Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme

Department of Food Science and Nutrition, California Polytechnic State University, San Luis Obispo, CA 93407, USA
*
Author to whom correspondence should be addressed.
Fermentation 2017, 3(4), 53; https://doi.org/10.3390/fermentation3040053
Submission received: 2 September 2017
/
Revised: 30 September 2017
/
Accepted: 4 October 2017
/
Published: 10 October 2017
(This article belongs to the Special Issue Microbial Control)
Abstract
:The fungi Botrytis cinerea, Fusarium pallidoroseum, and Fusarium moniliforme are the causative agents of several plant diseases and can cause significant crop loss both before and after harvest. Fungicides are employed to control these phytopathogens, but fungicide use has led to an increase in resistance and may negatively affect the environment and human health. Hence, more environmentally sustainable solutions such as biological control methods are needed. The purpose of this study was to screen 22 bacterial isolates for inhibitory activity against fungal phytopathogens. To evaluate antifungal activity, the bacterial isolates were individually spot-inoculated onto Tryptic Soy Agar or de Man, Rogosa, Sharpe agar, and then a plug of fungal-colonized agar was placed onto the center of the isolate-inoculated plate. Plates were incubated at 24 °C for 10 days and fungal growth was evaluated. Nine of the 22 isolates screened inhibited all three fungi; inhibition by these isolates ranged from 51–62%, 60–68%, and 40–61% for B. cinerea, F. pallidoroseum, and F. moniliforme, respectively. Isolates were also screened for biosurfactant activity using the drop-collapse test. Bacillus megaterium, Bacillus coagulans, Bacillus thuringiensis and three Bacillus amyloliquefaciens isolates demonstrated strong biosurfactant activity and suppression of all three fungi, and therefore are recommended for further study.
1. Introduction
Crop loss due to phytopathogenic microorganisms has a dramatic impact on the agricultural industry. Botrytis cinerea and Fusarium species are fungal phytopathogens that cause substantial losses across a wide variety of crops both before and after harvest. In the field, B. cinerea, the fungus responsible for the plant diseases gray mold and Botrytis rot, affects over 200 crop species ranging from ornamentals to fruits and vegetables like lettuce, grapes, and strawberries [1,2]. Postharvest, B. cinerea causes the rapid decay of fresh produce because of its ability to invade damaged, weak, or rapidly senescing tissue [3], leading to a decreased shelf life. Fusarium Wilt and Fusarium Rot, caused by various Fusarium species, also affect a wide array of crops pre-harvest including lettuce, soybeans, strawberries, tomatoes, peppers, potatoes, and oranges [4,5,6]. Fusarium species are also problematic during postharvest storage, particularly in cereal grains and animal feeds [7]. The mycotoxins fumonisin B1 and B2, trichothecenes, and zearalenone are produced by some species of Fusarium and can cause disease in humans and animals [8,9]. Control measures to eliminate or mitigate the impact of B. cinerea and Fusarium species on quality and safety before or after harvest are needed.
Management of fungal phytopathogen pre- and post-harvest often includes the use of chemical fungicides. However, increases in fungicide resistance as well as health and environmental concerns associated with the use of harsh chemicals have created the need for an alternative means of treatment. The use of bioprotective bacteria pre-harvest or as a postharvest treatment could help to protect crops against fungi. Several bacterial species across a range of genera produce compounds active against phytopathogenic fungi. Synthesis of the antifungal lipopeptides has been identified in species of Bacillus, including Bacillus amyloliquefaciens [10,11,12], Bacillus subtilis [13,14], and Bacillus thuringiensis [15,16]. Some species of lactic acid bacteria (LAB) also produce antifungal compounds; these include organic acids, phenyllactic acid, 3-hydroxy fatty acids, and cyclic dipeptides [17,18,19]. Additional compounds produced by bacteria that have demonstrated antifungal activity include biosurfactant lipopeptides [20,21,22,23,24,25]. These bioactive lipopeptides, including surfactin, mycosubtilin, fengycin, and iturin, act on cell membranes, causing membrane disruption and cell leakage [26]. The antifungal activity of bioprotective bacteria in vitro has translated into fungal suppression when applied both in the field and postharvest.
Bioprotective bacteria have shown antifungal efficacy when applied to crops before harvest. Species of Bacillus have reduced the incidence of Fusarium disease in maize and tomato plants when applied pre-harvest [27,28] and decreased fruit rot caused by B. cinerea on strawberry plants [29]. Biocontrol agents have also suppressed fungal growth on fruits and vegetables when applied postharvest. Various species of Bacillus have been shown to reduce B. cinerea infection on pears, strawberries, and tomatoes [30,31,32]. Lactobacillus plantarum suppressed B. cinerea and Fusarium graminearum on cucumbers [33], and Pediococcus pentosaceus suppressed the growth of Penicillium expansum on pears [34]. Fungal suppression by various bacterial species demonstrates their potential use as biocontrol agents against fungal phytopathogens both before and after harvest.
Several species of Bacillus are commercially available as pre-harvest biocontrol treatments for various fruits and vegetables; Serenade® Optimum utilizes B. subtilis [35], Sonata® contains B. pumilus [36], and DoubleNickel LC uses B. amyloliquefaciens as the active ingredients [37].
The aim of this study was to screen and identify bacterial isolates that could potentially be used for the biological control of Botrytis cinerea, Fusarium pallidoroseum, and Fusarium moniliforme. Of the 22 isolates screened, six showed characteristics that indicated the potential for use as biocontrol agents and are therefore recommended for application-based research.
2. Materials and Methods
2.1. Fungal Pathogens
All fungi were obtained from the American Type Culture Collection (ATCC; Manassas, VA, USA). Working cultures of Botrytis cinerea ATCC 46522, Fusarium pallidoroseum ATCC 48152, and Fusarium moniliforme ATCC 60846 (teleomorph, Gibberella fujikuroi) (Table 1) were maintained on Potato Dextrose Agar (PDA, Becton, Dickson and Company; Sparks, MD, USA) at 4 °C and as fungal-colonized PDA plugs suspended in Potato Dextrose Broth (PDB, Becton, Dickson and Company; Sparks, MD, USA) with 25% glycerol at −70 °C for long-term storage.
2.2. Bioprotective Isolates
All bioprotective isolates were provided by BiOWiSH Technologies (Cincinnati, OH, USA). Working cultures of LAB isolates were maintained on de Man, Rogosa, Sharpe agar (MRS; Oxnoid; Basingstoke, Hampshire, UK) at 4 °C and all other bioprotective isolates on Tryptic Soy Agar (TSA; Remel, Lenexa, KS, USA) or TSA with 5% NaCl (w/w) at room temperature. For long-term storage, isolates were kept in the appropriate growth media (Table 1) with 15% glycerol at −70 °C with the exception of Bacillus clausii and Bacillus firmus—these two isolates were stored as spore crops at 6 °C. Prior to use in the screening, LAB isolates were inoculated into 10 mL of MRS Broth and incubated at 35 °C for 18–24 h. All other isolates were inoculated into 10 mL of Tryptic Soy Broth (TSB; Remel, Lenexa, KS, USA) or TSB with 5% NaCl (w/w) and incubated with agitation on an orbital shaker at either 30 or 35 °C, depending on the isolate, for 18–24 h.
