3.1. Biomass Growth
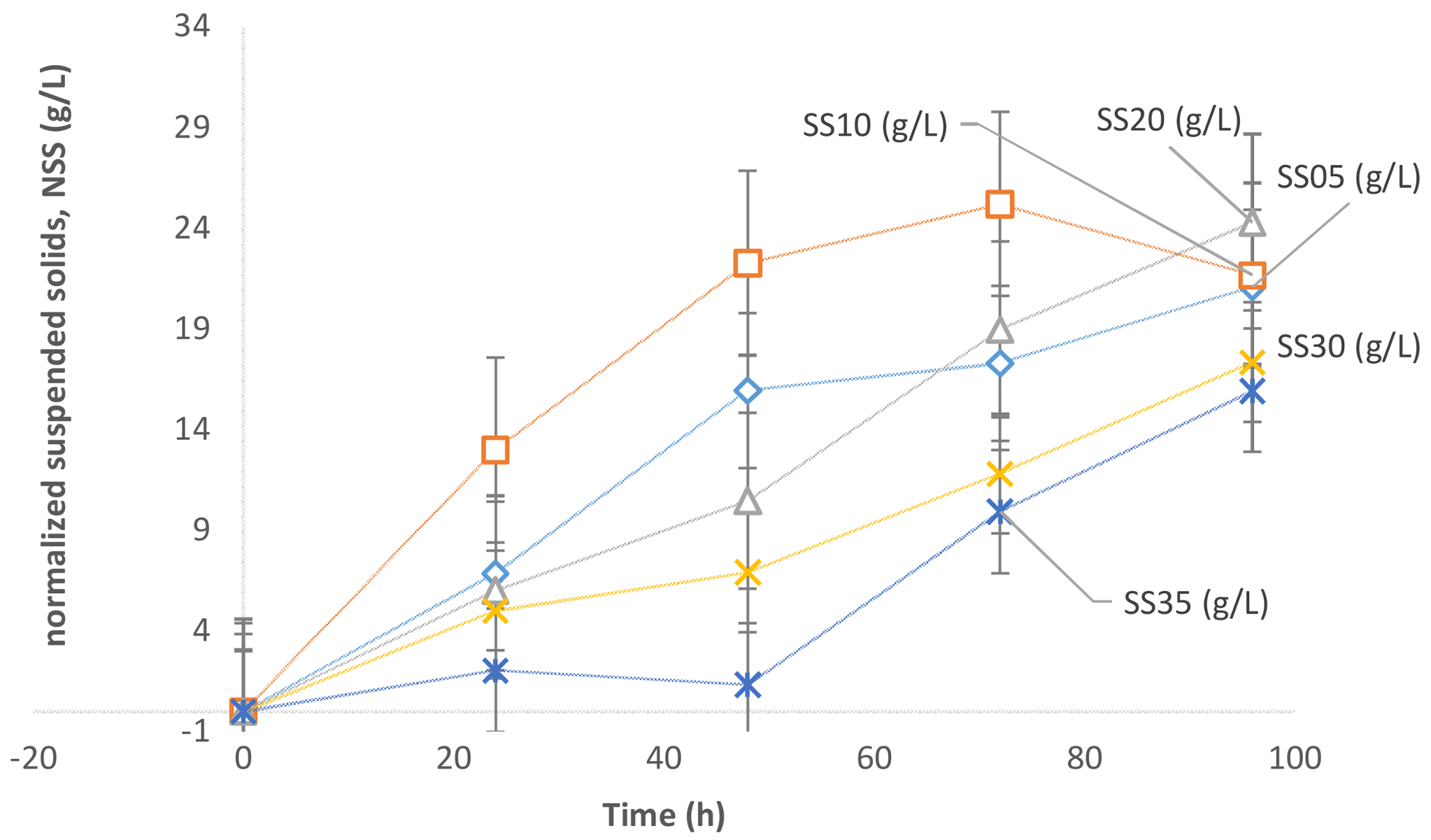
The normalized suspended solids (NSS) profiles in the case of all of the flask fermentations with different sludge solid concentrations are presented in
Figure 1. In this context, normalized suspended solids (NSS) is the net increase in solid concentration obtained by subtracting the 0th-hour suspended solid concentration (after basic thermal sterilization) from the suspended solid concentration at any time, t.
The solid sludge concentrations depicted in
Figure 1 (NSS) are solid concentrations, including intracellular lipids. In
Figure 1, the suspended sludge concentration has a significant impact on the microbial growth of the yeast
Yarrowia lipolytica. The maximum biomass was generated with a sludge concentration of 20 g/L (SS10). The net biomass generated was 24.29 g/L. The biomass, in this case, increased from an initial SS of 10.57 g/L (after sterilization) to a final biomass concentration of 34.86 g/L (
Table 3). With further increase in sludge concentration beyond 20 g/L the net increase in biomass concentration decreases. Subsequently, the net biomass generated was in the order of (case of) 20 g/L > 10 g/L > 5 g/L > 30 g/L and 35 g/L with a net biomass value of 24.29, 21.66, 21.08, 17.34, and 15.93, respectively. The results indicate a clear inhibition to the microbial growth of the yeast
Y. lipolytica at concentrations higher than 20 g/L.
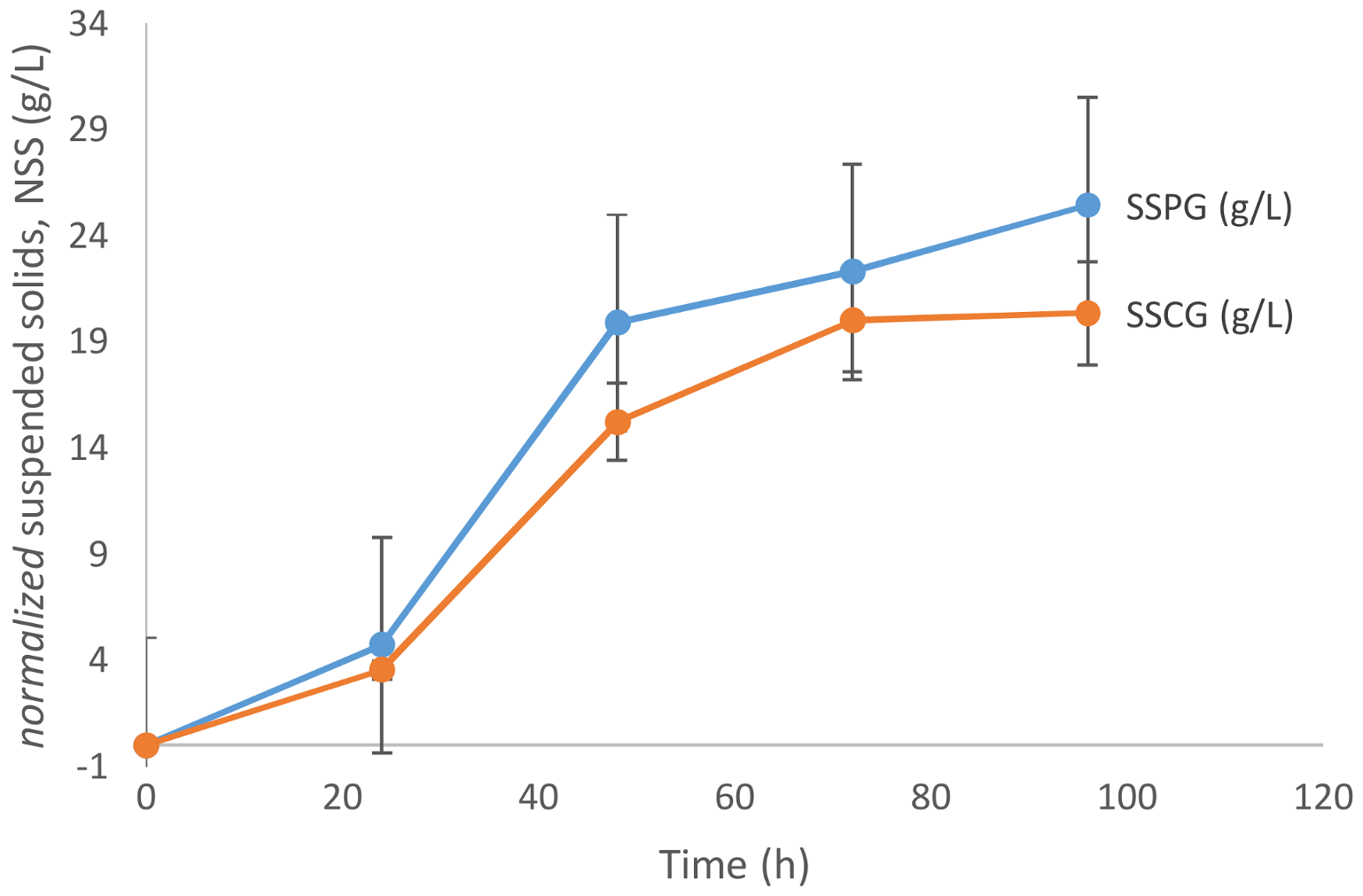
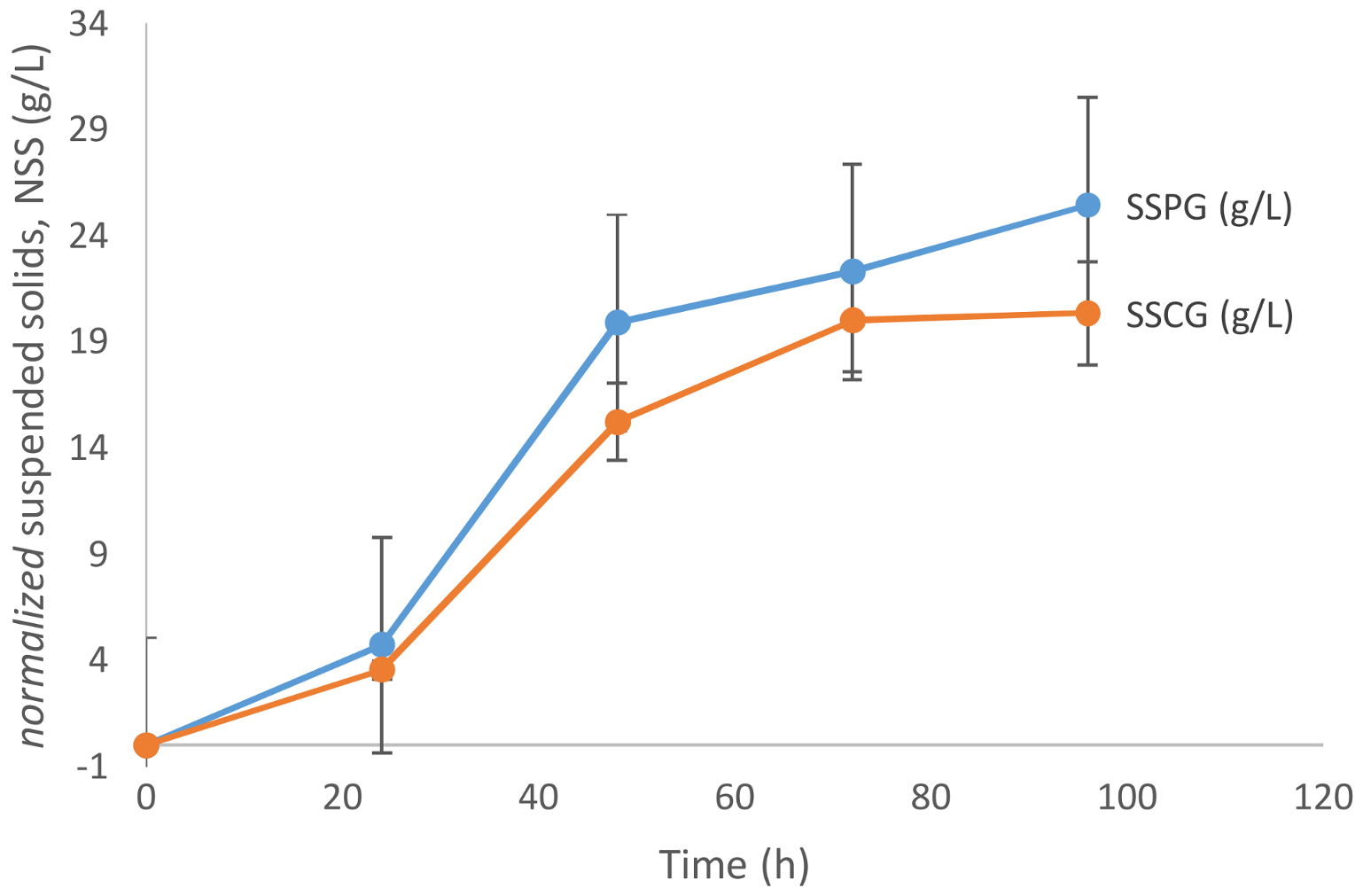
As is evident from
Figure 2, the biomass growth in the presence of pure glycerol is relatively higher than that in crude glycerol. The maximum biomass concentration achieved in the case of crude glycerol is 20.36 g/L after 96 h of fermentation, while in the case of pure glycerol the maximum biomass concentration observed was 25.45 g/L. Papanikolaou and Aggelis [
13] in their study found that crude glycerol (65%
w/
w glycerol content) poses some inhibition to the microbial growth of
Y. lipolytica due to its matrix composition. Despite the probable matrix inhibition, the growth with crude and pure glycerol was comparable. This can be attributed to the fact that the strains of
Y. lipolytica have higher tolerance towards the major impurities found in crude glycerol-like methanol, free fatty acids (FFA), basic salts (NaOH, KOH, and methoxides), and soap.
