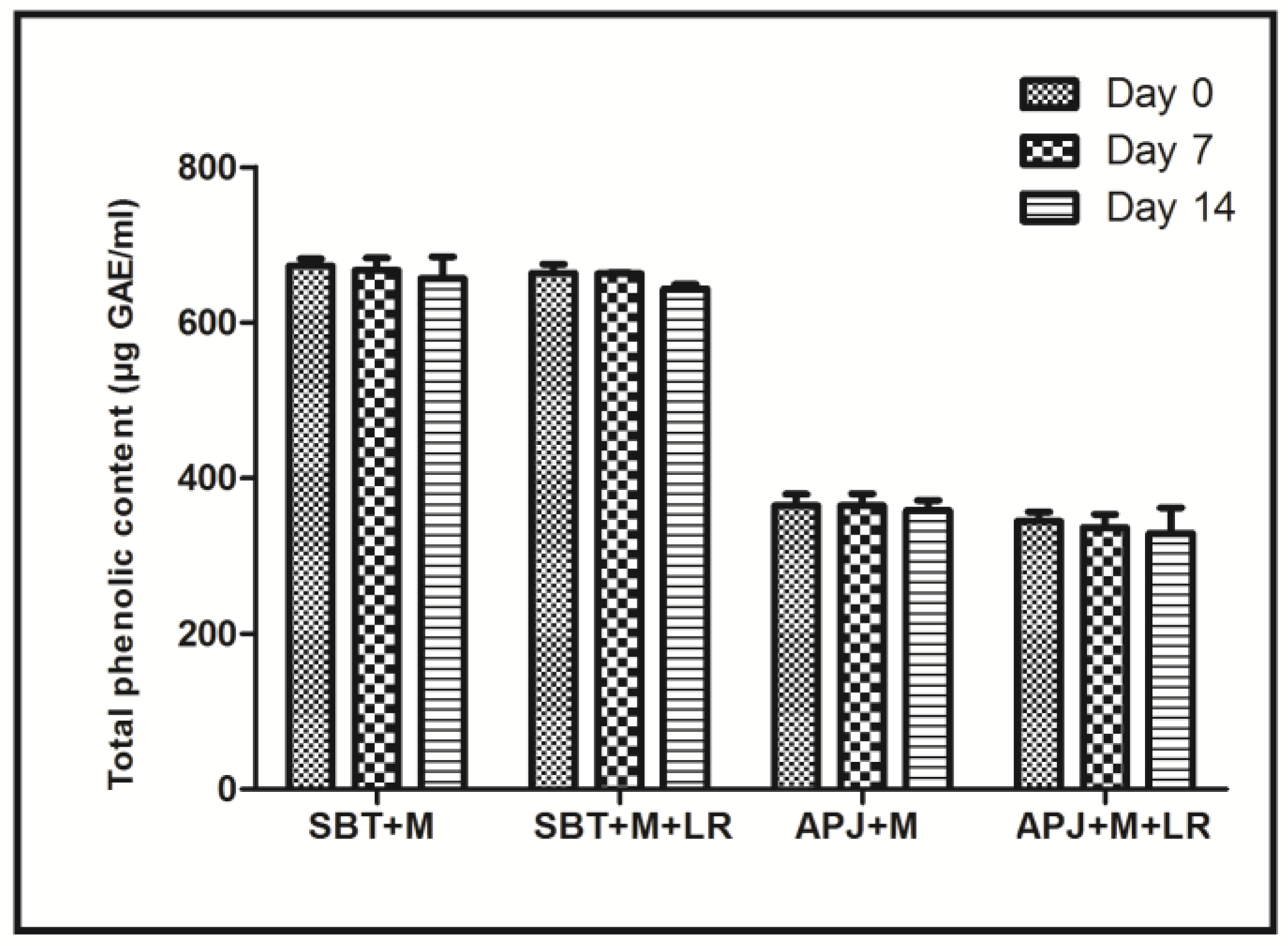
2.4. Total Phenolic Content (TPC)
2.5. Statistical Analysis
3. Results and Discussion
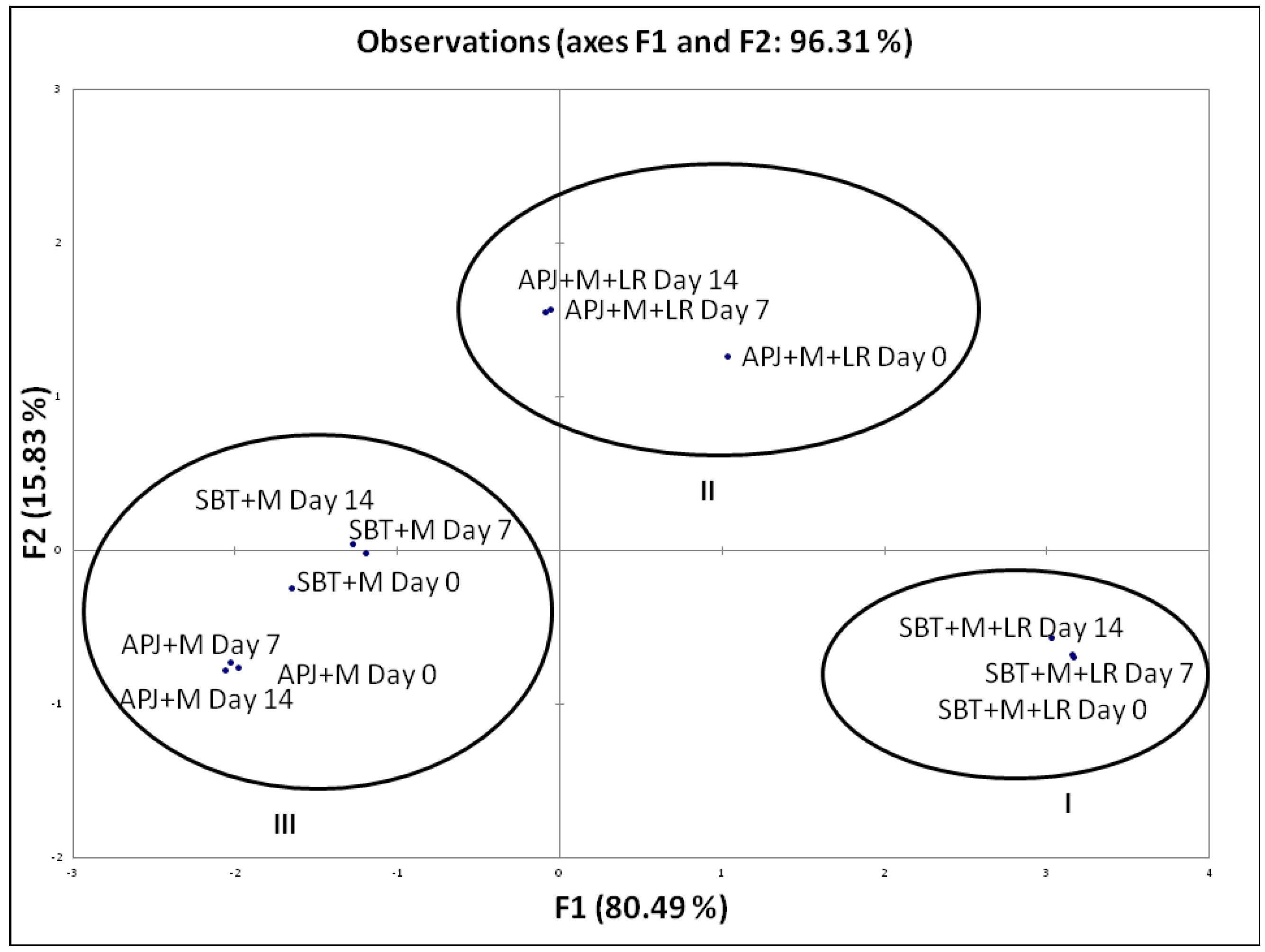
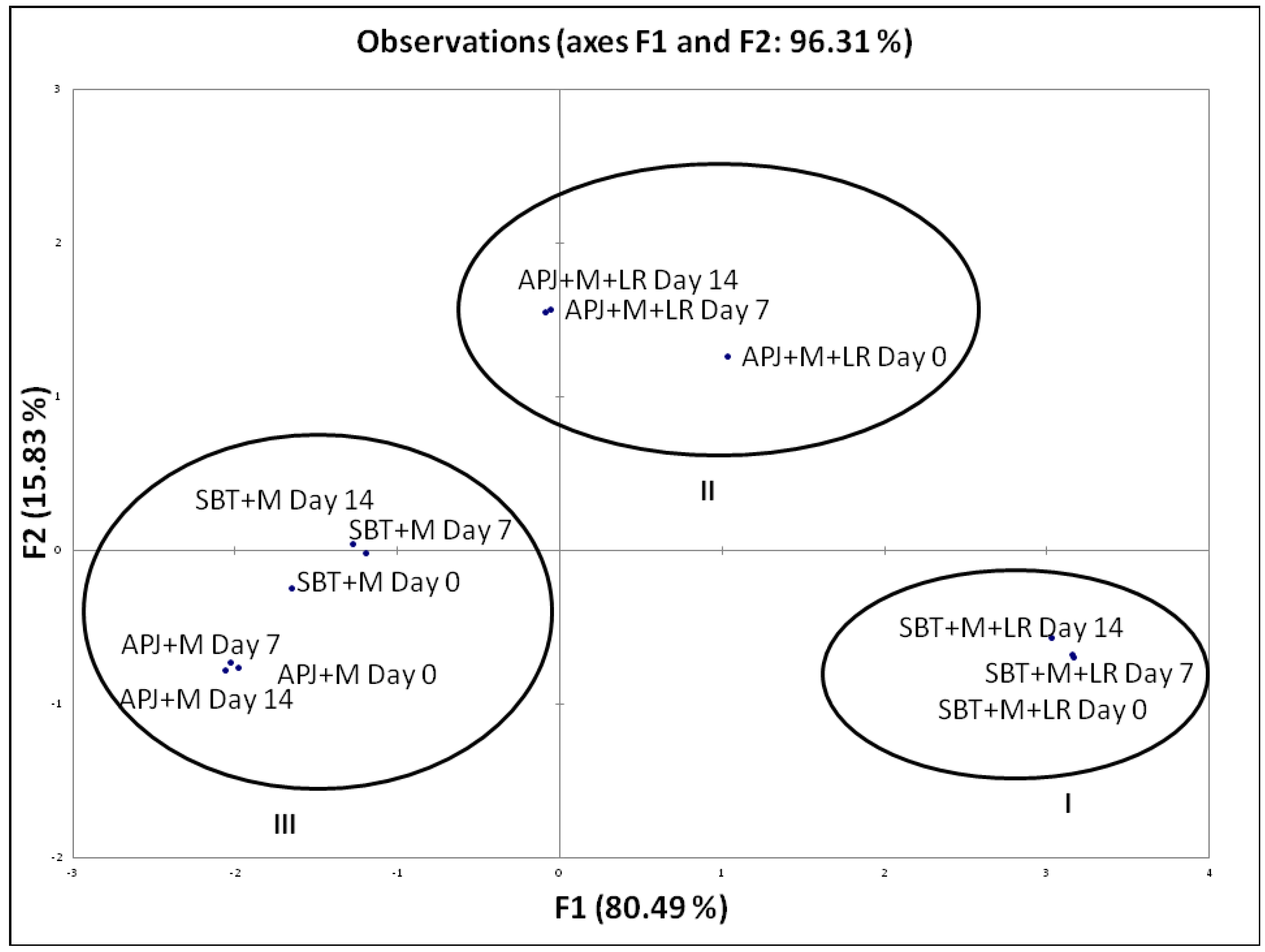
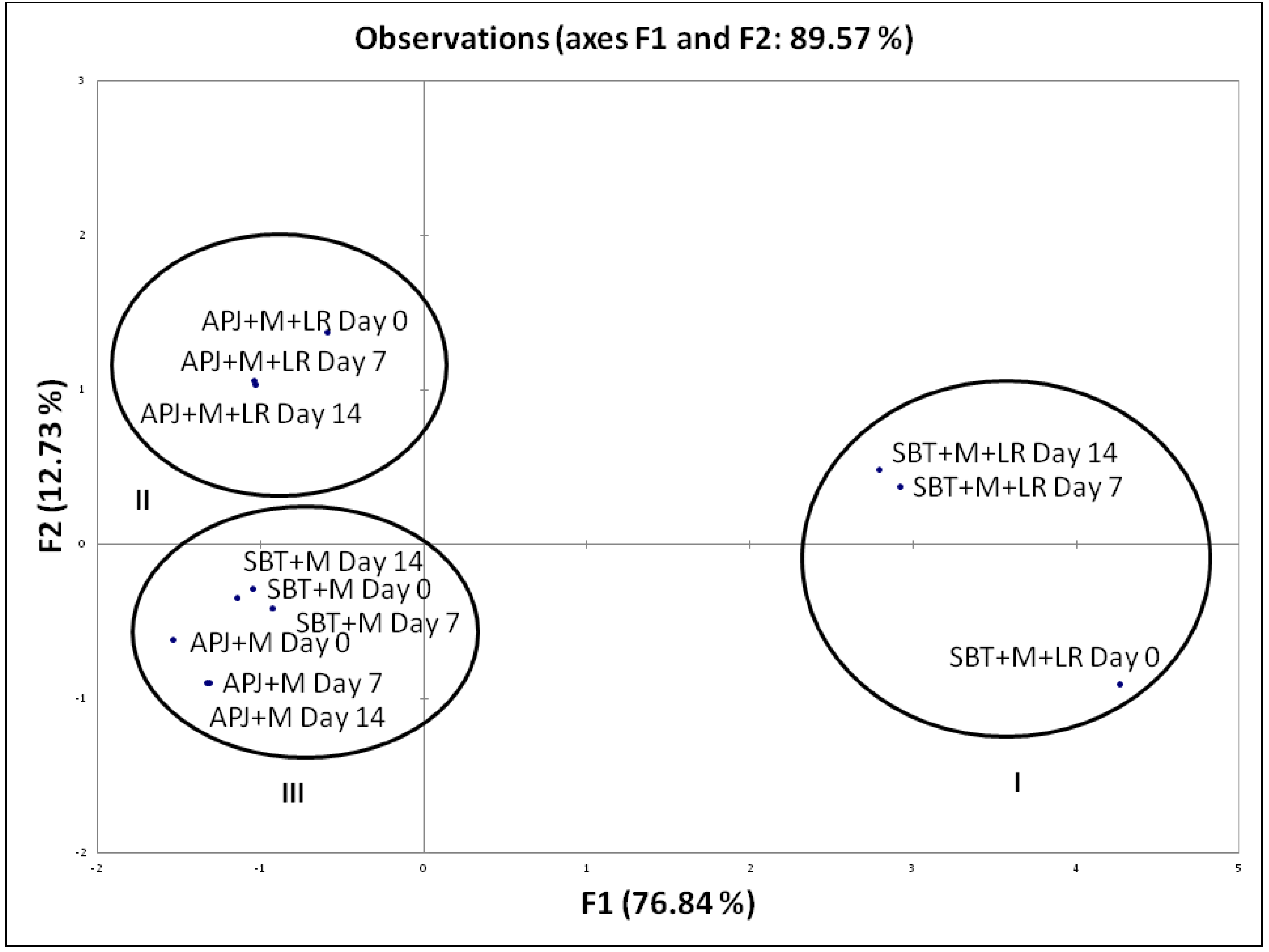
Percentage Pathogen Clearance and Principal Component Analysis (PCA)
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef]
- GBD. Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 acute and chronic diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar]
- Kotloff, K.L. The burden and etiology of diarrheal illness in developing countries. Pediatr. Clin. N. Am. 2017, 64, 799–814. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, S.K. Shigellosis. J. Microbiol. 2005, 43, 133–143. [Google Scholar] [PubMed]
- Sur, D.; Ramamurthy, T.; Deen, J.; Bhattacharya, S.K. Shigellosis: Challenges & management issues. Indian J. Med. Res. 2004, 20, 454–462. [Google Scholar]
- Greenhill, C. Diarrhoea: Tackling the problem of moderate-to-severe diarrhoea in developing countries. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 384–385. [Google Scholar] [CrossRef] [PubMed]
- Rabaa, M.A.; Thanh, D.P.; De Lappe, N.; Cormican, M.; Valcanis, M.; Howden, B.P.; Wangchuk, S.; Bodhidatta, L.; Mason, C.J.; Nguyen, T.N.; et al. South Asia as a reservoir for the global spread of ciprofloxacin-resistant Shigella sonnei: A cross-sectional study. PLoS Med. 2016, 13, e1002055. [Google Scholar] [CrossRef]
- Livio, S.; Strockbine, N.A.; Panchalingam, S.; Tennant, S.M.; Barry, E.M.; Marohn, M.E.; Antonio, M.; Hossain, A.; Mandomando, I.; Ochieng, J.B.; et al. Shigella isolates from the global enteric multicenter study inform vaccine development. Clin. Infect. Dis. 2014, 59, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Youn, S.K.; Lee, S.; Choi, Y.H. Epidemiological Characteristics of Imported Shigellosis in Korea, 2010–2011. Osong Public Health Res. Perspect. 