2.3. Antifungal Screening
A loopful of prepared bioprotective culture was spot-inoculated in two locations directly across from each other near the perimeter of solidified media; the type of screening media was determined by the growth requirements of the bacterial isolate (Table 1). Using a sterile cork borer, a 9 mm plug was cut from the perimeter of a working culture of the fungal pathogen and placed in the center of the isolate-inoculated plate. Plates were wrapped in Parafilm® and incubated at 24 °C for 10 d. The diameter of the fungal colony between the two isolate inoculation locations was measured daily, beginning on Day 3. A fungus-inoculated plate of each screening media was used as a control. Each test was done in duplicate.
2.4. Biosurfactant Screening
Biosurfactant production by bioprotective isolates was evaluated via the drop-collapse test, which was adapted from Turgrul and Cansunar (2005) [38]. Briefly, each bioprotective isolate was cultured as previously described and incubated for 7 d; all isolates except LAB were incubated with agitation on an orbital shaker. After incubation, cultures were centrifuged at 3000 RPM for 15 min to separate the bacterial cells from the culture media; the resultant supernatant was used in the biosurfactant screening.
The surface of a petri dish was coated with mineral oil and allowed to sit at room temperature for at least 1 h. A 20 µL aliquot of bioprotective culture supernatant was placed onto the oiled surface, with the micropipette at a 45° angle relative to the surface of the petri dish. The diameter of the drop was then measured after 2 min. A 20 µL aliquot of fresh media (i.e., TSB or MRS) was treated in the same manner described above to serve as a negative media control.
The presence of a biosurfactant would reduce the surface tension of the bioprotective culture supernatant, thus causing the drop to spread on the oiled surface of the petri dish rather than hold its shape. Therefore, an isolate was considered to be a biosurfactant producer if the diameter of its supernatant drop was significantly larger than the diameter of the appropriate negative media control (α = 0.05).
2.5. Data Analysis
The antifungal screening was completed once in duplicate. Fungal diameters were averaged and percent fungal inhibition was determined using the following equation:
Calculation for the percent fungal inhibition:
The biosurfactant screening was completed twice in duplicate. Data were analyzed using Analysis of Variance (ANOVA); analysis was completed with Minitab 16 (Minitab Inc., State College, PA, USA). Dunnett’s test was used to identify isolate drop diameters that were significantly larger than the diameter of the negative media control (α = 0.05).
3. Results and Discussion
3.1. Antifungal Activity
3.1.1. Bacillus Species
Bacillus species have been investigated as possible biocontrol agents because of their ability to synthesize antifungal compounds, including lipopeptides and other antibiotics. Several Bacillus species have demonstrated antifungal activity against fungal phytopathogens, including B. cinerea and/or Fusarium species, in vitro [39,40,41,42,43]. However, antifungal capability is a function of bacterial strain and fungal pathogen. In this study, eight of the Bacillus isolates suppressed the growth of B. cinerea, F. pallidoroseum, and F. moniliforme (Table 2 and Figure 1); Bacillus megaterium exhibited the largest zones of inhibition against all three fungi, followed by Bacillus coagulans and several strains of B. amyloliquefaciens. Fungal inhibition by these isolates ranged from 51–62% for B. cinerea, 60–69% for F. pallidoroseum, and 40–61% for F. moniliforme (Table 2). Three Bacillus isolates inhibited B. cinerea but did not suppress the growth of the Fusarium species tested. While this screening identified 11 Bacillus isolates capable of fungal suppression in vitro; additional testing will be required to determine the efficacy against the phytopathogens when applied to produce.
Inhibition of fungal phytopathogens by Bacillus species has not only been observed in vitro but also in planta. The application of Bacillus species pre-harvest reduced the incidence and severity of diseases caused by F. oxysporum [42], Alternaria solani, and Phytophthora infestans [44] on tomato plants and B. cinerea fruit rot on strawberry plants [29]. Postharvest application of Bacillus species has also caused fungal suppression; decay was reduced on pears, tomatoes, bananas, and oranges [12,31,32,45]. Further testing of the isolates used in this study, including challenge studies on various types of produce, is needed to determine their possible use as biocontrol agents.
3.1.2. Lactic Acid Bacteria
Antifungal activity has also been detected from some LAB species; bioactive compounds produced by LAB include organic acids, phenyllactic acid, 3-hydroxy fatty acids, bacteriocins and bacteriocin-like substances, and cyclic dipeptides [17,18,19,46,47,48]. Three of the four LAB isolates used in this study inhibited B. cinerea, but not F. pallidoroseum or F. moniliforme. P. acidilactici demonstrated the strongest antifungal activity, followed by P. pentosaceus and L. plantarum; inhibition was 56%, 46%, and 36% respectively (Table 2). Although these isolates were only effective against B. cinerea, other LAB species have demonstrated broad-spectrum fungal inhibition when used as biocontrol agents on produce, but again efficacy varies between LAB and fungus species.
Pre- and pos-tharvest application of LAB to agricultural crops have shown mixed results. P. acidilactici and P. pentosaceus decreased Fusarium infection on wheat seeds when germinated on PDA, but failed to significantly reduce disease incidence during greenhouse trials [49]. In contrast, Lactobacillus species administered to tomato plants as a seed treatment or soil drench have promoted plant growth and mitigated the impact of F. oxysporum in potted trials [50]. When applied as a postharvest treatment, LAB reduced the decay caused by B. cinerea and F. graminearum on cucumbers [33] and decay by P. expansum on apples [51] and pears [34]. Again, additional investigation of the three antifungal LAB isolates identified in this study is needed to evaluate their use as biocontrol agents for the control of phytopathogenic fungi on produce.
3.1.3. Serratia Plymuthica
Serratia plymuthica is another species of bacteria that has demonstrated suppression of fungal phytopathogens. Synthesis of antifungal compounds such as pyrrolnitrin, haterumalide, siderophores, chitinolytic enzymes, and proteases has been detected in strains of S. plymuthica [52,53]. In this study, S. plymuthica suppressed the growth of B. cinerea, F. pallidoroseum, and F. moniliforme (Table 2), making it a possible candidate for additional evaluation. The use of other strains of S. plymuthica as a biocontrol agent both pre- and post-harvest has been investigated. Foliar application of S. plymuthica reduced disease caused by B. cinerea and Sclerotinia sclerotiorum in cucumber plants [52] and postharvest treatment of potatoes decreased the severity of dry rot caused by Fusarium sambucinum [54].