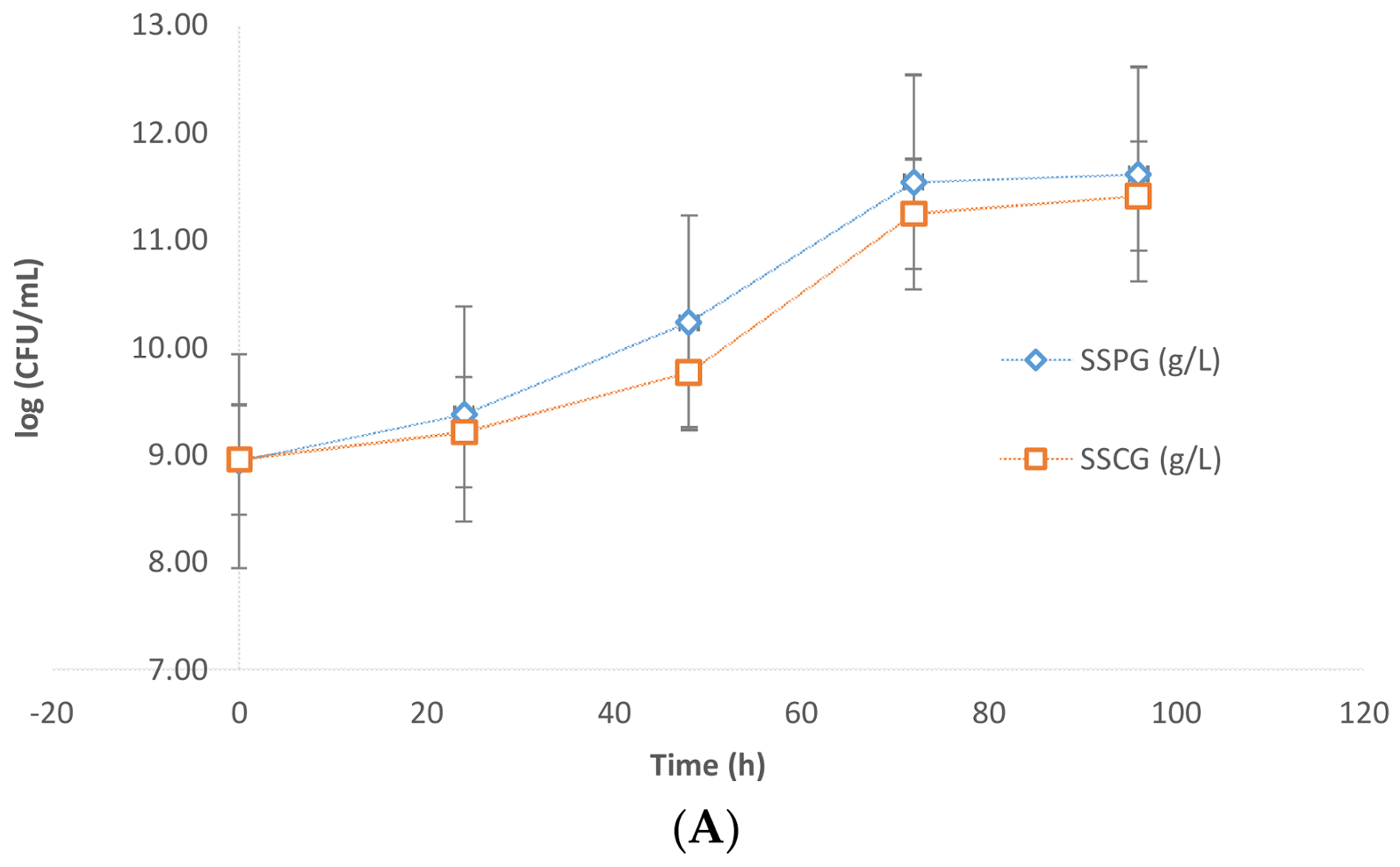
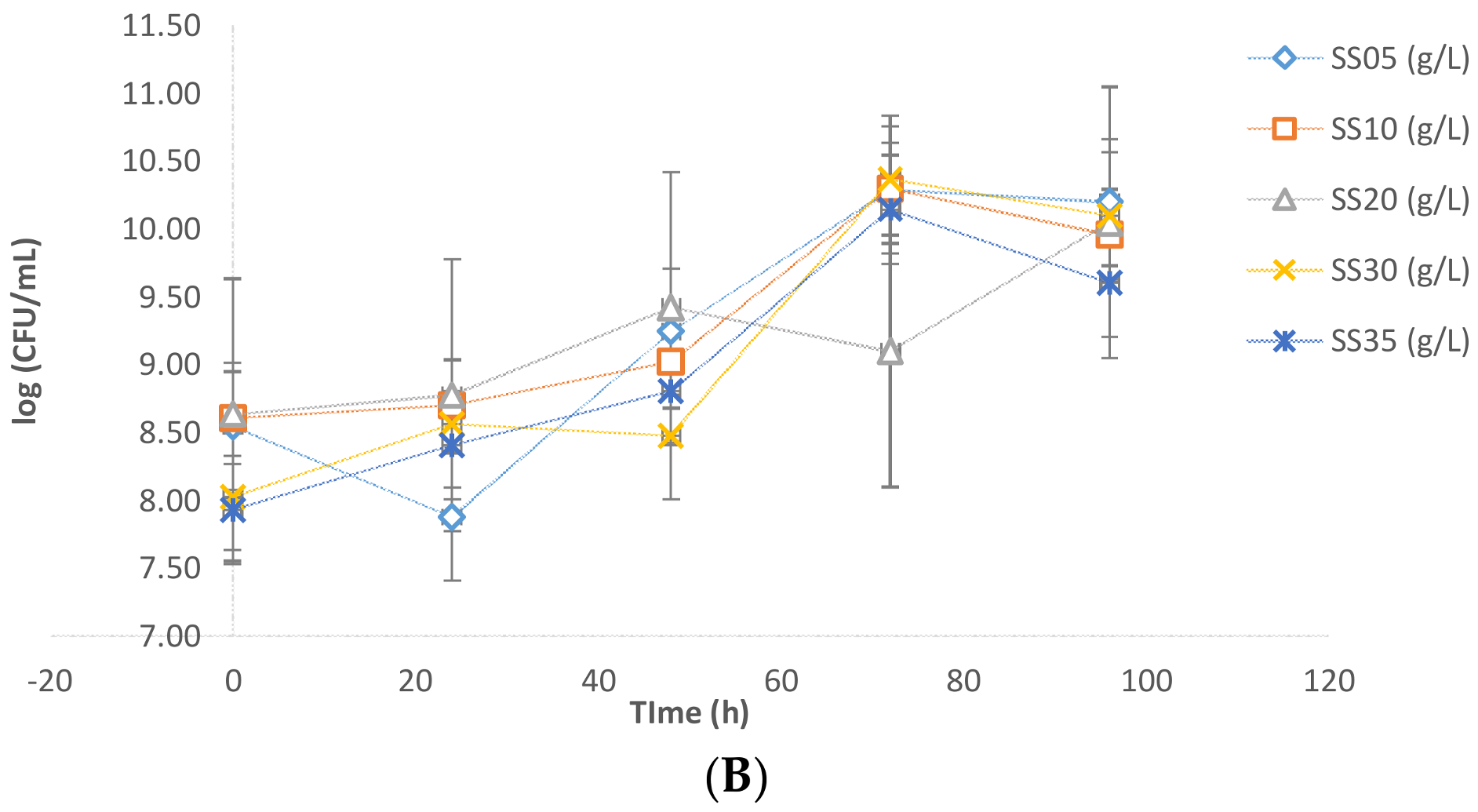
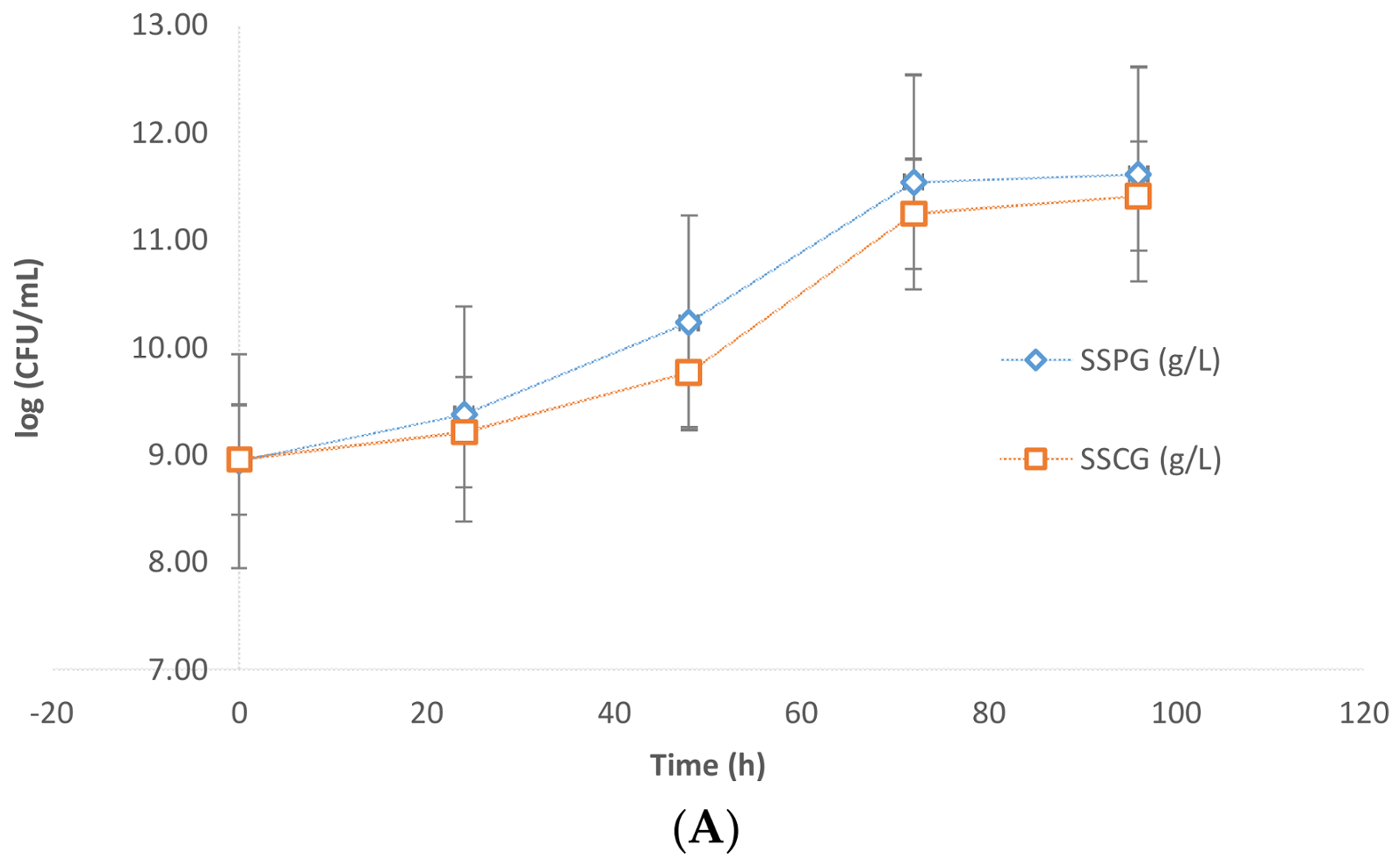
Evidently, the presence of sludge imposes some inhibition on the microbial growth of the yeast. The results were further confirmed by CFU analysis data. In
Figure 3A,B, it is apparent that the values of Log (CFU/mL) in the presence of sludge were in the order of 10–10.4 while in the case of flask fermentations without sludge these values were in the order of 11.5. Further slight inhibition was observed due to crude glycerol. The CFU/mL in the case of pure glycerol was found to be higher than that of crude glycerol (
Figure 3A).
Statistical Analysis of the net biomass generation using single factor ANOVA was performed using the MS-Excel 2013 data analysis tool. The duplicates data for the net biomass generation was used for the statistical analysis. The results are summarized in
Table 4. According to the test results, the F ratio (of variances) value was 13.899, which was higher than the F critical value of 5.19217 as recommended by the Fisher’s F test table for statistical significance for 95% confidence for the F(4,5) degree of freedom data set. Further, the probability value (
p value) of the test was 0.006 < 0.05, less than the set