2013, 4, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Kosek, M.; Yori, P.P.; Olortegui, M.P. Shigellosis update: Advancing antibiotic resistance, investment empowered vaccine development, and green bananas. Curr. Opin. Infect. Dis. 2017, 23, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology 2010, 156, 3216–3223. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.S.; Jalgaonkar, S.; Shahani, S.; Kulkarni, V.N. Probiotics: Current trends in the treatment of diarrhoea. Hong Kong Med. J. 2010, 16, 213–218. [Google Scholar] [PubMed]
- Hemarajata, P.; Versalovic, J. Effects of probiotics on gut microbiota: Mechanisms of intestinal immunomodulation and neuromodulation. Therap. Adv. Gastroenterol. 2013, 6, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Tejero-Sariñena, S.; Barlow, J.; Costabile, A.; Gibson, G.R.; Rowland, I. In vitro evaluation of the antimicrobial activity of a range of probiotics against pathogens: Evidence for the effects of organic acids. Anaerobe 2012, 18, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Davoodabadi, A.; Dallal, M.M.; Foroushani, A.R.; Douraghi, M.; Harati, F.A. Antibacterial activity of Lactobacillus spp. isolated from the feces of healthy infants against enteropathogenic bacteria. Anaerobe 2015, 34, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, L.; Du, M.; Yi, H.; Guo, C.; Tuo, Y.; Han, X.; Li, J.; Zhang, L.; Yang, L. Antimicrobial activity against Shigella sonnei and probiotic properties of wild lactobacilli from fermented food. Microbiol. Res. 2011, 167, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Kakisu, E.; Bolla, P.; Abraham, A.G.; De Urraza, P.; de Antoni, G.L. Lactobacillus plantarum isolated from kefir: Protection of cultured Hep-2 cells against Shigella invasion. Int. Dairy J. 2013, 33, 22–26. [Google Scholar] [CrossRef]