3.1.4. Isolates with No Antifungal Activity
Antifungal activity is not only a function of bioprotective isolates and fungus species. It can also be affected by growth conditions. Culture conditions such as media composition, pH, and incubation period and temperature can affect the production of antifungal compounds [55,56]. In this study, four Bacillus isolates, Lactobacillus amylovorus, Oceanobacillus sojae, and Ochrobactrum anthropi did not suppress B. cinerea, F. pallidoroseum, or F. moniliforme. Although these seven isolates did not inhibit any of the fungi against which they were challenged, this does not mean these isolates are incapable of producing antifungal compounds. The culture media and testing conditions used in this study may not have been optimal for the synthesis of bioactive compounds and further investigation is required to determine if these seven isolates truly do not inhibit B. cinerea, F. pallidoroseum, or F. moniliforme.
3.2. Biosurfactant Activity
3.2.1. Bacillus Species, Oceanobacillus sojae, and Ochrobactrum anthropi
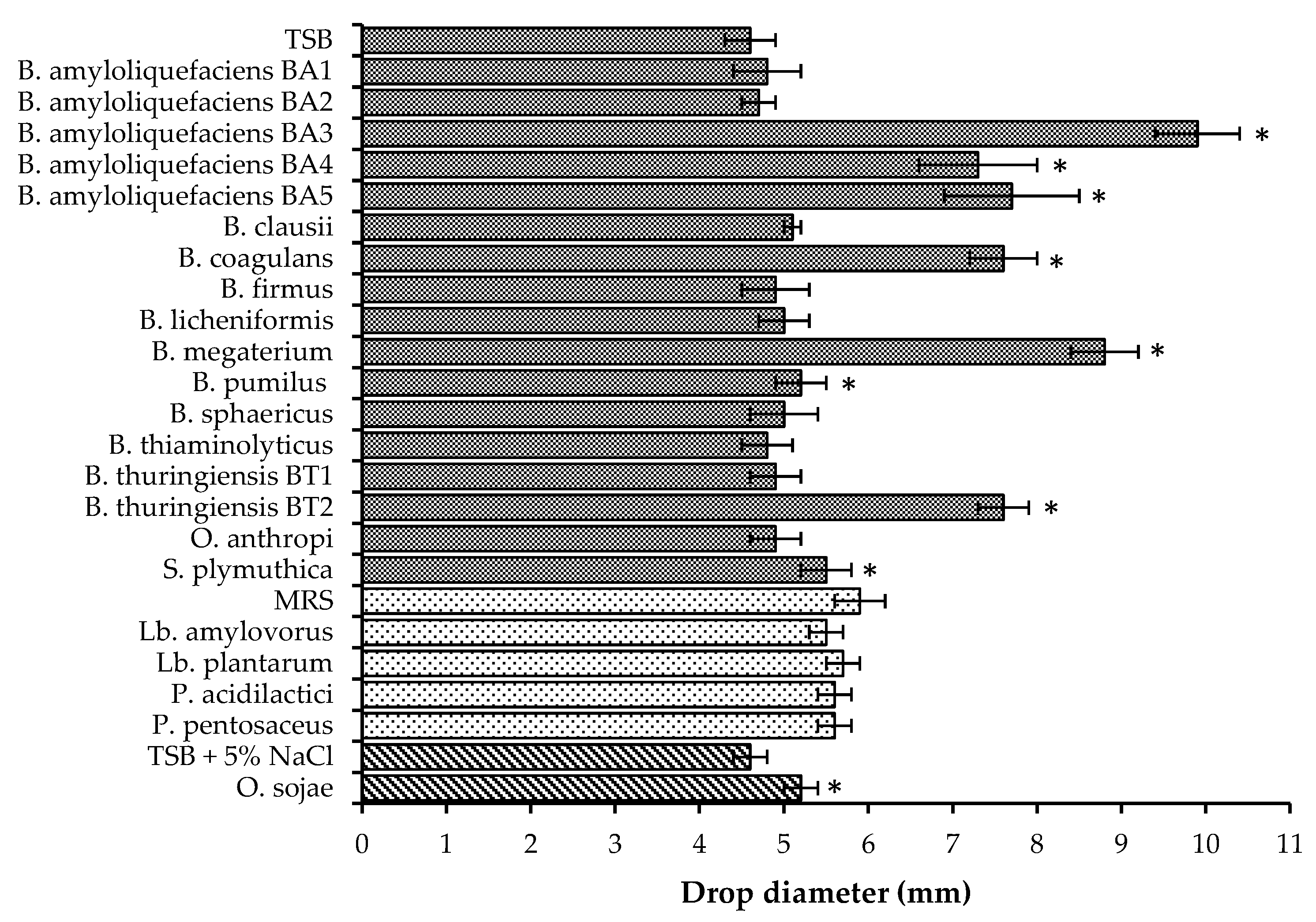
In this study, biosurfactant production was identified in seven Bacillus isolates (Figure 2). B. megaterium, B. coagulans, B. thuringiensis BT2 and several B. amyloliquefaciens isolates demonstrated both strong biosurfactant activity and suppression of B. cinerea, F. pallidoroseum, and F. moniliforme, implying the ability to synthesize antifungal lipopeptides. The production of antifungal lipopeptides has been identified in other strains of the species of Bacillus used in this study, including B. amyloliquefaciens [11,12,24], B. coagulans [21], B. megaterium [23], and B. thuringiensis [15,16,57]. B. pumilus and O. sojae demonstrated weak biosurfactant activity but no fungal suppression. This suggests that the biosurfactants secreted either did not have antifungal capabilities against the three fungi or that they were not produced in large enough quantities to suppress fungal growth.
Bacillus species also produce other compounds with antifungal activity, including antibiotics [22,41,58,59], proteinaceous substances [43,60], siderophores [22,61], and chitinolytic enzymes [27,40,62]. In this study, five Bacillus isolates demonstrated antifungal activity, but biosurfactant secretion was not detected; this indicates either the production of antifungal lipopeptides at low levels or that antifungal activity was not due to synthesized lipopeptides, but to some other bioactive compound. Biosurfactant production and antifungal activity were not detected in four Bacillus isolates and O. anthropi. The lack of both fungal inhibition and biosurfactant activity suggests that these strains did not secrete antifungal lipopeptides.
3.2.2. Lactic Acid Bacteria Isolates
Biosurfactant production was not detected in the four LAB isolates used in this study (Figure 2), but this does not mean that these isolates are incapable of producing biosurfactants. Biosurfactants produced by bacteria can either be excreted or remain cell-bound. Gudiña et al. [63] found that Lactobacillus coryniformis, Lactobacillus paracasei, L. plantarum and Leuconostoc mesenteroides produced cell-bound biosurfactants at higher levels than excreted ones. Cell-bound biosurfactant production has been observed in several species of Lactobacillus [64], including L. plantarum [65]. The drop-collapse test used in this study to determine biosurfactant production was only able to detect excreted biosurfactants and not those which are cell-bound; rescreening of LAB isolates with the addition of an extraction step may yield different results.