p value for this test. Therefore, the results indicate that the null hypothesis can be easily rejected to conclude that the variation in sludge suspended solid concentrations have a significant effect on the biomass growth of the microbe.
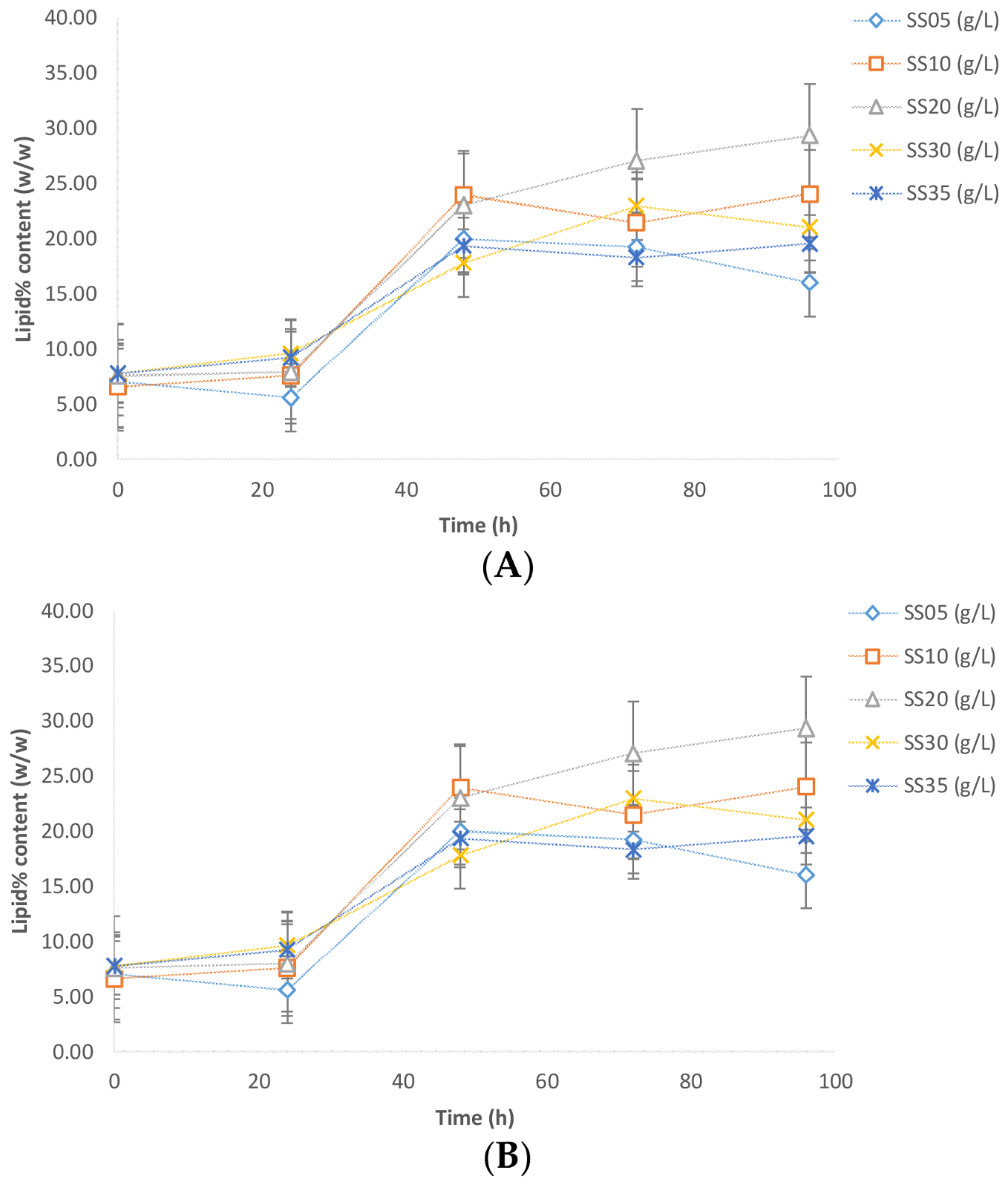
3.2. Lipid Production
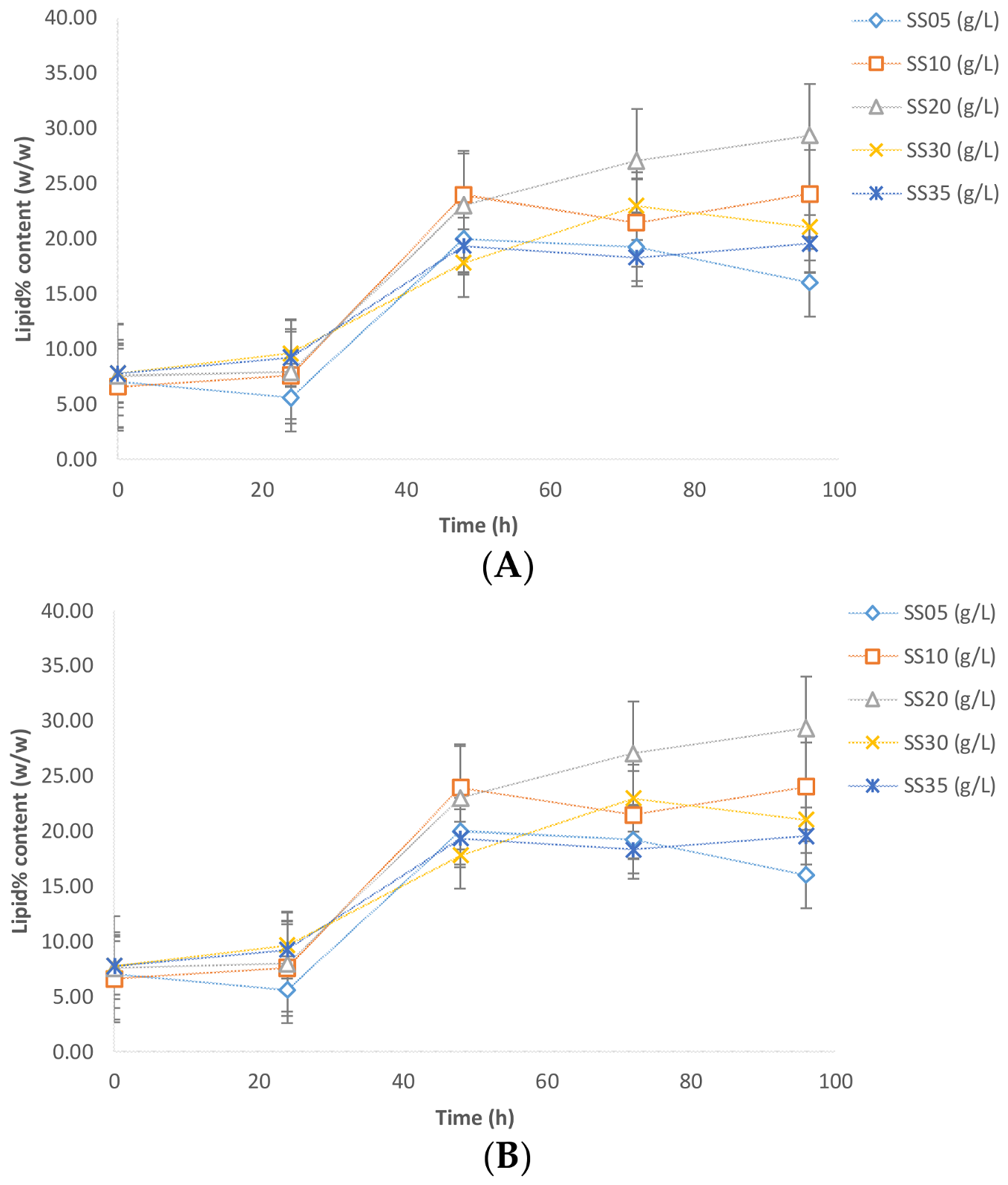
The maximum lipid content (29.35%
w/
w) was observed with the sludge solid concentration of 20 g/L, whereas the lowest lipid content was observed with the 5 g/L sludge solid concentration (
Figure 4). The lipid content in the cases of the 10, 30, and 35 g/L sludge concentrations were 20–24%
w/
w. A possible account for such variation is the (carbon and nitrogen) nutrients furnished by the sludge. If the nitrogen concentration is too low, then the lipid accumulation process is negatively affected by virtue of the fact that a minimal concentration of nitrogen is required for basal cell metabolism and accumulation activity. On the other hand, if an excess of nitrogen is available (low C:N ratio), the cell favors growth metabolism as against the accumulation of carbon as lipid reserves [
3]. This suggests that an optimum level of nitrogen is required for the microbes to perform lipid accumulation. The presence of different concentrations of sludge solids furnishes different concentrations of the nutrients (carbon and nitrogen), resulting in different sub-optimal C:N ratios for lipid accumulation.