- Carlson, J.; Slavin, J. Health benefits of fibre, prebiotics and probiotics: A review of intestinal health and related health claims. Qual. Assur. Saf. Crop 2016, 8, 539–554. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Funct. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Galgano, F.; Condelli, N.; Caruso, M.C.; Colangelo, M.A.; Favati, F. Probiotics and prebiotics in fruits and vegetables: Technological and sensory aspects. In Beneficial Microbes in Fermented and Functional Foods; CRC Press-Taylor & Francis Group: Abingdon, UK, 2014; pp. 189–206. [Google Scholar]
- Güler-Akın, M.B.; Ferliarslan, I.; Akın, M.S. Apricot probiotic drinking yoghurt supplied with inulin and oat fiber. Adv. Microbiol. 2016, 6, 999–1009. [Google Scholar] [CrossRef]
- Homayoni Rad, A.; Vaghef Mehrabany, E.; Alipoor, B.; Vaghef Mehrabany, L. The comparison of food and supplement as probiotic delivery vehicles. Crit. Rev. Food Sci. Nutr. 2016, 56, 896–909. [Google Scholar] [CrossRef] [PubMed]
- Alwis, A.D.P.S.; Perera, O.D.A.N.; Weerahewa, H.L.D. Development of a Novel Carrot-based Synbiotic Beverage using Lactobacillus casei 431®. J. Agric. Sci. 2016, 11, 178–185. [Google Scholar] [CrossRef]
- Di Cagno, R.; Coda, R.; De Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.N.; Messaoud, G.B.; Desobry, S.; Costa, J.M.C.; Rodrigues, S. Effect of drying technique and feed flow rate on bacterial survival and physicochemical properties of a non-dairy fermented probiotic juice powder. J. Food Eng. 2016, 189, 45–54. [Google Scholar] [CrossRef]