3.2.3. Serratia Plymuthica
The S. plymuthica isolate used in this study demonstrated weak biosurfactant activity (Figure 2) and suppressed the growth of B. cinerea, F. pallidoroseum, and F. moniliforme (Table 2). The production of antifungal compounds including pyrrolnitrin, haterumalide, siderophores, chitinolytic enzymes, and proteases has been identified in strains of S. plymuthica. This suggests that antifungal activity may have been due to the production of antifungal lipopeptides at low levels, the synthesis of some other type of bioactive compound, or a combination of lipopeptides and antibiotics.
4. Conclusions
The present study identified bacterial isolates with antifungal and/or biosurfactant activity. Of the 22 isolates tested, nine isolates—eight Bacillus isolates and S. plymuthica—suppressed the growth of B. cinerea, F. pallidoroseum, and F. moniliforme and six isolates—three Bacillus and three LAB—inhibited only B. cinerea. Biosurfactant production was detected in many of the Bacillus species which showed antifungal activity, suggesting the production of antifungal lipopeptides. Based on the results of this screening, six isolates—B. amyloliquefaciens strains BA3, BA4, and BA5, B. coagulans, B. megaterium, and B. thuringiensis BT2—are recommended for further evaluation. Additional evaluation should include screening with a larger panel of environmentally isolated fungal strains and characterization of the antifungal compounds produced by the bioprotective isolates. Finally, an investigation into the isolates’ antifungal efficacy on produce, both before and after harvest, is needed to evaluate their potential use as biocontrol agents in the agricultural industry.
Acknowledgments
This project was funded by the California State University Agricultural Research Institute (CSU-ARI) and BiOWiSH Technologies.
Author Contributions
Amanda Lathrop and Antoinette de Senna conceived and designed the experiments; Antoinette de Senna performed the experiments; Antoinette de Senna and Amanda Lathrop analyzed the data; Amanda Lathrop contributed reagents/materials/analysis tools; Antoinette de Senna and Amanda Lathrop wrote the paper.
Conflicts of Interest
The funding sponsors had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Swartzberg, D.; Kirshner, B.; Rav-David, D.; Elad, Y.; Granot, D. Botrytis cinerea induces senescence and is inhibited by autoregulated expression of the IPT gene. Eur. J. Plant Pathol. 2007, 120, 289–297. [Google Scholar] [CrossRef]
- UC IPM: UC Management Guidelines for Fusarium Wilt on Strawberry. Available online: http://www.ipm.ucdavis.edu/PMG/r734101411.html (accessed on 21 June 2014).
- Extension FactSheet: Fusarium and Verticillium Wilts of Tomato, Potato, Pepper, and Eggplant. Available online: https://www.oardc.ohio-state.edu/sallymiller/extension/factsheets/fusariumtomato.pdf (accessed on 21 June 2014).
- Díaz Arias, M.M.; Leandro, L.F.; Munkvold, G.P. Aggressiveness of Fusarium species and impact of root infection on growth and yield of soybeans. Phytopathology 2013, 103, 822–832. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Placinta, C.M.; MacDonald, A.M.C. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol. 1999, 80, 183–205. [Google Scholar] [CrossRef]
- Peraica, M.; Radic, B.; Lucic, A.; Pavlovic, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar] [PubMed]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Romero, D.; Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol Plant-Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Quan, C.; Wang, Y.; Wang, J.; Fan, S. Bacillus amyloliquefaciens Q-426 as a potential biocontrol agent against Fusarium oxysporum f. sp. spinaciae. J. Basic Microbiol. 2014, 54, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Jacobs, R.; Korsten, L. Iturin A is the principal inhibitor in the biocontrol activity of Bacillus amyloliquefaciens PPCB004 against postharvest fungal pathogens. J. Appl. Microbiol. 2010, 108, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Vanittanakom, N.; Leoffler, W. Fengycin—A novel antifungal lipopeptide antibiotic produced by Bacillus subtilis F-29-3. J. Antibiot. 1986, 39, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Moyne, A.; Shelby, R.; Cleveland, T.E.; Tuzun, S. Bacillomycin D: An iturin with antifungal activity against Aspergillus flavus. J. Appl. Microbiol. 2001, 90, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Mahata, D.; Paul, D.; Korpole, S.; Franco, O.L.; Mandal, S.M. Purification, biochemical characterization and self-assembled structure of a fengycin-like antifungal peptide from Bacillus thuringiensis strain SM1. Front. Microbiol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, P.I.; Bai, H.; Bai, D.; Chae, H.; Chung, S.; Kim, Y.; Park, R.; Chi, Y.T. Purification and characterization of a lipopeptide produced by Bacillus thuringiensis CMB26. J. Appl. Microbiol. 2004, 97, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.A.M.; Zannini, E.; Dal Bello, F.; Pawlowska, A.; Koehler, P.; Arendt, E.K. Lactobacillus amylovorus DSM 19280 as a novel food-grade antifungal agent for bakery products. Int. J. Food Microbiol. 2011, 146, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Strom, K.; Sjogren, J.; Broberg, A.; Schnurer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo (L-Phe-L-Pro) and cyclo(L-Phe-trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef] [PubMed]