In experiments performed without sludge to compare pure glycerol and crude glycerol, the lipid contents were 35% and 28% for crude glycerol and pure glycerol, respectively (
Figure 4B). This result is in agreement with the results obtained by previous studies [
3,
8,
13,
14]. Similar to biomass, inhibition of lipid accumulation was also observed by the presence of sludge (
Figure 4A,B). Maximum lipid content with sludge (in case of 20 g/L) was 29.35% (
w/
w), while without sludge the maximum lipid content was 37% (
w/
w). Further, crude glycerol is approvingly a favorable source for lipid accumulation using
Y. lipolytica. A comparison between lipid content and the yield of previous studies and this study has been presented in
Table 5.
3.3. Glycerol Consumption
Many researchers have studied the effect of crude glycerol composition and found that crude glycerol obtained from the biodiesel industry is a very suitable carbon source for lipid production [
16]. In this study, crude glycerol obtained from the biodiesel industry via Bio-Liq, Saint-Marc-des-Carrières, QC, Canada was used. An initial C:N ratio of 100 (mol-C/mol-N) was maintained in all of the experiments as a C:N ratio of 75–100 has been reported to be optimum for lipid production [
3,
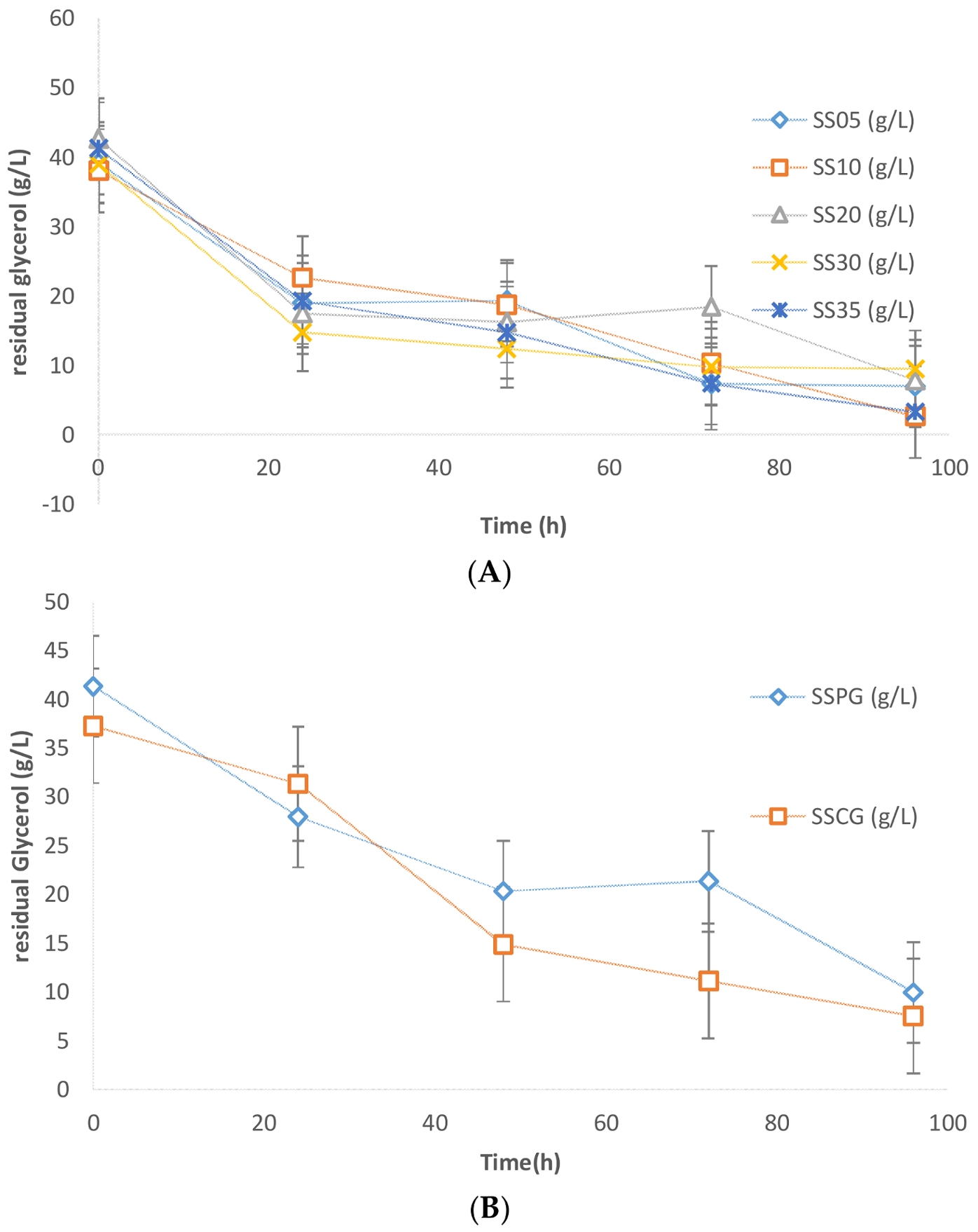
16]. The glycerol consumption profile is presented in
Figure 5, and it is observed that glycerol was rapidly consumed in the first 24 h of fermentation in all of the different sludge solid concentrations. After 96 h of fermentation, the residual glycerol concentration was found to be in the range of 3–9 g/L. Similar glycerol consumption profiles were observed in case of experiments performed without sludge solids in the media. The residual glycerol concentration in the case of zero sludge solids was also in the range of 7.5–9.9 g/L. As seen earlier in the SS and lipid data, the crude glycerol used in this study apparently poses insignificant inhibition due to the traces of impurities. Also, no significant difference in substrate consumption rates was observed between pure glycerol and crude glycerol (
Figure 5B). An untraceable amount of citric acid was produced in these experiments, which can be attributed to the composition of the crude glycerol matrix and evolving C:N ratio. These results conform with the findings of previous studies [
15].