- Dharmasena, M.; Barron, F.; Fraser, A.; Jiang, X. Refrigerated Shelf Life of a Coconut Water-Oatmeal Mix and the Viability of Lactobacillus Plantarum Lp 115-400B. Foods 2015, 4, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of fermentation with different lactic acid bacteria and in vitro digestion on the biotransformation of phenolic compounds in fermented pomegranate juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, R.; Ibrahim, S.A. Impact of plant derivatives on the growth of foodborne pathogens and the functionality of probiotics. Appl. Microbiol. Biotechnol. 2012, 95, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Wishon, L.M.; Song, D.; Ibrahim, S. Effect of metals on growth and functionality of Lactobacillus and Bifidobacteria. Milchwissenschaft 2010, 65, 369–372. [Google Scholar]
- Sireswar, S.; Dey, G.; Sreesoundarya, T.K.; Sarkar, D. Design of probiotic-fortified food matrices influence their antipathogenic potential. Food Biosci. 2017, 20, 28–35. [Google Scholar] [CrossRef]
- Sireswar, S.; Dey, G.; Dey, K.; Kundu, A. Evaluation of Probiotic L. rhamnosus GG as a Protective Culture in Sea Buckthorn-Based Beverage. Beverages 2017, 3, 48. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Sánchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure–function relationships of the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 11, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Gunenc, A.; Khoury, C.; Legault, C.; Mirrashed, H.; Rijke, J.; Hosseinian, F. Seabuckthorn as a novel prebiotic source improves probiotic viability in yogurt. LWT-Food Sci. Technol. 2016, 66, 490–495. [Google Scholar] [CrossRef]
- Terpou, A.; Gialleli, A.I.; Bosnea, L.; Kanellaki, M.; Koutinas, A.A.; Castro, G.R. Novel cheese production by incorporation of sea buckthorn berries (Hippophae rhamnoides L.) supported probiotic cells. LWT-Food Sci. Technol. 2017, 79, 616–624. [Google Scholar] [CrossRef]