- Magnusson, J.; Strom, K.; Roos, S.; Sjogren, J.; Schnurer, J. Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef]
- Arguelles-Arias, A.; Ongena, M.; Halimi, B.; Lara, Y.; Brans, A.; Joris, B.; Fickers, P. Bacillus amyloliquefaciens GA1 as a source of potent antibiotics and other secondary metabolites for biocontrol of plant pathogens. Microbiol. Cell Fact. 2009, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Vitullo, D.; Senatore, M.; Lima, G.; Lanzotti, V. Antifungal cyclic lipopeptides from Bacillus amyloliquefaciens strain BO5A. J. Nat. Prod. 2013, 76, 2019–2025. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Liu, X.; Li, C.; Tian, W.; Shen, Q.; Shen, B. Isolation of Bacillus amyloliquefaciens S20 and its application in control of eggplant bacterial wilt. J. Environ. Manag. 2014, 137, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Huszcza, E.; Burczyk, B. Surfactin isoforms from Bacillus coagulans. Z. Naturforsch. C 2006, 61, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Troyano Pueyo, M.; Bloch, C.; Carmona-Ribeiro, A.M.; di Mascio, P. Lipopeptides produced by a soil Bacillus megaterium strain. Microbiol. Ecol. 2009, 57, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Kalinovskaya, N.I.; Kuznetsova, T.A.; Ivanova, E.P.; Romanenko, L.A.; Voinov, V.G.; Huth, F.; Laatsch, H. Characterization of surfactin-like cyclic depsipeptides synthesized by Bacillus pumilus from Ascidian Halocynthia aurantium. Mar. Biotechnol. 2002, 4, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Makovitzki, A.; Shai, Y. pH-dependent antifungal lipopeptides and their plausible mode of action. Biochemistry 2005, 44, 9775–9784. [Google Scholar] [CrossRef] [PubMed]
- Bressan, W.; Figueiredo, J.E.F. Chitinolytic Bacillus spp. isolates antagonistic to Fusarium moniliforme in maize. J. Plant Pathol. 2010, 92, 343–347. [Google Scholar]
- Prabhukarthikeyan, R.; Saravanakumar, D.; Raguchander, T. Combination of endophytic Bacillus and Beauveria for the management of Fusarium wilt and fruit borer in tomato. Pest Manag. Sci. 2014, 70, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Ilhan, K.; Karabulut, O.A. Efficacy and population monitoring of bacterial antagonists for gray mold (Botrytis cinerea Pers. ex. Fr.) infecting strawberries. Biocontrol 2013, 58, 457–470. [Google Scholar] [CrossRef]
- Donmez, M.F.; Esitken, A.; Yildiz, H.; Ercisli, S. Biocontrol of Botrytis cinerea on strawberry fruit by plant growth promoting bacteria. J. Anim. Plant Sci. 2011, 21, 758–763. [Google Scholar]
- Mari, M.; Guizzardi, M.; Brunelli, M.; Folchi, A. Postharvest biological control of grey mould (Botrytis cinerea Pers.: Fr.) on fresh-market tomatoes with Bacillus amyloliquefaciens. Crop Prot. 1996, 15, 699–705. [Google Scholar] [CrossRef]
- Mari, M.; Guizzardi, M.; Pratella, G.C. Biological control of Gray Mold in pears by antagonistic bacteria. Biol. Control. 1996, 7, 30–37. [Google Scholar] [CrossRef]
- Sathe, S.J.; Nawani, N.N.; Dhakephalkar, P.K.; Kapadnis, B.P. Antifungal lactic acid bacteria with potential to prolong shelf-life of fresh vegetables. J. Appl. Microbiol. 2007, 103, 2622–2628. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Mahony, J.; van Sinderen, D. Broad-spectrum antifungal-producing lactic acid bacteria and their application in fruit models. Folia Microbiol. Praha 2013, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Serenade Optimum (130205). Available online: https://greenbook-assets.s3.amazonaws.com/L1302.05A.pdf (accessed on 19 June 2014).
- Bayer CropScience Safety Data Sheet, Sonata. Available online: http://ag.wilburellis.com/Products/Product%20Documents/Actives/SDS/SONATA-SDS%20US-Default.pdf (accessed on 19 June 2014).
- DoubleNickel LC Biofungicide. Available online: http://www.certisusa.com/pdf-labels/DoubleNickel_LC_label.pdf (accessed on 19 June 2014).
- Turgrul, T.; Cansunar, E. Detecting surfactant-producing microorganisms by the drop-collapse test. World J. Microbiol. Biotechnol. 2005, 21, 851–853. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, Y.; Zheng, L.; Yang, X.; Liu, H.; Guo, J. Isolation and characterization of an antifungal protein from Bacillus licheniformis HS10. Biochem. Biophys. Res. Commun. 2014, 454, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Gomaa, E.Z. Chitinase production by Bacillus thuringiensis and Bacillus licheniformis: Their potential in antifungal biocontrol. J. Microbiol. 2012, 50, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Tendulkar, S.R.; Saikumari, Y.K.; Patel, V.; Raghotama, S.; Munshi, T.K.; Balaram, P.; Chattoo, B.B. Isolation, purification and characterization of an antifungal molecule produced by Bacillus licheniformis BC98, and its effect on phytopathogen Magnaporthe grisea. J. Appl. Microbiol. 2007, 103, 2331–2339. [Google Scholar] [CrossRef] [PubMed]
- Saidi, N.; Kouki, S.; Hiri, F.M.; Hajlaoui, M.R.; Mahrouk, M.; Ouzari, H.; Jedidi, N.; Hassen, A. Characterization and selection of Bacillus sp. strains, effective biocontrol agents against Fusarium oxysporum f. sp. radicis-lycopersici, the causal agent of Fusarium crown and root rot in tomato. Ann. Microbiol. 2009, 59, 191–198. [Google Scholar] [CrossRef]