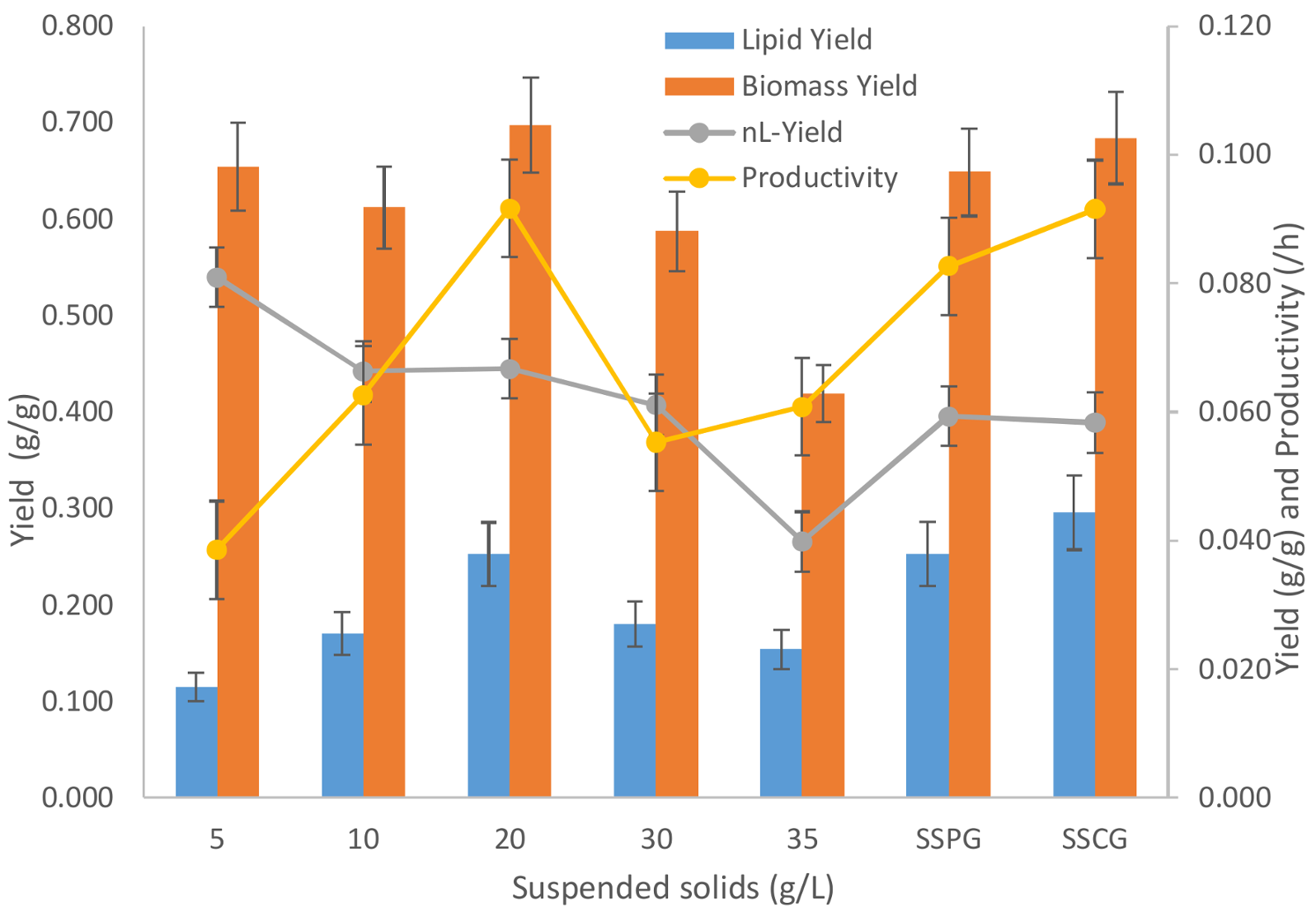
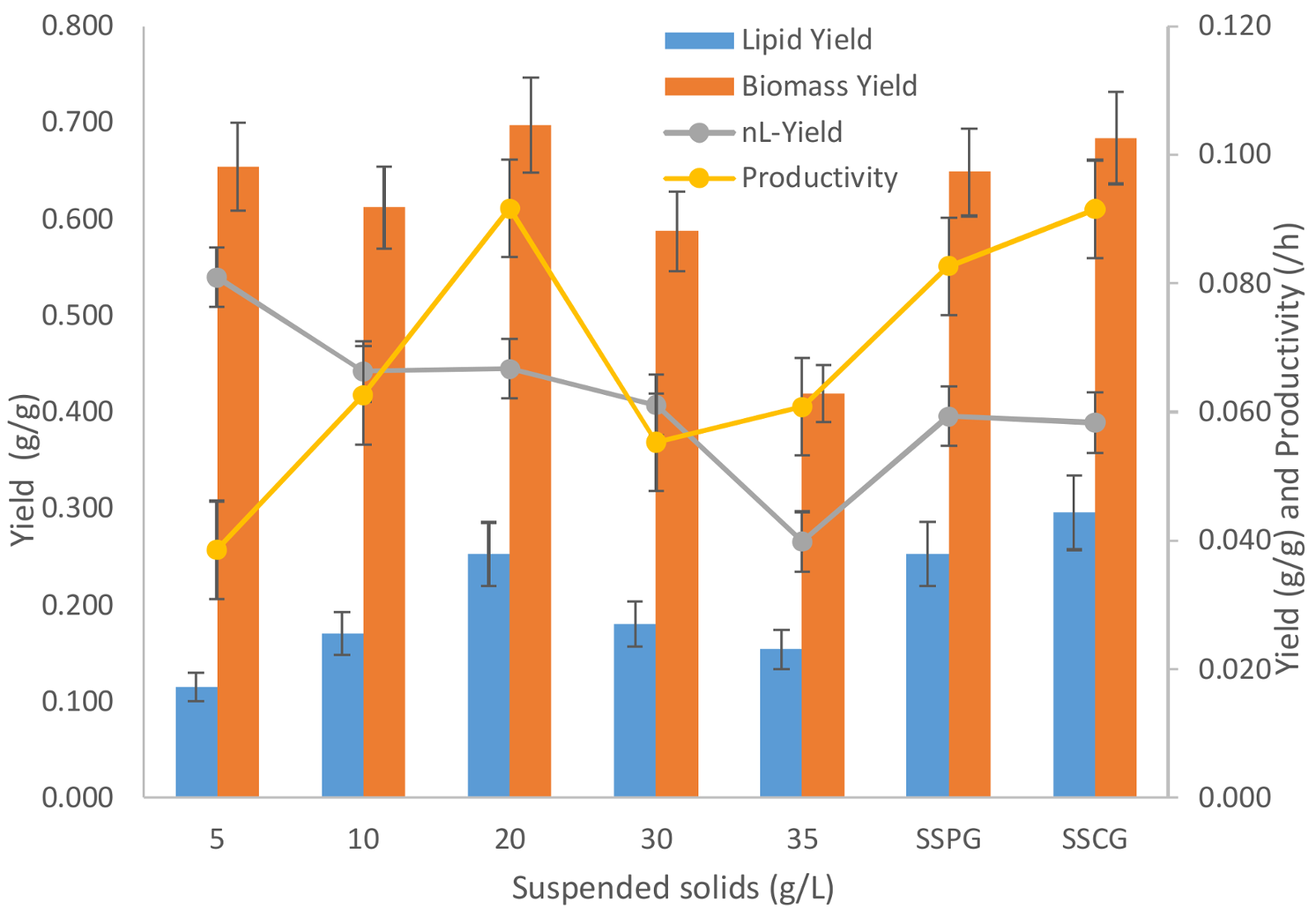
3.4. Yield and Productivity Analysis
The yield and productivity data for different suspended solid concentrations have been presented in
Figure 6. As is evident from the graphics, with an increase in sludge suspended solid concentration higher than 20 g/L in the media, the total biomass (including lipid) yield w.r.t glycerol decreases monotonously. This, as discussed earlier, is the clear indication of the growth inhibition caused by sludge solids.
The significant decrease in biomass yield can be accounted for with the hindrance caused by the sludge solids at higher concentrations. At higher concentrations of sludge solids, any inhibitory compound furnished by sludge solids will also be present in higher concentrations, which can affect the microbial growth. Further, a higher solid concentration can cause mass transfer limitations for nutrients and oxygen. A higher solid concentration can cause inhomogeneity resulting in anaerobic pockets in the flask fermentation, which can severely affect the microbial growth by creating hostile and stressful conditions [
17]. Under stressful conditions, cells invest a significant portion of their resources in the form of maintenance energy [
18]. Retarded microbial growth is beneficial for any kind of storage molecule accumulation as can be seen in this study that lipid yield increases when the sludge solid concentration is increased from 5 to 20 g/L (
Figure 6). Increasing the sludge solid concentration beyond 20 g/L decreases the lipid yield as the inhibitory effect of sludge solids scale the complete cellular metabolism causing neither growth (non-lipid biomass) nor accumulation of lipids.
The maximum lipid accumulation yield of 0.253 g-lipid/g-glycerol was found with 20 g/L of sludge solid concentration. The maximum productivity with 20 g/L sludge concentration was 0.092 g-lipid/L.h. The results of this study are comparable with previous studies done with this strain without sludge (
Table 5).
Comparing the lipid accumulation in the presence of the optimum sludge solid concentration (of 20 g/L) with experiments performed without sludge (SSCG, using crude glycerol), it is evident that while using municipal secondary sludge in the media the lipid yield and productivity are comparably invariant as compared to fermentation performed in absence of sludge (SSCG). When sludge was used, the biomass and n-Lipid biomass yields were 0.697 and 0.445 g/g, respectively. On the other hand, the biomass and n-Lipid biomass yields were 0.684 and 0.389 g/g, respectively, when sludge was not used (SSCG). In both the cases, the lipid content and glycerol usage were not compromised significantly. The lipid yield with sludge (20 g/L) was 0.253 g-lipid/g-glycerol, while the one without sludge (SSCG) was 0.295 g-lipid/g-glycerol. A comparison between the two cases has been presented in
Table 6.
Thus, it can be concluded that sludge at a higher concentration causes significant inhibition to microbial metabolism. If the optimum concentration of sludge solid is used for fermentation, then the lipid productivity and lipid content is not compromised as compared to its negative control (SSCG) where fermentation was performed without sludge. Use of crude glycerol (SSCG) as compared to pure glycerol (SSPG) is also beneficial in all aspects as it gives comparable lipid yield and higher lipid productivities. The productivity of crude glycerol (SSCG) is 0.092 whereas lipid yield for pure glycerol is 0.083 g-lipid/L.h. Therefore, crude glycerol is a preferable substrate which enhances the lipid production with a slight inhibition to microbial growth.