- Arora, R.; Mundra, S.; Yadav, A.; Srivastava, R.B.; Stobdan, T. Antimicrobial activity of seed, pomace and leaf extracts of sea buckthorn (Hippophae rhamnoides L.) against foodborne and food spoilage pathogens. Afr. J. Biotechnol. 2012, 11, 10424–10430. [Google Scholar] [CrossRef]
- Van Opstal, I.; Bagamboula, C.F.; Theys, T.; Vanmuysen, S.C.M.; Michiels, C.W. Inactivation of Escherichia coli and Shigella in acidic fruit and vegetable juices by peroxidase systems. J. Appl Microbiol. 2006, 101, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Vinderola, G.; Binetti, A.; Burns, P.; Reinheimer, J. Cell viability and functionality of probiotic bacteria in dairy products. Front. Microbiol. 2011, 2, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Madureira, A.R.; Amorim, M.; Gomes, A.M.; Pintado, M.E.; Malcata, F.X. Protective effect of whey cheese matrix on probiotic strains exposed to simulated gastrointestinal conditions. Food Res. Int. 2011, 44, 465–470. [Google Scholar] [CrossRef]
- In, Y.W.; Kim, J.J.; Kim, H.J.; Oh, S.W. Antimicrobial activities of acetic acid, citric acid and lactic acid against Shigella species. J. Food Saf. 2013, 33, 79–85. [Google Scholar] [CrossRef]
- Bagamboula, C.F.; Uyttendaele, M.; Debevere, J. Acid tolerance of Shigella sonnei and Shigella flexneri. J. Appl. Microbiol. 2002, 93, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wang, L.; Wang, Y.; Li, P.; Zhu, J.; Qiu, S.; Hao, R.; Wu, Z.; Li, W.; Song, H. hfq regulates acid tolerance and virulence by responding to acid stress in Shigella flexneri. Res. Microbiol. 2015, 166, 166–476. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Wu, V.C.; White, J.; Tadepalli, S.; Andre, E.E. The antimicrobial properties of the lowbush blueberry (Vaccinium angustifolium) fractional components against foodborne pathogens and the conservation of probiotic Lactobacillus rhamnosus. Food Microbiol. 2012, 30, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Rodrıguez, E.; Calzada, J.; Arqués, J.L.; Rodrıguez, J.M.; Nunez, M.; Medina, M. Antimicrobial activity of pediocin-producing Lactococcus lactis on Listeria monocytogenes, Staphylococcus aureus and Escherichia coli O157: H7 in cheese. Int. Dairy J. 2005, 15, 51–57. [Google Scholar] [CrossRef]
- McAuliffe, O.; Hill, C.; Ross, R.P. Inhibition of Listeria monocytogenes in cottage cheese manufactured with a lacticin 3147-producing starter culture. J. Appl. Microbiol. 1999, 86, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Arqués, J.L.; Rodríguez, E.; Gaya, P.; Medina, M.; Guamis, B.; Nunez, M. Inactivation of Staphylococcus aureus in raw milk cheese by combinations of high-pressure treatments and bacteriocin-producing lactic acid bacteria. J. Appl. Microbiol. 2005, 98, 254–260. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percentage Pathogen Clearance (%) | 1 h | 2 h | 3 h | 4 h | 8 h |
|---|---|---|---|---|---|
| SBT + M (Day 0) | 6.78 | 9.88 | 17.26 | 37.82 | 76.1 |
| SBT + M (Day 7) | 5.56 | 8.19 | 27.92 | 48.19 | 69.37 |
| SBT + M (Day 14) | 4.48 | 9.16 | 11.28 | 28.28 | 69.29 |
| SBT + M + LR (Day 0) | 99.99 | 99.99 | 99.99 | 99.99 | 99.99 |
| SBT + M + LR (Day 7) | 99.26 | 99.62 | 99.99 | 99.99 | 99.99 |
| SBT + M + LR (Day 14) | 89.28 | 99.08 | 99.99 | 99.99 | 99.99 |
| APJ + M (Day 0) | 5.43 | 8.12 | 10.75 | 20.18 | 57.19 |
| APJ + M (Day 7) | 4.49 | 8 | 11.18 | 26.27 | 52.16 |
| APJ + M (Day 14) | 4.41 | 7.72 | 16.9 | 22.27 | 55.34 |