- Agarry, O.O.; Akinyosoye, F.A.; Adetuyi, F.C. Antagonistic properties of microogranisms associated with cassava (Manihot esculenta Crantz) products. Afr. J. Biotechnol. 2005, 4, 627–632. [Google Scholar] [CrossRef]
- Chowdappa, P.; Mohan Kumar, S.P.; Jyothi Lakshmi, M.; Upreti, K.K. Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum OTPB3. Biol. Control. 2013, 65, 109–117. [Google Scholar] [CrossRef]
- Alvindia, D.G.; Natsuaki, K.T. Biocontrol activities of Bacillus amyloliquefaciens DGA14 isolated from banana fruit surface against banana crown rot-causing pathogens. Crop Prot. 2009, 28, 236–242. [Google Scholar] [CrossRef]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H.S. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Voulgari, K.; Hatzikamari, M.; Delepoglou, A.; Georgakopoulos, P.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Antifungal activity of non-starter lactic acid bacteria isolates from dairy products. Food Control. 2010, 21, 136–142. [Google Scholar] [CrossRef]
- Gerez, C. L.; Torres, M.J.; Font de Valdez, G.; Rollán, G. Control of spoilage fungi by lactic acid bacteria. Biol. Control. 2013, 64, 231–237. [Google Scholar] [CrossRef]
- Suproniene, S.; Semaskiene, R.; Juodeikiene, G.; Mankeviciene, A.; Cizeikiene, D.; Vidmantiene, D. Seed treatment with lactic acid bacteria against seed-borne pathogens of spring wheat. Biocontrol. Sci. Technol. 2014, 25, 144–154. [Google Scholar] [CrossRef]
- Hamed, H.A.; Moustafa, Y.A.; Abdel-Aziz, S.M. In vivo efficacy of Lactic Acid Bacteria in biological control against Fusarium oxysporum for protection of tomato plant. Life Sci. J. 2011, 8, 462–468. [Google Scholar] [CrossRef]
- Trias, R.; Bañeras, L.; Montesinos, E.; Badosa, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. Microbiol. 2008, 11, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Kamensky, M.; Ovadis, M.; Chet, I.; Chernin, L. Soil-borne strain IC14 of Serratia plymuthica with multiple mechanisms of antifungal activity provides biocontrol of Botrytis cinerea and Sclerotinia sclerotiorum diseases. Soil Biol. Biochem. 2003, 35, 323–331. [Google Scholar] [CrossRef]
- Levenfors, J.J.; Hedman, R.; Thaning, C.; Gerhardson, B.; Welch, C.J. Broad-spectrum antifungal metabolites produced by the soil bacterium Serratia plymuthica A 153. Soil Biol. Biochem. 2004, 36, 677–685. [Google Scholar] [CrossRef]
- Gould, M.; Nelson, L.M.; Waterer, D.; Hynes, R.K. Biocontrol of Fusarium sambucinum, dry rot of potato, by Serratia plymuthica 506. Biocontrol. Sci. Technol. 2008, 18, 1005–1016. [Google Scholar] [CrossRef]
- Mandal, V.; Sen, S.K.; Mandal, N.C. Optimized culture conditions for bacteriocin production by Pediococcus acidilactici LAB 5 and its characterization. Indian J. Biochem. Biophys. 2008, 45, 106–110. [Google Scholar] [PubMed]
- Shih, I.L.; Kuo, C.Y.; Hsieh, F.C.; Kao, S.S.; Hsieh, C. Use of surface response methodology to optimize culture conditions for iturin A production by Bacillus subtilis in solid-state fermentation. J. Chin. Inst. Chem. Eng. 2008, 39, 635–643. [Google Scholar] [CrossRef]
- Hathout, Y.; Ho, Y.; Ryzhov, V.; Demirev, P.; Fenselau, C. Kurstakins: A new class of lipopeptides isolated from Bacillus thuringiensis. J. Nat. Prod. 2000, 63, 1492–1496. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, B.; Zhang, N.; Waseem, R.; Shen, Q.; Huang, Q. Production of bacillomycin- and macrolactin-type antibiotics by Bacillus amyloliquefaciens NJN-6 for suppressing soilborne plant pathogens. J. Agric. Food Chem. 2012, 60, 2976–2981. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.; Chen, X.H.; Vater, J.; Franke, P.; Nicholson, G.; Borriss, R.; Süssmuth, R.D. Macrolactin is the polyketide biosynthesis product of the pks2 cluster of Bacillus amyloliquefaciens FZB42. J. Nat. Prod. 2007, 70, 1417–1423. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Lin, B.; Zhang, R. A novel antifungal protein of Bacillus subtilis B25. Springerplus 2013, 2, 543. [Google Scholar] [CrossRef] [PubMed]
- Chaiharn, M.; Chunhaleuchanon, S.; Lumyong, S. Screening siderophore producing bacteria as potential biological control agent for fungal rice pathogens in Thailand. World J. Microbiol. Biotechnol. 2009, 25, 1919–1928. [Google Scholar] [CrossRef]
- Aktuganov, G.E.; Melent’ev, A.I.; Kuz’mina, L.Y.; Galimzyanova, N.F.; Shirokov, A.V. The chitinolytic activity of Bacillus cohn bacteria antagonistic to phytopathogenic fungi. Microbiology 2003, 72, 313–317. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactant-producing lactobacilli: Screening, production profiles, and effect of medium composition. Appl. Environ. Soil Sci. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Antimicrobial and anti-adhesive properties of biosurfactant produced by lactobacilli isolates, biofilm formation and aggregation ability. J. Gen. Appl. Microbiol. 2013, 59, 425–436. [Google Scholar] [CrossRef]
- Velraeds, M.M.; Van Der Mei, H.C.; Reid, G.; Busscher, H.J. Inhibition of initial adhesion of uropathogenic Enterococcus faecalis by biosurfactants from Lactobacillus isolates. Appl. Environ. Microbiol. 1996, 62, 1958–1963. [Google Scholar] [PubMed]
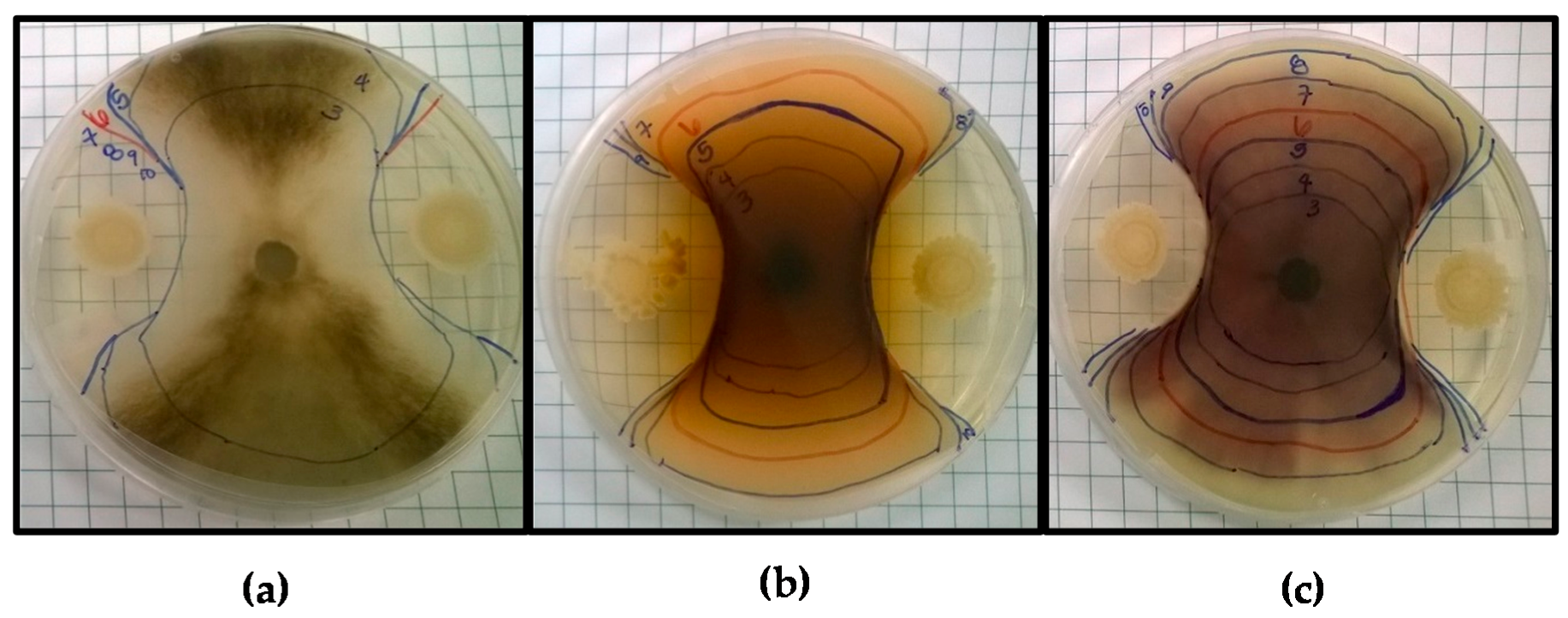
Figure 1.