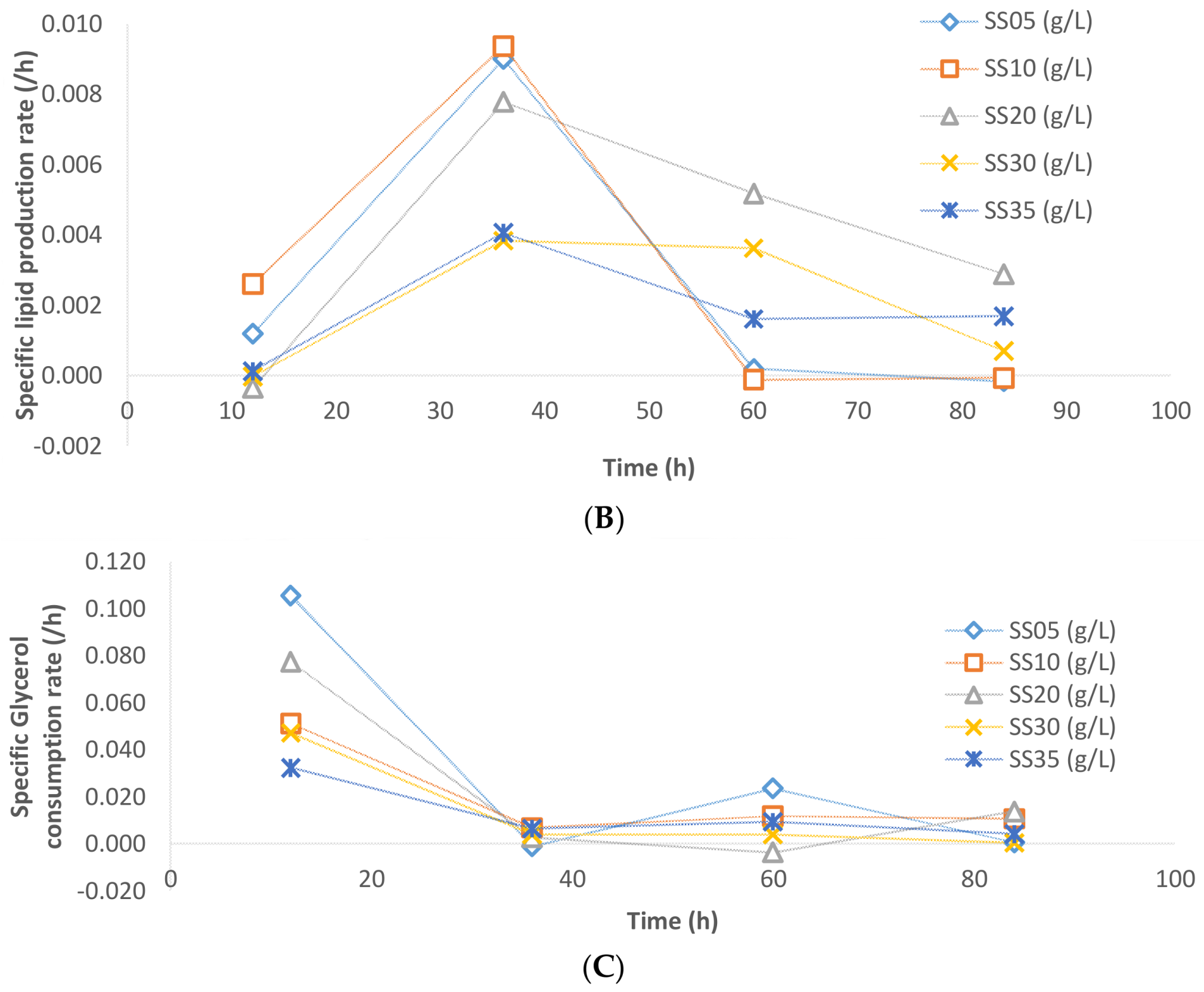
3.5. Specific Rates and Kinetics Analysis
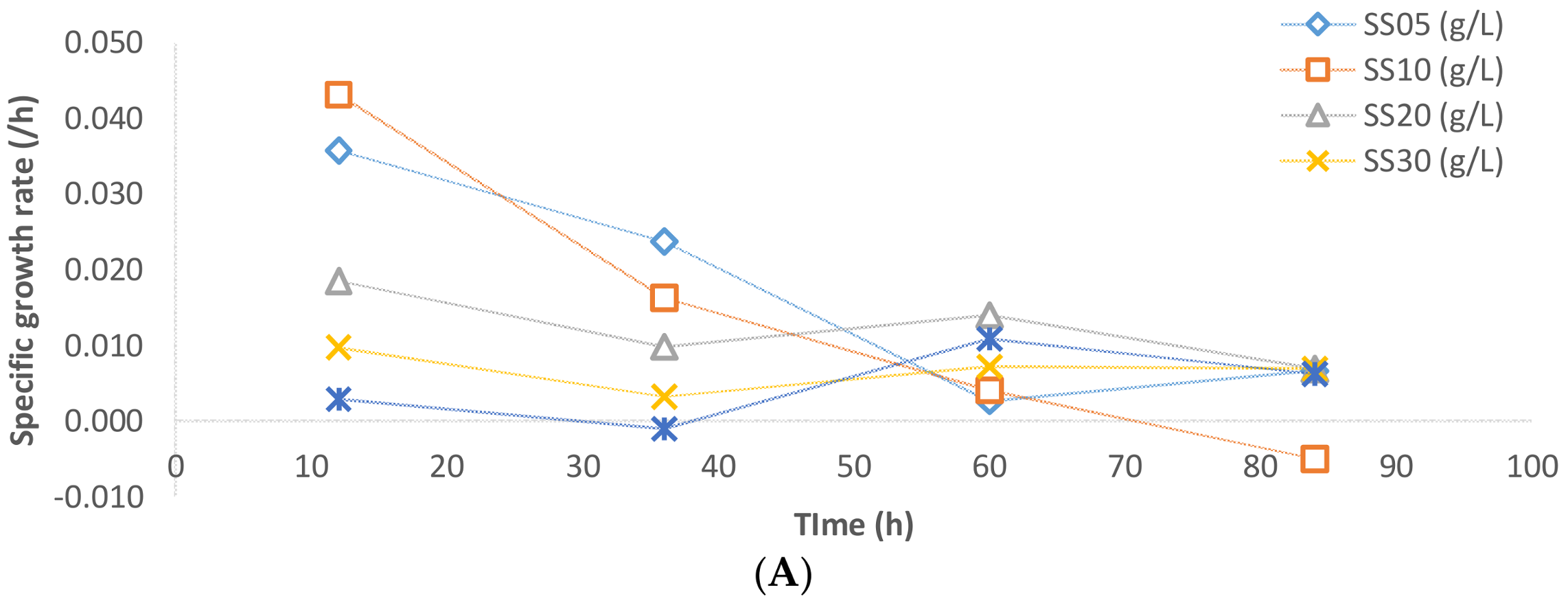
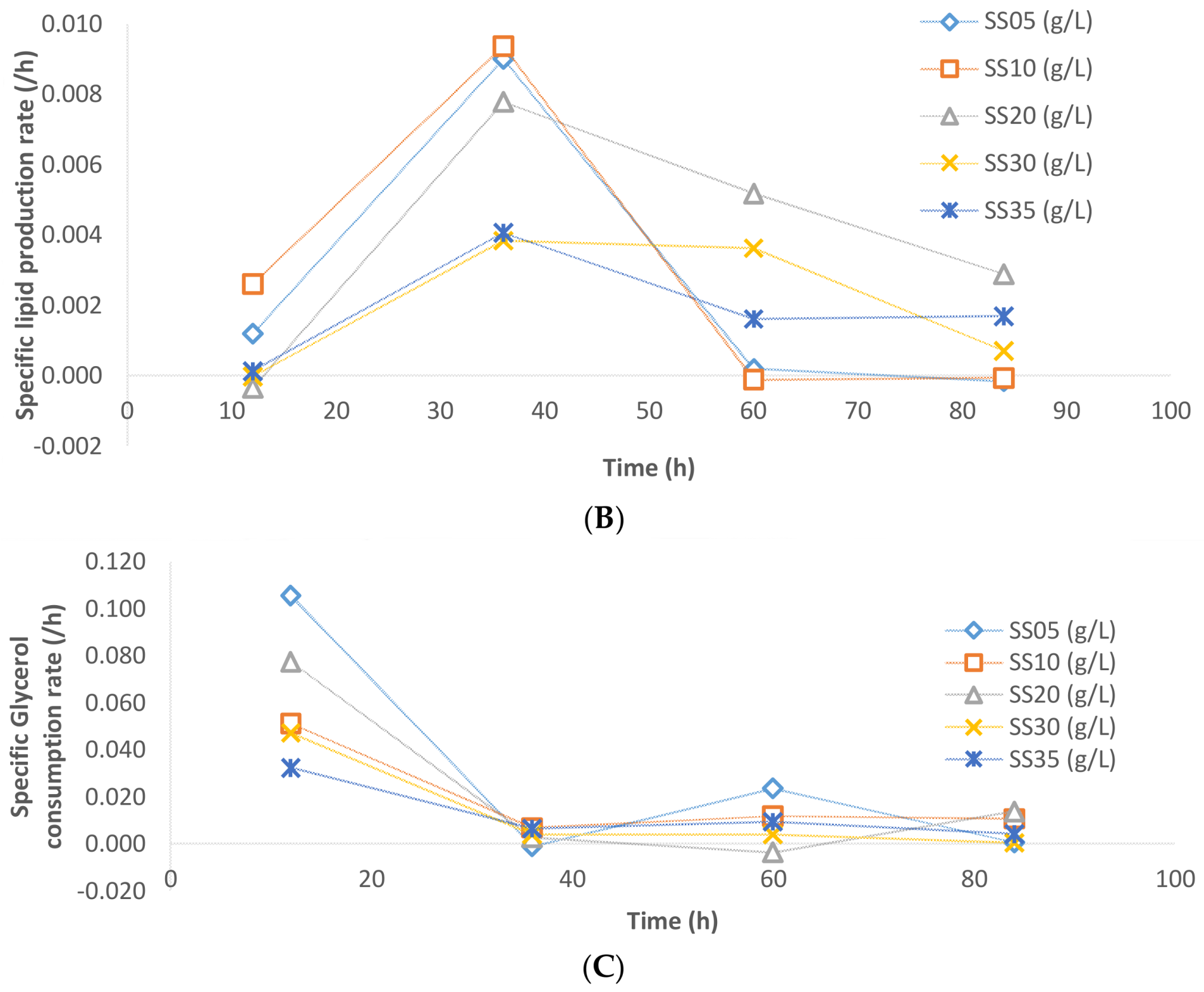
The specific rate of biomass formation, lipid accumulation, and glycerol consumption have been presented in
Figure 7 for various sludge solid concentrations. The specific biomass growth rate for 5 and 10 g/L solid concentration was monotonously decreasing throughout the fermentation. On the other hand, the specific biomass growth rate increased for the cases of 20, 30, and 35 g/L, where the SS of 20 g/L sludge fermentation has the highest specific rates of biomass growth (
Figure 7A).
In the case of lipid accumulation, during the 48 h of fermentation in the cases of 5, 10, and 20 g/L sludge SS, the specific lipid accumulation reached a maximum and started decreasing thereafter (
Figure 7B). At the 60th hour, the sludge fermentation with 20 g/L of sludge solids had the highest specific lipid accumulation rate while in the cases of 5 and 10 g/L sludge solids, the lipid accumulation ceased. A major fraction of lipid accumulation occurred after 30 h of fermentation. This is probably due to the fact that during fermentation, when carbon and nitrogen are consumed dynamically (in disproportionate quantities), the C:N ratio of the media changes inducing a metabolic shift to lipogenesis phase [
16,
19]. The specific glycerol uptake rates were similar in all cases of sludge solid concentration. At the beginning of the fermentation, the specific uptake rates were high mainly because of the active biomass synthesis that takes place during this period (
Figure 7C). Thereafter, the rate of glycerol consumption is significantly lower in all of the cases of sludge solid concentration.