| APJ + M + LR (Day 0) | 10.75 | 15.28 | 91.68 | 99.99 | 99.99 |
| APJ + M + LR (Day 7) | 9.26 | 10.26 | 13.43 | 92.89 | 99.99 |
| APJ + M + LR (Day 14) | 8.27 | 9.17 | 15.29 | 90.48 | 99.99 |
| Percentage Pathogen Clearance (%) | 1 h | 2 h | 3 h | 4 h | 8 h |
|---|---|---|---|---|---|
| SBT + M (Day 0) | 4.7 | 5.67 | 10.18 | 22.19 | 56.1 |
| SBT + M (Day 7) | 4.56 | 6.18 | 17.98 | 27.63 | 54.29 |
| SBT + M (Day 14) | 4.3 | 5.56 | 6.11 | 28.82 | 55.28 |
| SBT + M + LR (Day 0) | 8.4 | 90.58 | 99.73 | 99.87 | 99.99 |
| SBT + M + LR (Day 7) | 8.38 | 17.19 | 90.3 | 99.36 | 99.99 |
| SBT + M + LR (Day 14) | 8.29 | 11.19 | 90.26 | 99.17 | 99.99 |
| APJ + M (Day 0) | 4.67 | 4.89 | 5.08 | 5.78 | 42.98 |
| APJ + M (Day 7) | 4.3 | 6.19 | 8.19 | 28.98 | 40.17 |
| APJ + M (Day 14) | 4.29 | 5.56 | 6.98 | 30.27 | 40.1 |
| APJ + M + LR (Day 0) | 5.09 | 5.18 | 5.56 | 6.87 | 99.99 |
| APJ +M + LR (Day 7) | 4.08 | 5.09 | 5.42 | 5.95 | 90.72 |
| APJ + M + LR (Day 14) | 4 | 5.18 | 5.87 | 6.9 | 90.36 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sireswar, S.; Montet, D.; Dey, G. Principal Component Analysis for Clustering Probiotic-Fortified Beverage Matrices Efficient in Elimination of Shigella sp. Fermentation 2018, 4, 34. https://doi.org/10.3390/fermentation4020034
Sireswar S, Montet D, Dey G. Principal Component Analysis for Clustering Probiotic-Fortified Beverage Matrices Efficient in Elimination of Shigella sp. Fermentation. 2018; 4(2):34. https://doi.org/10.3390/fermentation4020034
Chicago/Turabian StyleSireswar, Srijita, Didier Montet, and Gargi Dey. 2018. "Principal Component Analysis for Clustering Probiotic-Fortified Beverage Matrices Efficient in Elimination of Shigella sp." Fermentation 4, no. 2: 34. https://doi.org/10.3390/fermentation4020034