Antifungal screening of Bacillus amyloliquefaciens BA4 against (a) Botrytis cinerea; (b) Fusarium pallidoroseum, and (c) Fusarium moniliforme. Markings indicate the border of the fungal colony each day of incubation, beginning on Day 3 up until Day 10 or the last day of observed growth.
Figure 1.
Antifungal screening of Bacillus amyloliquefaciens BA4 against (a) Botrytis cinerea; (b) Fusarium pallidoroseum, and (c) Fusarium moniliforme. Markings indicate the border of the fungal colony each day of incubation, beginning on Day 3 up until Day 10 or the last day of observed growth.
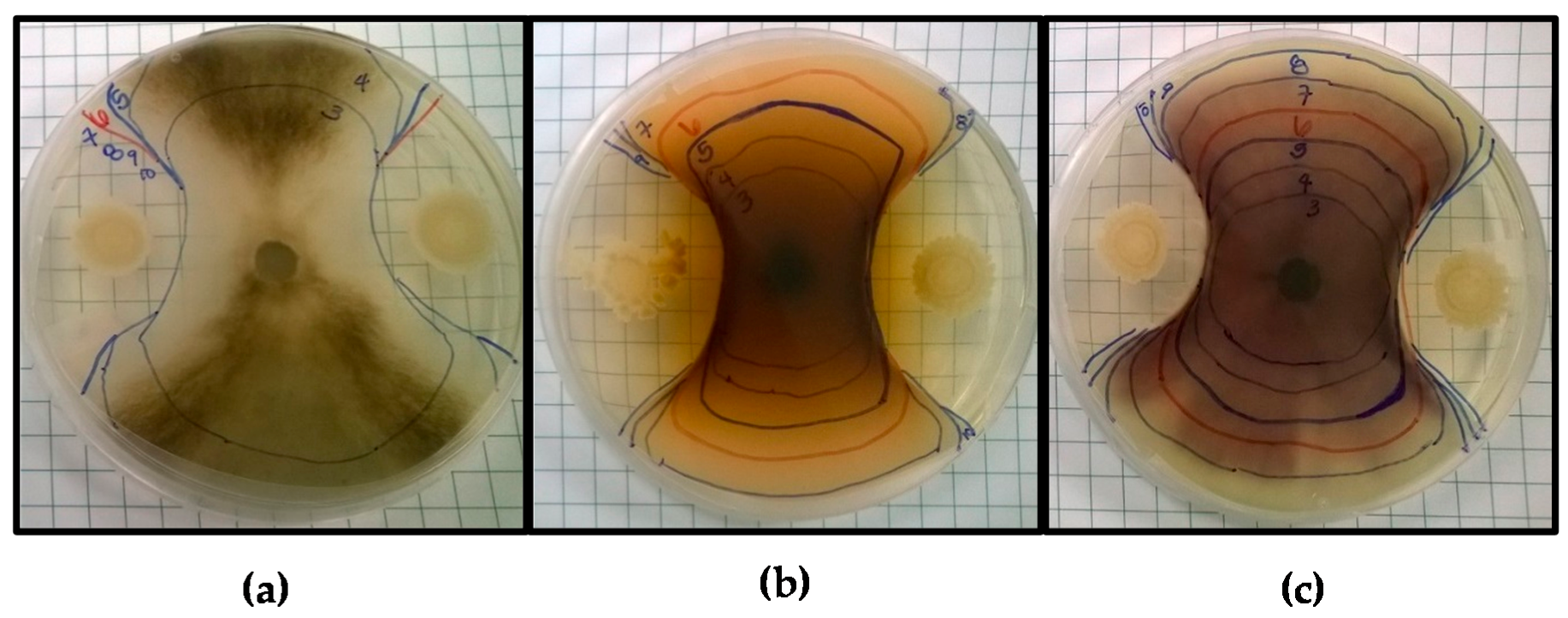
Figure 2.
Biosurfactant activity of bioprotective isolates. Error bars indicate standard deviation. Media used are designated by bars with corresponding patterns. Cultures grown in TSB ![Fermentation 03 00053 i001]() , MRS
, MRS ![Fermentation 03 00053 i002]() , and TSB + 5% NaCl
, and TSB + 5% NaCl ![Fermentation 03 00053 i003]() . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
. Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
 , MRS
, MRS  , and TSB + 5% NaCl
, and TSB + 5% NaCl  . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
. Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
Figure 2.
Biosurfactant activity of bioprotective isolates. Error bars indicate standard deviation. Media used are designated by bars with corresponding patterns. Cultures grown in TSB ![Fermentation 03 00053 i001]() , MRS
, MRS ![Fermentation 03 00053 i002]() , and TSB + 5% NaCl
, and TSB + 5% NaCl ![Fermentation 03 00053 i003]() . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
. Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).
, MRS , and TSB + 5% NaCl . Asterisks indicate drop diameters significantly larger than that of the media control (α = 0.05).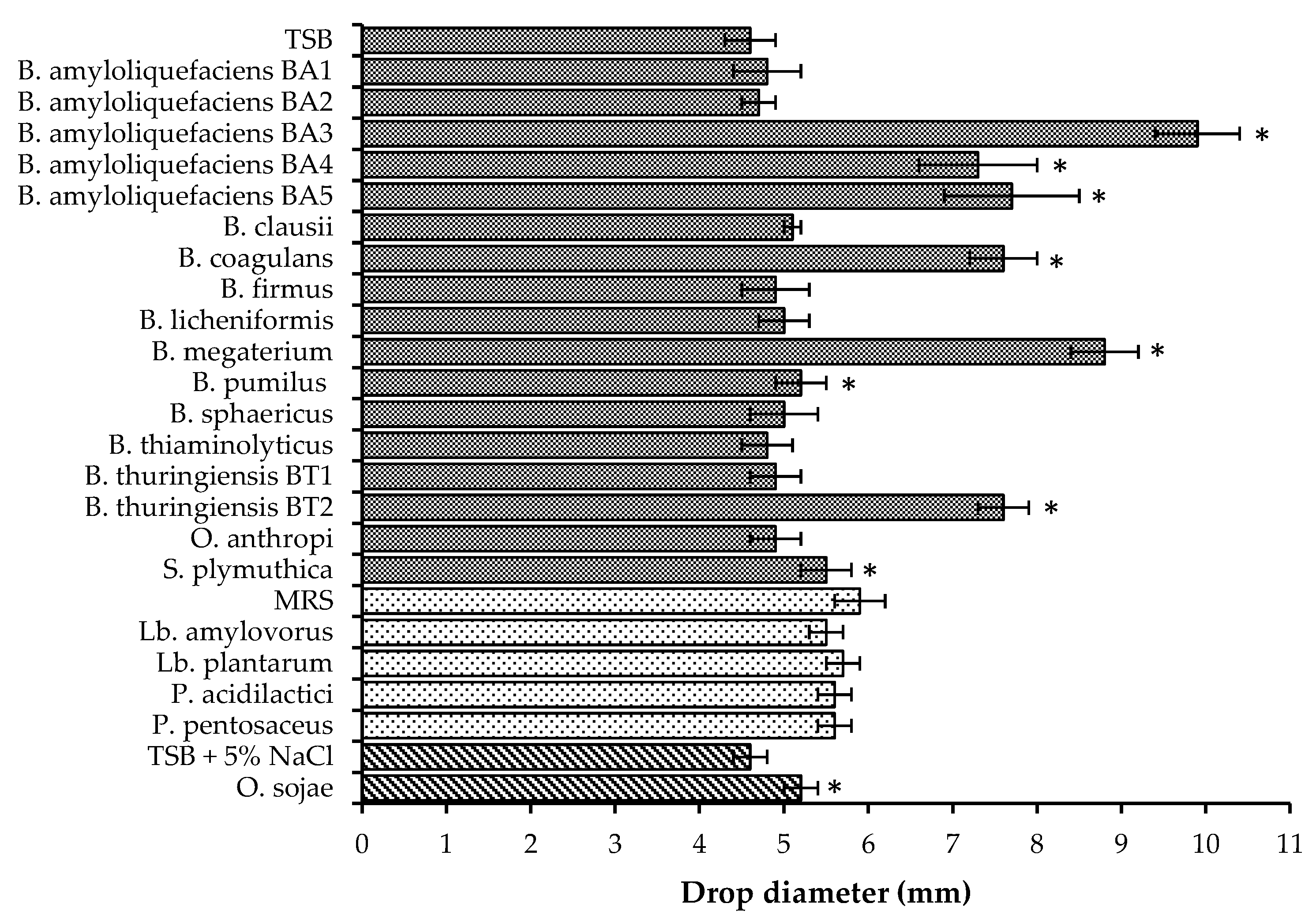

{kind=link}
{kind=link}
Table 1.