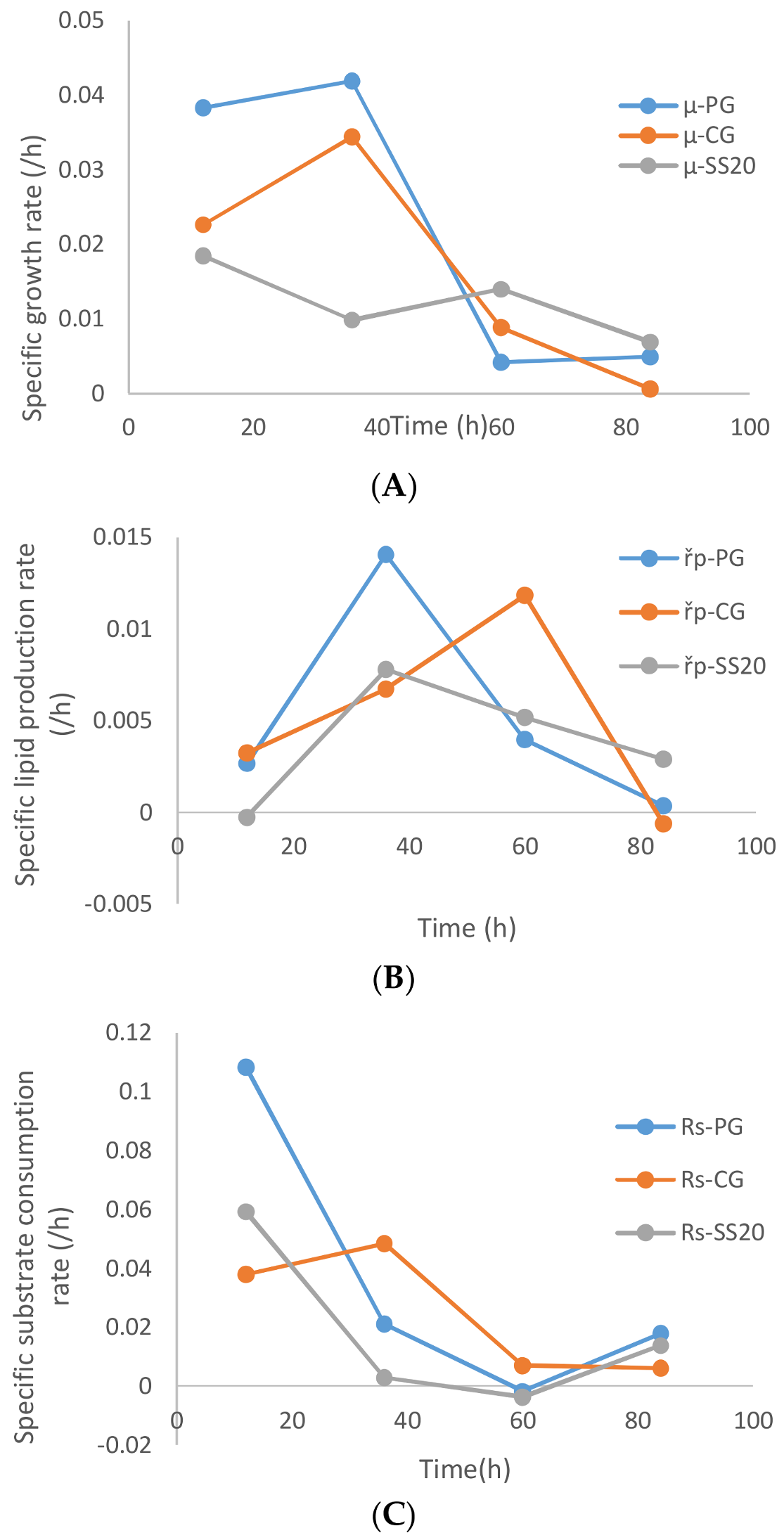
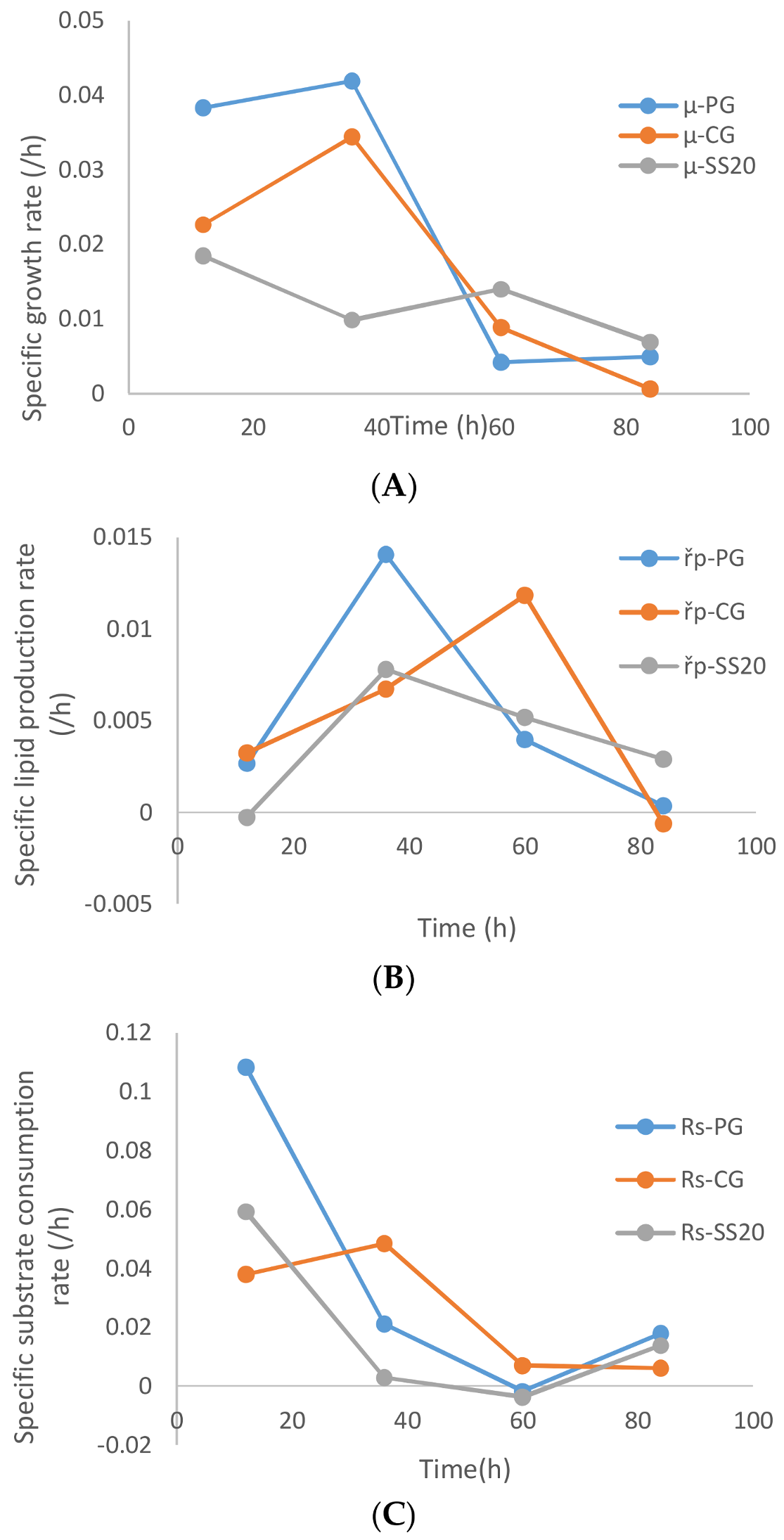
A comparison of the specific growth rate, product formation rate, and glycerol uptake rate between the cases of sludge solids, (20 g/L), crude glycerol, and pure glycerol is presented in
Figure 8A. As is apparent from
Figure 8A, the pure glycerol (without sludge) resulted in the highest specific biomass growth rate followed by crude glycerol and then sludge (optimized conc. 20 g/L). Exceptionally near zero specific biomass growth rate in the first 24 h of fermentation was observed with higher suspended solid concentrations (30 and 35 g/L,
Figure 7A). The stagnant biomass concentrations can be the results of the microbial action on the metabolizable suspended solids that are accompanied by sludge-like humic substances, loosely bound extra polysaccharides, nucleic acids, and other organics. The biomass growth of the microbe is compensated for by the decrease in sludge suspended solids concentration, which does not necessarily become completely assimilated (intracellularly) by the microbe. Similar observations were made by previous researchers [
5,
20].
The specific lipid accumulation rates were also highest for pure glycerol until the 36th hour of fermentation. Thereafter, the specific lipid accumulation rates decreased monotonously for the case of sludge and pure glycerol, but in the case of crude glycerol the specific lipid accumulation rates lagged at the 36th hour and major accumulation occurred after 50 h of fermentation (
Figure 8B). The specific glycerol uptake rates followed the trend of the specific lipid accumulation rate, where crude glycerol showed lower (but steady) consumption rates until the 36th hour of fermentation before it started decreasing from the 36th hour onward. In the case of sludge and pure glycerol, the glycerol uptake rates decreased continuously until 40 h of fermentation and then remained insignificantly low until the end of the fermentation.
3.6. Degree of Sludge Disposal Problem
During the process when the sludge is integrated into the media with the expectation that it will provide an alternate solution for sludge disposal toward the end, it is essential to analyze to what extent the problem is solved. A simple mass balance over the amount of solids available for disposal is used to evaluate the degree of sludge disposal (DSD) problem.