Bioprotective isolates and fungal phytopathogens.
| Isolate | Growth Medium | Incubation Temperature | Screening Medium |
|---|---|---|---|
| Bioprotective isolate | |||
| Bacillus amyloliquefaciens BA1 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA2 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA3 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA4 | TSB | 35 °C | PDA |
| Bacillus amyloliquefaciens BA5 | TSB | 35 °C | PDA |
| Bacillus clausii | TSB | 30 °C | PDA |
| Bacillus coagulans | TSB | 35 °C | PDA |
| Bacillus firmus | TSB | 30 °C | TSA |
| Bacillus licheniformis | TSB | 35 °C | PDA |
| Bacillus megaterium | TSB | 35 °C | PDA |
| Bacillus pumilus | TSB | 35 °C | PDA |
| Bacillus sphaericus | TSB | 30 °C | PDA |
| Bacillus thiaminolyticus | TSB | 30 °C | TSA |
| Bacillus thuringiensis BT1 | TSB | 35 °C | PDA |
| Bacillus thuringiensis BT2 | TSB | 35 °C | PDA |
| Lactobacillus amylovorus | MRS | 35 °C | MRS |
| Lactobacillus plantarum | MRS | 35 °C | MRS |
| Oceanobacillus sojae | TSB w/5% NaCl (w/w) | 35 °C | TSA |
| Ochrobactrum anthropi | TSB | 35 °C | PDA |
| Pediococcus acidilactici | MRS | 35 °C | MRS |
| Pediococcus pentosaceus | MRS | 35 °C | MRS |
| Serratia plymuthica | TSB | 30 °C | PDA |
| Fungal phytopathogen | |||
| Botrytis cinerea | PDA | 24 °C | N/A 1 |
| Fusarium pallidoroseum | PDA | 24 °C | N/A 1 |
| Fusarium moniliforme | PDA | 24 °C | N/A 1 |
TSB: Tryptic Source Broth, MRS: Man, Rogosa, Sharpe agar, PDA: Potato Dextrose Agar. TSA: Tryptic Source Agar. N/A 1: not applicable.
Table 2.
Fungal inhibition of bioprotective isolates against Botrytis cinerea, Fusarium moniliforme, and Fusarium pallidoroseum after 10 d incubation at 24 °C.
Table 2.
Fungal inhibition of bioprotective isolates against Botrytis cinerea, Fusarium moniliforme, and Fusarium pallidoroseum after 10 d incubation at 24 °C.
| Bioprotective Isolate | Inhibition | ||
|---|---|---|---|
| Botrytis cinerea | Fusarium pallidoroseum | Fusarium moniliforme | |
| B. amyloliquefaciens BA1 | 52% | 62% | 53% |
| B. amyloliquefaciens BA2 | 56% | 63% | 60% |
| B. amyloliquefaciens BA3 | 58% | 67% | 61% |
| B. amyloliquefaciens BA4 | 60% | 67% | 60% |
| B. amyloliquefaciens BA5 | 61% | 69% | 61% |
| B. clausii | 30% | N/I 1 | N/I 1 |
| B. coagulans | 62% | 66% | 59% |
| B. firmus | 49% | N/I 1 | N/I 1 |
| B. licheniformis | N/I 1 | N/I1 | N/I 1 |
| B. megaterium | 62% | 68% | 61% |
| B. pumilus | N/I 1 | N/I 1 | N/I 1 |
| B. sphaericus | N/I 1 | N/I 1 | N/I 1 |
| B. thiaminolyticus | 50% | N/I 1 | N/I 1 |
| B. thuringiensis BT1 | N/I 1 | N/I 1 | N/I 1 |
| B. thuringiensis BT2 | 56% | 68% | 58% |
| L. amylovorus | N/I 1 | N/I 1 | N/I 1 |
| L. plantarum | 36% | N/I 1 | N/I 1 |
| O. sojae | N/I 1 | N/I 1 | N/I 1 |
| O. anthropi | N/I 1 | N/I 1 | N/I 1 |
| P. acidilactici | 56% | N/I 1 | N/I 1 |
| P. pentosaceus | 46% | N/I 1 | N/I 1 |
| S. plymuthica | 51% | 60% | 40% |
N/I 1: no inhibition; fungal colony grew over/around bacterial isolate colony.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
De Senna, A.; Lathrop, A. Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme. Fermentation 2017, 3, 53. https://doi.org/10.3390/fermentation3040053
AMA Style
De Senna A, Lathrop A. Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme. Fermentation. 2017; 3(4):53. https://doi.org/10.3390/fermentation3040053
Chicago/Turabian StyleDe Senna, Antoinette, and Amanda Lathrop. 2017. "Antifungal Screening of Bioprotective Isolates against Botrytis cinerea, Fusarium pallidoroseum and Fusarium moniliforme" Fermentation 3, no. 4: 53. https://doi.org/10.3390/fermentation3040053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.