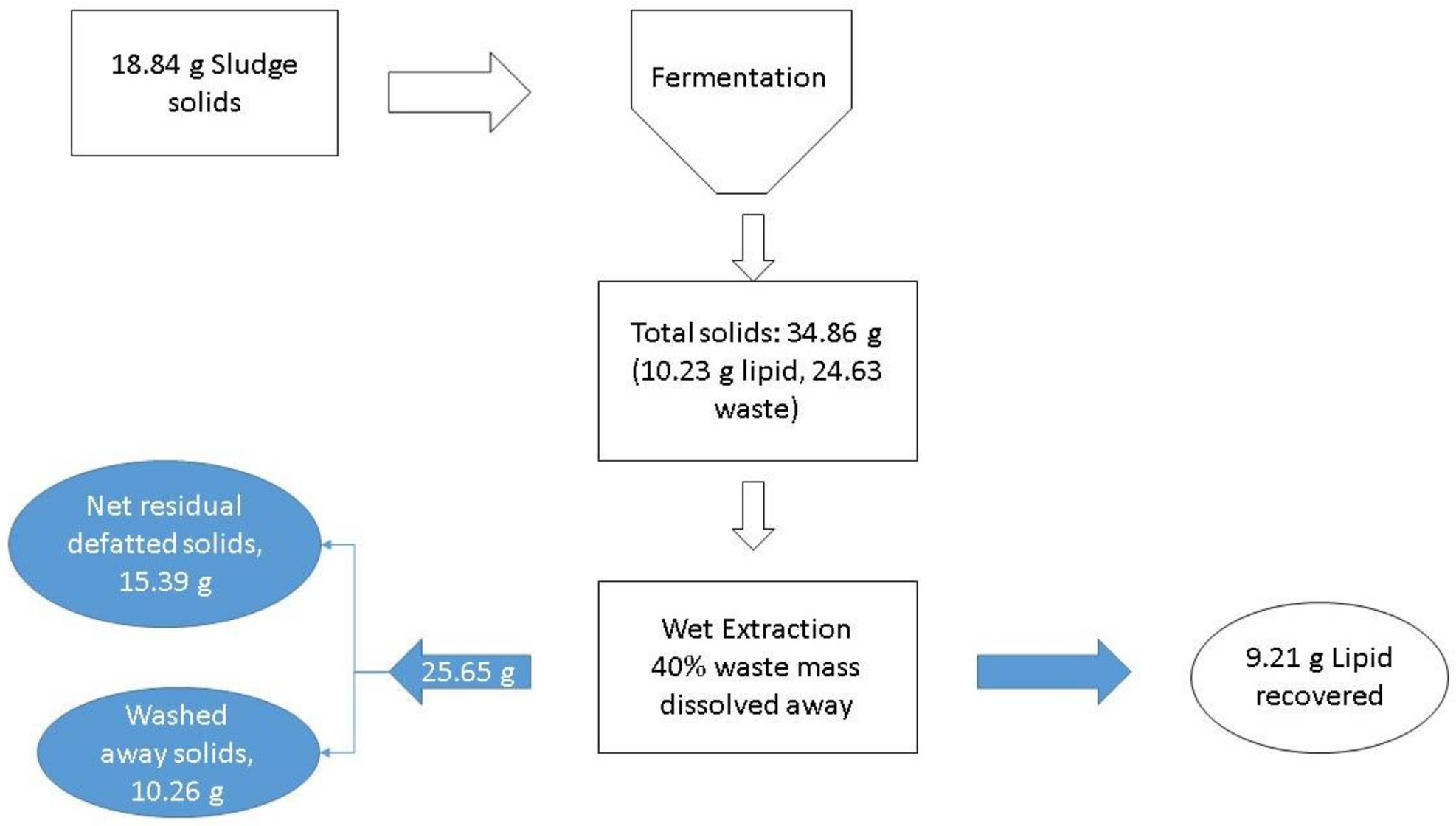
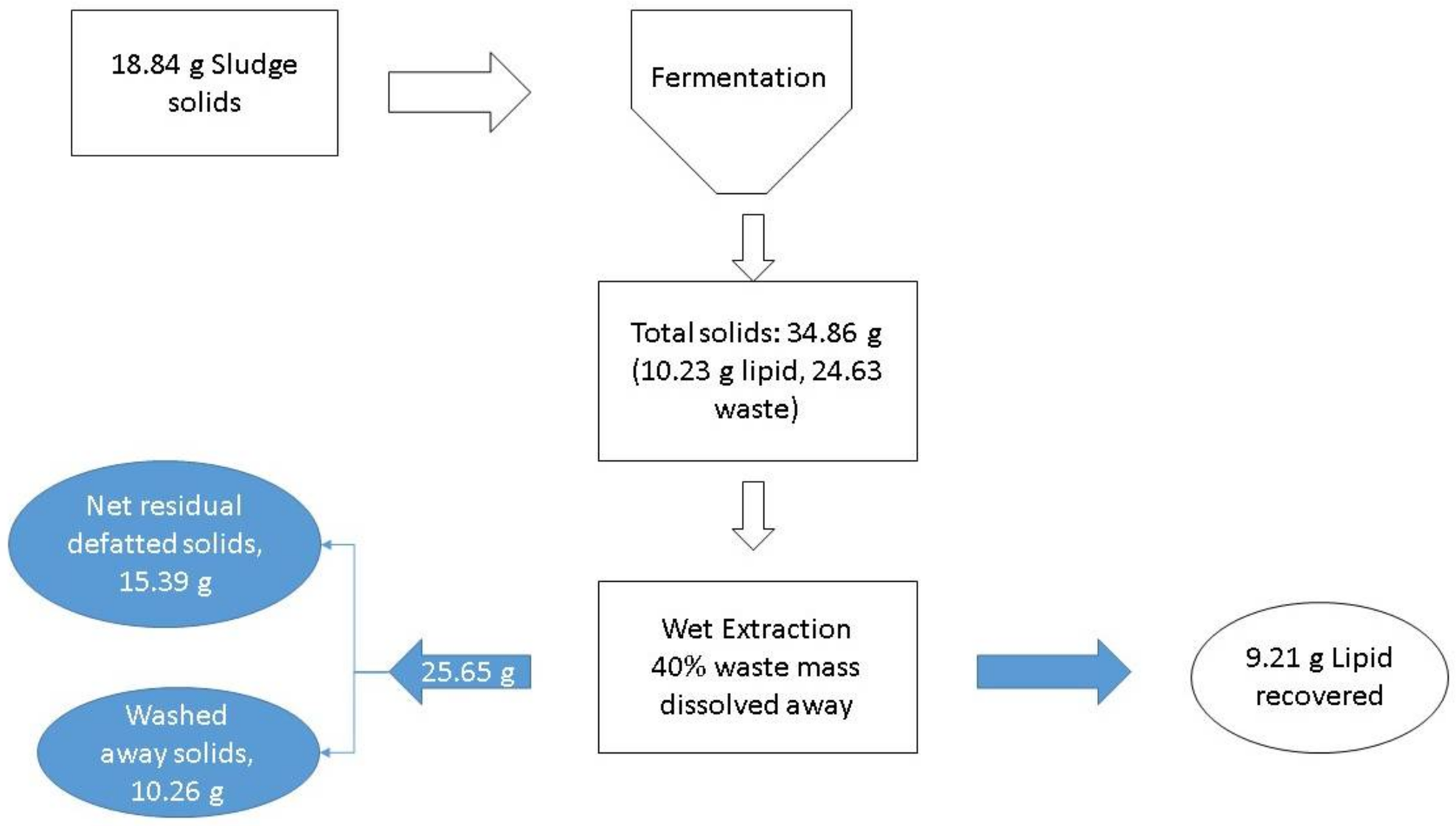
At the zeroth hour, the sludge to be disposed of is the sludge used in the process. For example, in the case of fermentation conducted with 20 g/L of sludge, at the zeroth hour 18.84 g of sludge was fed into the process (
Figure 9). Thus, the sludge to dispose of at the zeroth hour (SDZ) is 18.84 g. After the process is over, 34.86 g of solids are left which contain 10.23 g of lipids (lipid content 29.35%). Assuming 90% of these solids are recovered as microbial lipids, then the solids that remain to dispose of are 25.65 g-lipids. There are dry and wet methods of lipid extraction available in the literature. In the case of dry lipid, no further reduction is possible for these disposable solids, but in the case of wet extraction methods using detergents [
21], it is possible to dissolve a significant portion of de-fatted biomass by the chemical action of detergents. This soluble fraction in major part includes the proteins, carbohydrate, and nucleic acid (structural) components of the cell. Assuming 40% of the mass is lost during the extraction, the solids disposable after the process (SDP) will be 15.39 g [
22,
23]. Therefore, the DSD value for this case will be 81.69%. This indicates that there was a net reduction in the amount of solid waste to be disposed of. The lipid content is a very crucial parameter in determining the DSD of the process.
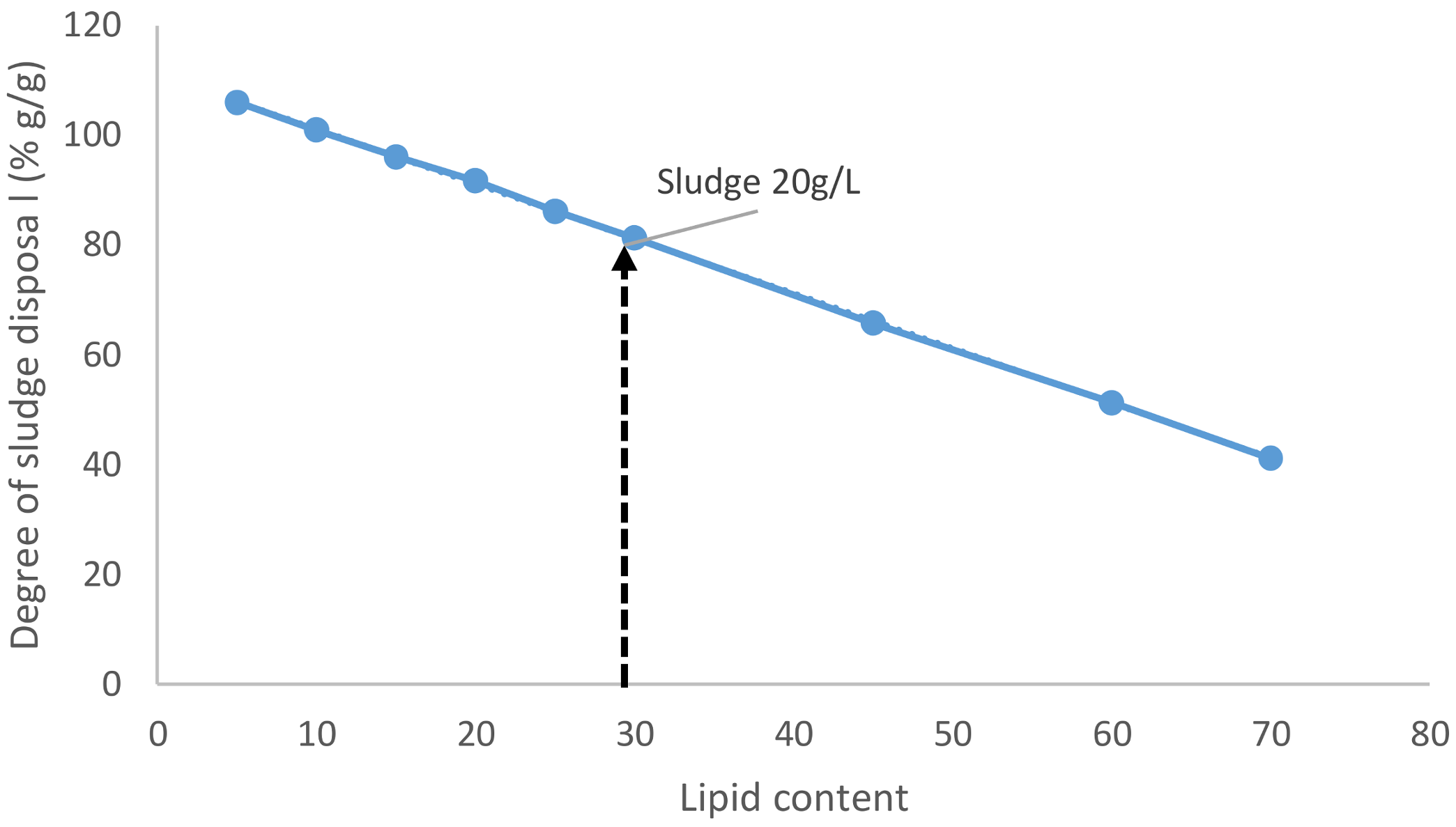
When the lipid content was varied from 5 to 70%, it was observed that at least 11.2% of lipid content is essential so that the situation remains neutral (with no net increase of solid waste) if not better off (
Figure 10). When the lipid content is increased, the amount of sludge available to dispose of after the process reduces further.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}